
Abstract
Rupture of vulnerable atheroma often underlies acute coronary syndromes. Vulnerable plaques exhibit a paucity of vascular smooth muscle cells (VSMCs) in the cap. Therefore, decreased VSMC migration into the neointima may predispose to vulnerability. The balance between cell surface plasminogen activator activity and its inhibition [mediated primarily by plasminogen activator inhibitor type 1 (PAI-1)] modulates migration of diverse types of cells. We sought to determine whether increased expression of PAI-1 would decrease migration of VSMCs in vitro and neointimal cellularity in vivo in apolipo-protein E knockout (ApoE−-/–) mice fed a high-fat diet. Increased vessel wall expression of PAI-1 in transgenic mice was induced with the SM22α promoter. VSMC migration through Matrigel in vitro was quantified with laser scanning cytometry. Expression of PAI-1 was increased threefold in the aortic wall of SM22-PAI transgene-positive mice. Neointimal cellularity of vascular lesions was decreased by 26% (p=0.01; n=5 each) in ApoE−-/– mice with the SM22-PAI transgene compared with ApoE−-/– mice. VSMCs explanted from transgene-positive mice exhibited twofold greater expression of PAI-1 and their migration was attenuated by 27% (p=0.03). Accordingly, increased expression of PAI-1 protein by VSMCs reduces their migration in vitro and their contribution to neointimal cellularity in vivo.
A
We have hypothesized that limitation of activation of intramural plasminogen activators by their primary inhibitor, plasminogen activator inhibitor type 1 (PAI-1), is one factor predisposing to evolution of vulnerable plaques by promoting or retarding neointimal migration of VSMCs, leading to a paucity of such cells and consequent elaboration of thin rather than thick fibrous plaques (Sobel 1999; Sobel et al. 2003). The present study was performed to determine whether decreased migration of VSMCs, characterized in vitro, would ensue if VSMC expression of PAI-1 were increased with the use of the promoter region of the VSMC gene SM-22α. In addition, we determined whether neointimal cellularity would be diminished in vivo by overexpression of VSMC PAI-1 during the evolution of atheroma in apolipoprotein E (ApoE)-deficient mice. A specific objective was to characterize the effect of overexpression of PAI-1 on the evolution of atheroma as opposed to the response to vascular injury. Because a paucity of VSMCs in the neointima contributes to the development of vulnerable plaques, we sought to characterize the impact of VSMC over-expression of PAI-1 on plaque morphology rather than on the size of the atherosclerotic lesion.
We generated transgenic mice in which VSMC expression of PAI-1 was increased with the use of the SM22α promoter (SM22-PAI+). Migration of VSMCs explanted from SM22-PAI-positive and -negative littermates through an artificial basement membrane matrix (Matrigel) was determined in vitro. Neointimal cellularity was quantified in ApoE−-/– mice that were either positive or negative with respect to the SM22-PAI transgene.
Materials and Methods
Generation of Transgenic Mice
All animals were used in protocols approved by the Institutional Animal Care and Use Committee at the University of Vermont. A chimeric gene designed to induce overexpression of PAI-1 in VSMCs was constructed with the use of p-Bluescript II SK(+) (Stratagene; La Jolla, CA). The multiple cloning site was replaced with that from PBK-CMV (Stratagene). A 400-bp segment of 5’ flanking DNA from SM-22α (–441 to +41) was subcloned into the multiple cloning site of PBK-CMV (in the p-Bluescript vector) (Solway et al. 1995). Next, murine PAI-1 cDNA (3.0 kb) was subcloned downstream of the mouse SM-22α promoter and a 2.1-kb human growth hormone intron/polyadenylation signal was subcloned downstream of the murine PAI-1 cDNA. The resulting chimeric gene (SM22-PAI) containing the SM-22α promoter, murine PAI-1 cDNA, and human growth hormone polyadenylation signal was isolated with NOT-1 restriction enzyme digestion. Orientation and insertion of chimeric components were confirmed with the use of restriction enzyme digestion. This fragment was introduced into B6C34Fl fertilized mouse oocytes via microinjection as described previously (Rincón et al. 1998).
The presence of the transgene was determined with DNA isolated from tail biopsies with the use of slot-blot analysis or the polymerase chain reaction (PCR). Slot-blot membranes were hybridized with a 500-bp fragment of human growth hormone cDNA. PCR was performed with the use of a primer in the PAI-1 cDNA (5'-TGAATGAATTTCCTAGCAGG-3') and a primer in the human growth hormone polyadenylation signal (5'-GAGAAACAGAGGGCAACAG-3') (GIBCO; Grand Island, NY). PCR conditions were as follows: 94C (1 min), 30 cycles 94C (30 sec), 60C (60 sec), 72C (2 min), 72C (10 min).
Animals with the transgene (SM22-PAI founders) were backcrossed into the C57Bl/6 line (Jackson Labs; Bar Harbor, ME). The in vitro comparative analyses were performed with the use of transgene-positive and -negative littermates fed a standard low-fat (0.28% cholesterol) diet. SM22-PAI+ mice that were congenic in the C57Bl/6 line were crossed with ApoE−-/– mice to generate ApoE−-/–SM22-PAI+ mice. ApoE−-/– and ApoE−-/–SM22-PAI+ mice were fed a high-fat (20% fat, 1.5% cholesterol, Teklad) diet after weaning. Neointimal composition was characterized in 20-week-old male ApoE−-/– and ApoE−-/–SM22-PAI+ mice. We elected to study male mice to limit variability that might occur as a result of gender, estrous cycle, or hormonal status.
Isolation of Aortas and VSMCs
Animals were euthanized with an IP injection of sodium pentobarbital. Hearts and aortas were flushed clear of blood by perfused PBS until blanching of the liver was noted and were then fixed in paraformaldehyde. Descending aortas were placed in sterile, cold (4C) Hank's balanced salt solution with 100 μg/ml streptomycin plus 300 U/ml penicillin, freed from adherent fat and connective tissue, cut into 1.0-mm3 pieces, placed on the etched surface of a six-well culture dish, and covered with sterile 22 × 22-mm glass cover-slips anchored with sterile silicon grease (Dow Corning high vacuum grease, autoclaved). Explants were cultured in 3 ml/wells in complete medium [Dulbecco's modified Eagle's medium (DMEM) supplemented with 20% fetal bovine serum (FBS), 100 U/ml penicillin, and 100 μg/ml streptomycin (GIBCO)] at 37C in room air supplemented with 10% CO2. Cells were propagated after dispersion with 0.25% trypsin in DMEM with 20% fetal bovine serum, 10 ng/ml epidermal growth factor, 50 U/ml penicillin, 50 μg/ml streptomycin, and 1 × ITS (10 ng/ml insulin, 0.55 ng/ml transferrin, 0.67 μg/ml selenium; GIBCO) at 37C in room air enriched with 10% CO2. Experiments were performed with VSMCs harvested from animals 7.5–30 weeks of age and were cultured for 3–8 passages. VSMC phenotype was verified by expression of α-smooth muscle actin.
Assessment of Cell Migration
Cell migration in vitro was quantified with a Biocoat insert system (Becton Dickinson; Bedford, MA) with the use of 24-well plates and inserts precoated with Matrigel (Becton Dickinson), an artificial basement membrane matrix that contains basement membrane structural components, enzymes, inhibitors, and growth factors. Inserts contained 1–2-μm-thick matrix on a polyethylene tetrachloride membrane perforated with 8-μm pores. Matrigel was hydrated with DMEM for 2 hr before cell seeding. Cells were cultured in DMEM with 1% FBS. Rat platelet-derived growth factor-BB (100 ng/ml; Sigma, St Louis, MO) was used as a chemoattractant in the lower chamber. Migration was determined 20 hr after the seeding of 5 × 104 VSMCs/ insert. VSMCs were cultured in DMEM without serum for 24 hr before addition to the chamber. Experiments were performed with fresh DMEM supplemented with 4% FBS. After 20 hr, Matrigel and cells were disrupted with a spatula on the upper aspects of the inserts and removed by aspiration. Cells were fixed in 70% methanol (–20C for 20 min), rinsed with PBS, and stained in a solution containing propidium iodide (20 μg/ml) and RNase A (200 μg/ml) (Sigma). Culture membranes were sharply separated from inserts and mounted on glass microscope slides in 50% PBS and glycerol.
The slides were evaluated with a laser scanning cytometer (CompuCyte; Cambridge, MA) (Taatjes et al. 2001). In each case, the entire insert mounted on the slide was scanned with a ×20 objective lens with the use of an argon ion laser with an orange/red fluorescence detector. The threshold contour was set based on the orange/red fluorescence intensity, with a minimum area of 10 μm2 required to trigger an event. Scanned cells depicted by points on a scattergram were randomly selected and visually relocated with the microscope component of the laser scanning cytometer to confirm their identity as whole cells rather than cell debris.
Immunohistochemical Analysis
Tissue Processing. Aortic tissue was processed for immunofluorescence essentially as described previously (Taatjes et al. 2000). In brief, the hearts and aortas were excised, placed in freshly prepared 3% formaldehyde (in PBS, pH 7.3), fixed overnight at 4C, placed in plastic embedding cassettes, and rinsed for 1 hr with cold running water. The cassettes were transferred into 5% gelatin (Type A from porcine skin; 300 bloom; Sigma) in distilled water for 2 hr at 42C, followed by infiltration in 10% gelatin in distilled water overnight at 42C, refrigerated for 3 hr (4C) to solidify the gelatin, trimmed rectangularly, and immersed immediately in 3% formaldehyde in PBS for 1 hr at 4C to crosslink and strengthen the gelatin. The capsules were then rinsed twice for 15 min each with PBS before freezing. The hearts were then embedded in OCT medium (Tissue-Tek; Sakura Finetek USA, Torrance, CA) in disposable cryostat embedding molds with the aortic arch side of the heart facing down, frozen by plunging into liquid nitrogen-cooled 2-methyl-butane, and stored at –80C until cryosectioning.
Cryostat Sectioning. The frozen hearts were removed from the embedding molds and attached to a cryostat chuck, flat side (arch side of the heart) down. Sixteen-μm-thick sections were collected from a 300-μm area beginning at the valve leaflets. We evaluated five 16-μm-thick aortic sections from each mouse for each component analyzed by quantitative microscopy.
Immunohistochemical Staining. After being rinsed in PBS containing 1% bovine serum albumin (PBS/BSA), the sections were incubated in 10% normal goat serum for 30 min, followed by an overnight incubation at 4C with a polyclonal rabbit anti-human PAI-1 antibody (courtesy of Drs. Lieve Moons and Desire Collen). After three PBS/BSA rinses, sections were incubated with a Cy5-conjugated goat anti-rabbit secondary antibody for 60 min. After a final rinse with PBS/BSA, a coverslip was applied with the use of 1% n-propyl gallate in 50% glycerol/50% PBS as an anti-fade mounting medium. In control incubations, the sections were incubated with secondary antibody alone (i.e., without primary antibody). VSMCs explanted from SM22-PAI-positive and -negative littermates were cultured on coverslips and stained in the same manner. Specificity of the antibody was confirmed by Western blotting analysis (Figure 1). The rabbit polyclonal antibody recognized both free and complexed PAI-1. To determine the identity of cells expressing PAI-1, cryostat sections were doubly stained with the anti-PAI-1 antibody and a monoclonal anti-smooth muscle actin antibody, rinsed with PBS/BSA for 10 min, and blocked with 10% normal goat serum for 30 min. After a 5-min rinse with PBS/BSA, the sections were incubated with 10% normal donkey serum for 30 min, followed by a 5-min PBS/BSA rinse. The sections were incubated overnight at 4C with rabbit anti-human PAI-1 (final concentration 15 mg/ml) and a murine anti-α-smooth muscle actin (clone αsm-1; Boehringer Mannheim, Indianapolis, IN; final concentration 5 mg/ml). After three 10-min rinses with PBS/BSA, sections were incubated for 60 min at RT with a mixture of goat anti-rabbit IgG conjugated to Cy5 (final concentration 10 mg/ml) and donkey anti-mouse IgG conjugated to Alexa 568 (final concentration 10 mg/ml), rinsed in PBS/BSA, stained with SYTOX Green (Molecular Probes, Eugene, OR; 5 mM in PBS) for 15 min at RT, as previously described (Taatjes et al. 2000), and rinsed in PBS/BSA. Coverslips were applied with 1% n-propyl gallate in 50% glycerol/50% PBS as an anti-fade mounting medium. For control incubations, the primary antibody was replaced by PBS/BSA.
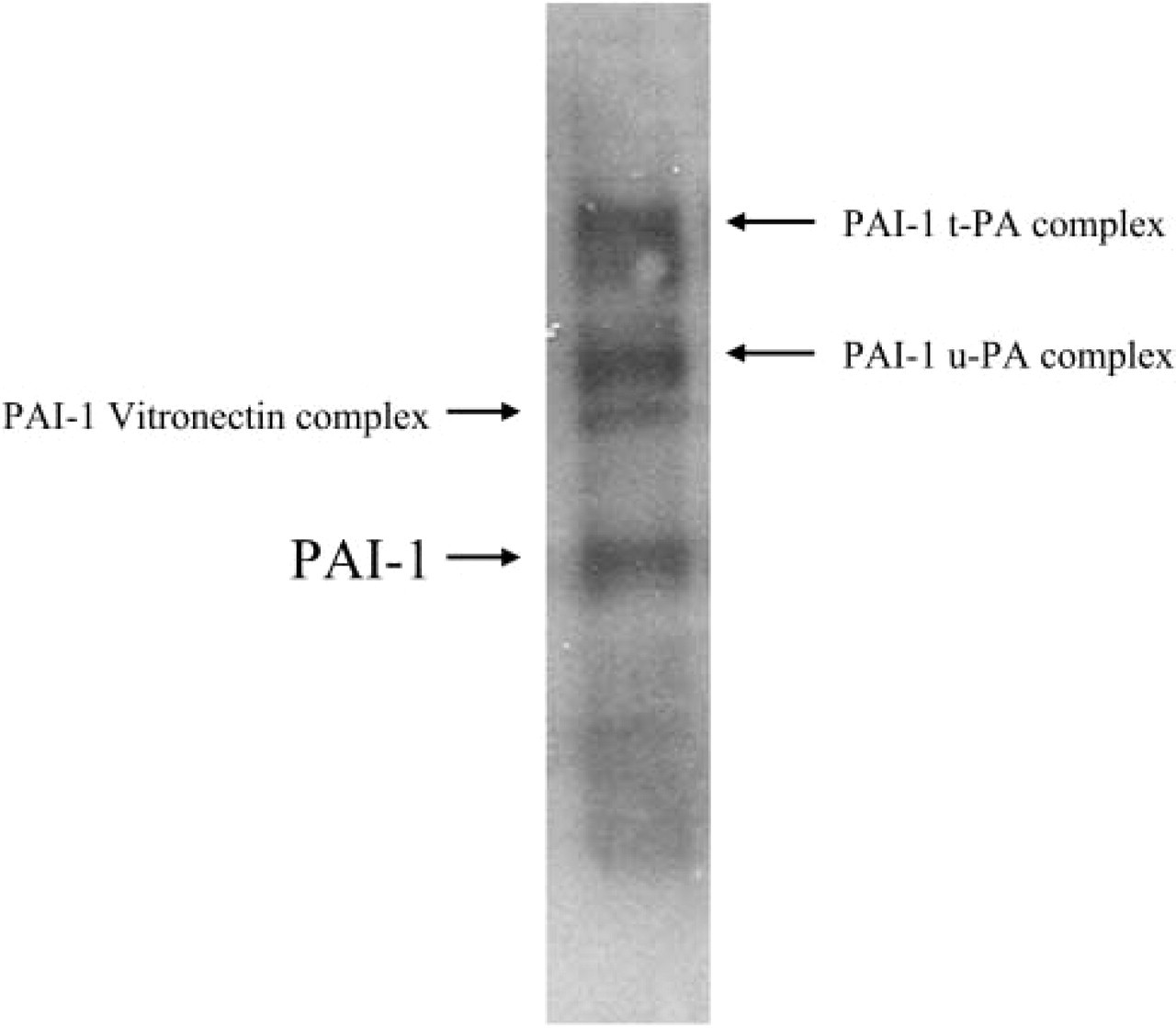
Specificity of the polyclonal rabbit anti-PAI-1 antibody. Ten μg of protein extracted from VSMCs that were explanted from a C57Bl/6 mouse was separated with the use of SDS-10% polyacrylamide gel electrophoresis. Proteins were transferred to a polyvinylidene difluoride membrane and probed with the rabbit anti-PAI-1 polyclonal antibody. Blots were washed and incubated with a secondary horseradish peroxidase-conjugated anti-rabbit IgG (Sigma) for 1 hr, then processed for antibody visualization with enhanced chemiluminescence detection reagents (Boehringer Mannheim) and imaged on Biomax film (Eastman Kodak; Rochester, NY). Free PAI-1 as well as PAI-1 complexed with vitronectin, urokinase-type plasminogen activator (u-PA), and tissue-type plasminogen activator (t-PA) were detected.
Confocal Scanning Laser Microscopy. The stained sections were examined with a Bio-Rad MRC 1024ES confocal scanning laser microscopy system (Bio-Rad Laboratories, Life Sciences Division; Hercules, CA). Immunostaining with PAI-1 antibody was visualized by excitation with the 647-nm wavelength line from a krypton/argon laser. For semiquantitative analysis, all images were captured with a ×20 objective lens (0.50 NA) and the following settings: image box size 512 × 512 pixels; laser power 30%; confocal iris 2.5 mm; gain 1250 V; black level –2; low signal, slow scan mode; and Kalman averaging ×3. Images were archived on 100 Mbyte Zip disks, and photographic prints made with a Fujix Pictrography 3000 video color printer.
Dual antibody-stained images were acquired with the use of sequential capture mode to avoid potential fluorescence bleed-through between channels. Images were captured in sequential mode in the following manner. The 488-nm laser line was used to excite SYTOX Green fluorescence (to label all cell nuclei for orientation purposes), the 568-nm laser line was used to excite the Alexa 568 fluorescence (indicating staining for α-smooth muscle actin antibody) and the 647-nm laser line was used to excite the Cy5 fluorescence (indicating staining with the anti-PAI-1 antibody). The three grayscale images were electronically merged to produce a pseudocolored image in which green depicts cell nuclei, red depicts PAI-1 immunoreactivity, and blue depicts smooth muscle actin staining.
Quantitative Analysis of PAI-1 Immunoreactivity. Analyses were performed by personnel who were blinded with respect to the genotype of the mice. Grayscale images acquired in proprietary BioRad.pic format were imported into Meta-Morph image analysis software (version 4.5r4; Universal Imaging, Downingtown, PA). The opened image was first calibrated for pixel size by applying a previously created calibration file. Segmented region areas, 40 × 40 pixel dimension, were applied to five randomly selected areas within the aortic media. A threshold encompassing an intensity range of 100–250 grayscale values was applied to the segmented regions of interest. The mean integrated pixel values from the five areas per animal were calculated and data converted into a graphic bar chart for comparative analysis. The concentration of PAI-1 in protein extracted from VSMCs was assessed also with the use of an ELISA assay (MPAIKT-TOT kit; Innovative Research, Southfield, MI).
Vascular Lesion Compositional Analysis
Lipid was stained with oil red O (Pearse method). DAPI (4',6-diamidine-2-phenylindole dihydrochloride), a nucleic acid-binding fluorescent dye (Roche Diagnostics; Mannheim, Germany) was used to delineate cellularity. Stained sections were examined with an Olympus BX50 upright light microscope (Olympus America; Lake Success, NY) with an attached Optronics MagnaFire digital camera (Optical Analysis; Nashua, NH). Images were acquired with the use of MagnaFire software (version 2.1B).
Computer-assisted Quantitative Image Processing and Analysis
The captured digital images were transferred to a Dell 400 NT workstation for cropping and processing to yield 24-bit RGB images per section isolated on a white background as previously described (Taatjes et al. 2000). The processed images were opened in MetaMorph Image Analysis software for analysis. The images were opened as a group for the component of interest and calibrated for the objective lens used during image collection. Appropriate color thresholding parameters were applied.
Qualitative Image Analysis
An independent observer blinded with respect to mouse genotype and other analyses performed qualitative assessments. Cropped images that were used also for quantitative analysis described above were used for this purpose. A qualitative score with a scale of 1 to 5 describing the extent of cellularity and lipid content was assigned to each lesion. Qualitative analysis was performed by comparison of lesions with a set of reference images that had been analyzed quantitatively. A score of 1 reflected the lowest extent of cellularity or lipid content and 5 the greatest. Each increment reflected an approximate increase of 20%.
Statistical Analysis
Values are means ± SD. Significance, defined as p<0.05, was determined with the use of Student's t-tests.
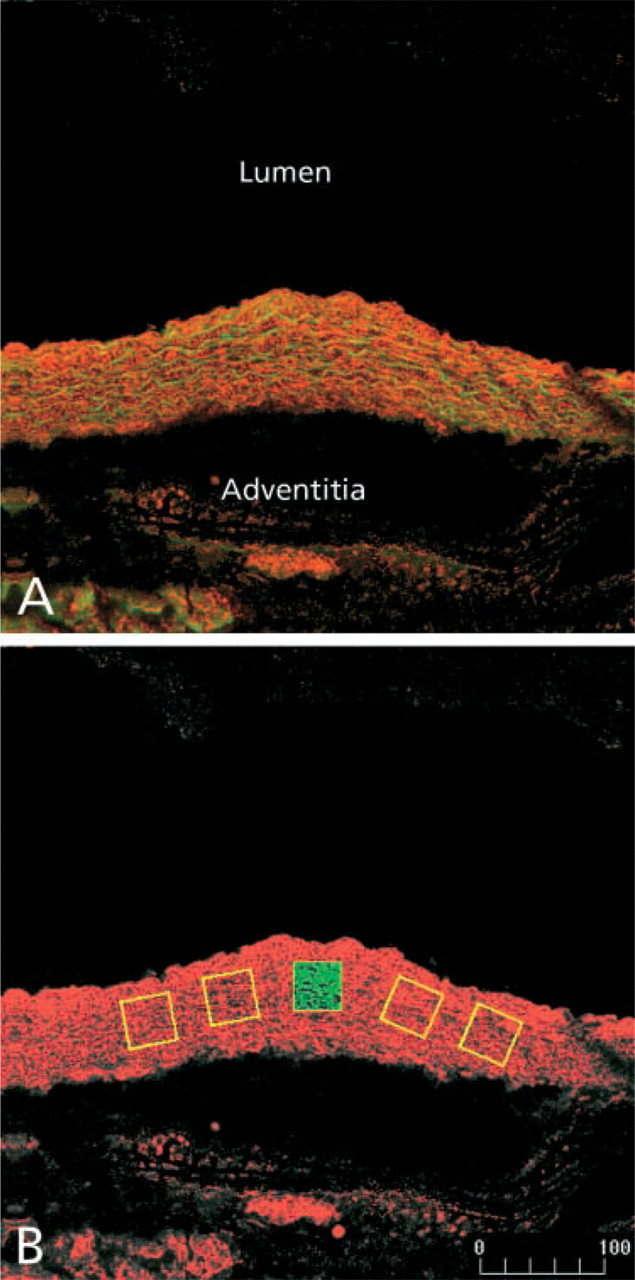
The method used for detection of PAI-1 protein with confocal scanning laser microscopy. Cryostat sections were stained with an anti-PAI-1 polyclonal antibody followed by a Cy5-conjugated secondary antibody. The stained sections were imaged with confocal scanning laser microscopy. Excitation of the Cy5 fluorophore was achieved with the 647-nm line of a krypton/argon laser. Autofluorescent emission from the elastic laminae was elicited by excitation with the 488-nm laser line. PAI-1 immunoreactivity was pseudocolored red and the autofluorescent elastic laminae green. (
Results
Expression of the SM22-PAI Transgene in C57Bl/6 Mice
Increased expression of PAI-1 protein was seen in the aortic wall from SM22-PAI+ mice that had been bred with C57Bl/6 mice compared with that in six gender-matched transgene-negative littermates that ranged in age from 7.5 to 30 weeks of age (Figures 2–4). Expression was compared in mice from F2 to F5 litters. Figure 2 describes the method used for quantitative comparison of PAI-1 immunoreactivity in the aortic wall. Cryostat sections were stained with an anti-PAI-1 polyclonal antibody followed by a Cy5-conjugated secondary antibody. Stained sections were imaged with the use of confocal scanning laser microscopy. Five regions were selected randomly (yellow boxes in Figure 2B) and the pixel values for each region were integrated to quantify expression of PAI-1.
Representative images of PAI-1 immunoreactivity in aortic vessel wall from both transgene-positive and -negative littermates are shown in Figure 3. Regardless of animal age, the transgene-positive animals always displayed more PAI-1 immunoreactivity than their transgene-negative counterparts. Figure 4 quantifies the intensity of immunostaining with the method described in Figure 3. The average intensity of immunostaining for PAI-1 was approximately threefold greater in the transgene-positive animals (p=0.01; Figure 4).
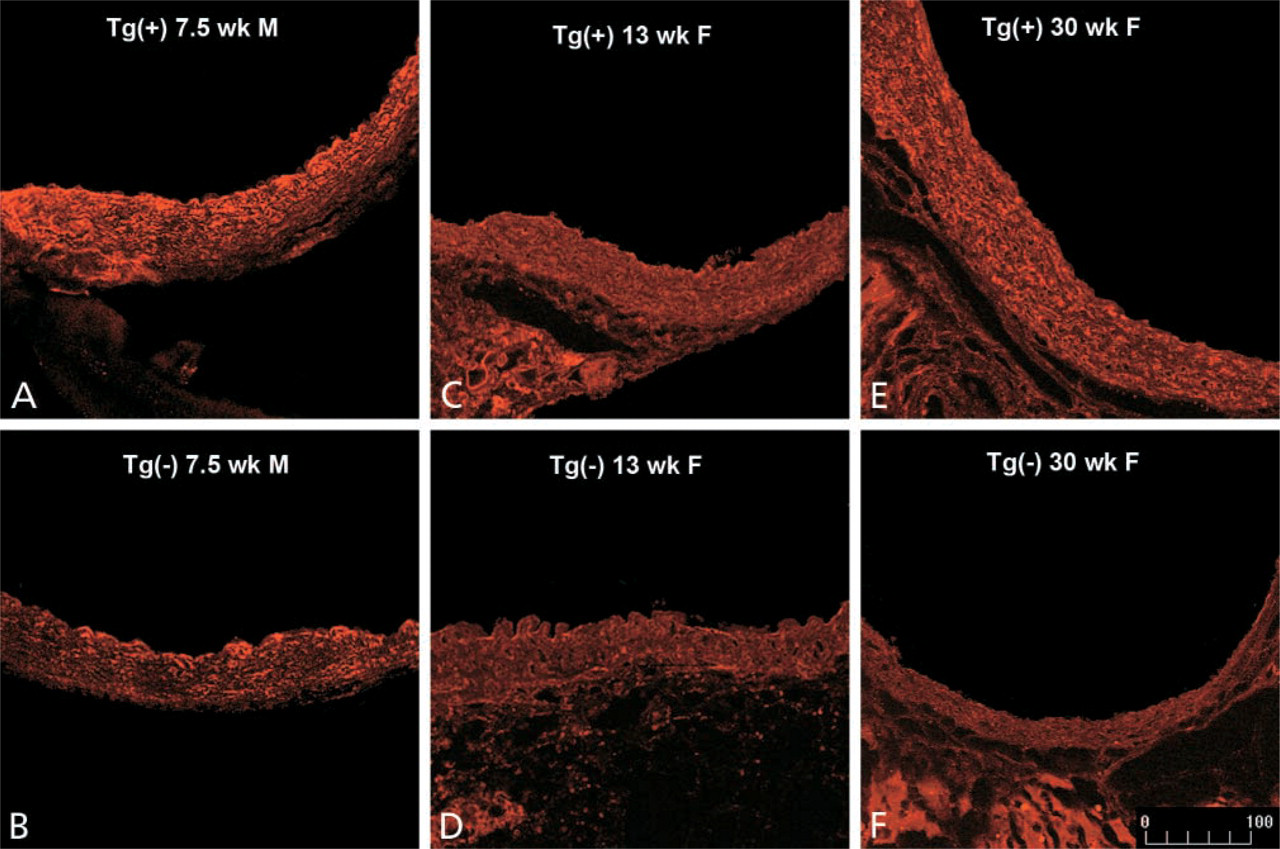
Detection of PAI-1 protein with confocal scanning laser microscopy in gender-matched SM22-PAI transgenic mice and negative control C57Bl/6 littermates. Immunoreactivity of PAI-1 was identified as described in the legend to Figure 2. Representative images from animals 7.5 weeks, 13 weeks, and 30 weeks of age are shown. Increased expression of PAI-1 protein was seen in transgenic (Tg+) animals (
To determine which cells expressed PAI-1, multiple-labeling confocal scanning laser microscopy was performed. Consistent with our previous work, Figure 5 demonstrates that cells expressing α-smooth muscle actin (indicative of VSMC origin) exhibited PAI-1 immunoreactivity (Taatjes et al. 1997). By contrast, immunohistochemical expression of PAI-1 in parenchymal tissue of the lung, heart, liver, and kidney from transgene-positive mice was not increased (data not shown). PAI-1 activity in blood was similar in both members of seven pairs of transgene-positive (16 ± 3 AU/ml) and -negative (17 ± 3 AU/ml) littermates. A second SM22-PAI founder line exhibited similar increases in aortic expression of PAI-1. SM22-PAI mice did not exhibit phenotypic alterations with respect to longevity and reproduction. Macroscopic and microscopic examination of aortas did not demonstrate morphological changes.
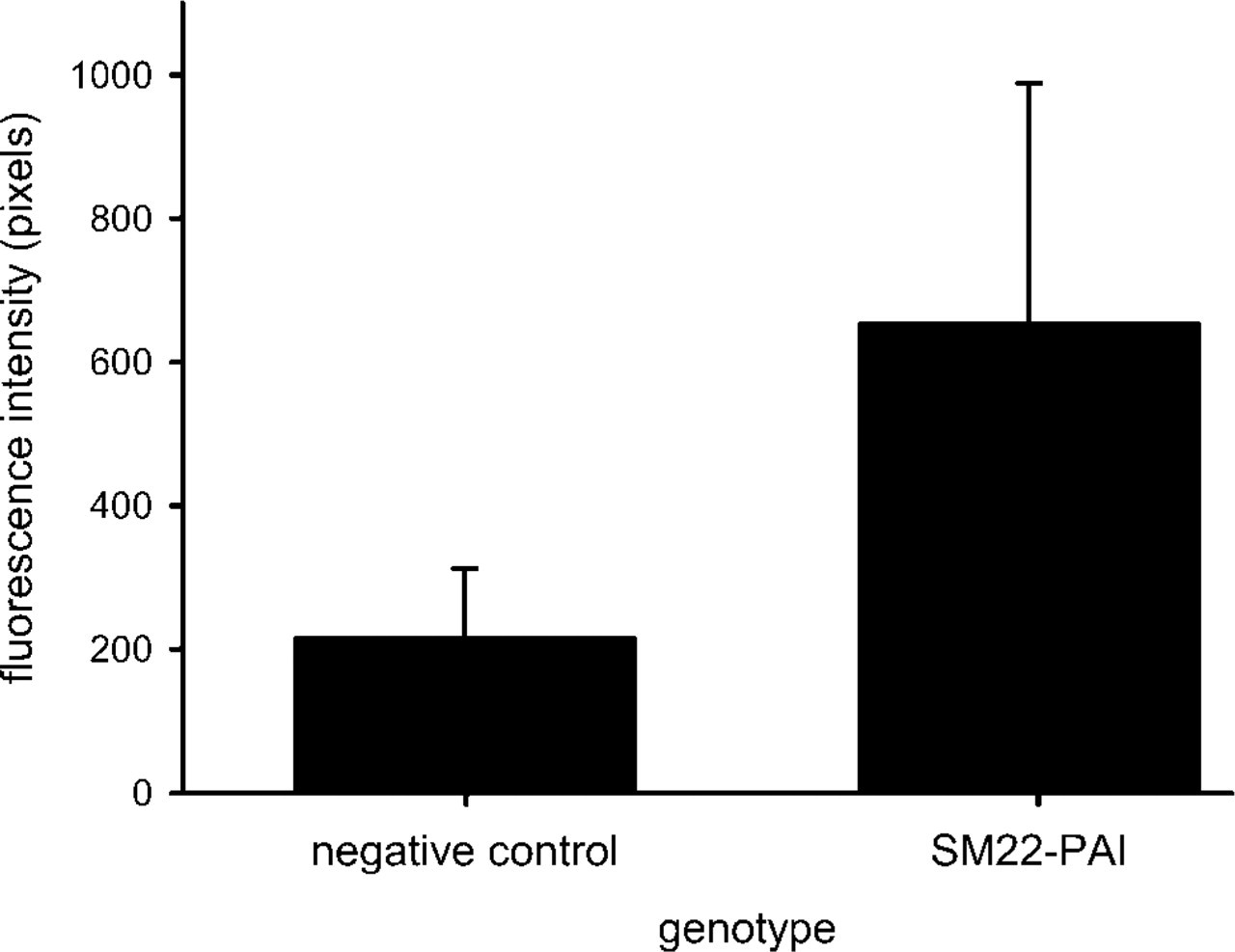
Average fluorescence intensity of PAI-1 protein in SM22-PAI transgenic mice and negative control C57Bl/6 littermates. Values are means ± SD of the integrated pixel intensity from six gender-matched littermates that ranged in age from 7.5 weeks to 30 weeks and included three pairs of female and three pairs of male littermates. The intensity of the immunofluorescence was quantified as described in Figure 1. A threefold increase in the protein expression of PAI-1 was seen (p=0.01).
VSMC Migration In Vitro
An approximately twofold increase in the immuno-histochemical expression of PAI-1 was exhibited by explanted VSMCs from SM22-PAI-positive mice compared with negative controls. Increased expression of PAI-1 protein by VSMCs from SM22-PAI+ mice was confirmed by determining the concentration of PAI-1 in 100 μg of protein extracted from explanted VSMCs. The protein was approximately twofold greater in SM22-PAI+ mice compared with that from negative control littermates (SM22-PAI+ = 8.6 ± 4.2, control = 3.6 ± 0.6; p=0.05; n=4). Their migration through Matrigel was diminished (Figure 6). Similar results were seen with VSMCs that had been explanted from three pairs of transgene-positive and -negative littermates.
Characteristics of Vascular Lesions in ApoE−-/– and ApoE−-/– SM22-PAI + Mice
The size of vascular lesions was similar in 20-week-old ApoE−-/– and ApoE−-/–SM22-PAI+ mice that had been fed a high-fat diet after weaning (Figure 7). The morphological characteristics of the neointima were assessed both qualitatively and quantitatively. Greater neointimal cellularity was apparent in aortas from ApoE−-/– mice with both the qualitative assessment (ApoE−-/– = 3.4 ± 0.2, ApoE−-/– SM22-PAI+ = 2.5 ± 0.6; p=0.01; n=5 for each) and the quantitative assessment (ApoE−-/– = 20.6 ± 2.6%, ApoE−-/– SM22-PAI+ = 16.4 ± 1.5%; p=0.01; n=5 for each; Figure 8). Lipid content was similar regardless of the SM22-PAI transgene status when assessed either qualitatively (ApoE−-/– = 4.2 ± 0.3, ApoE−-/– SM22-PAI+ = 3.8 ± 0.3) or quantitatively (ApoE−-/– = 56 ± 8%, ApoE−-/– SM22-PAI+ = 58 ± 4%).
Discussion
In previous work by others, ApoE−-/– transgenic mice generated to overexpress PAI-1 ubiquitously did not exhibit increased PAI-1 in the arterial wall (Sjoland et al. 2000). The transgenic mice that we developed with the SM-22α promoter exhibited increased aortic wall VSMC expression of PAI-1. This alone did not alter morphology of the vessel walls in the absence of vascular lesions. However, migration of explanted VSMCs in vitro was attentuated. Furthermore, neointimal cellularity of vascular lesions in ApoE−-/– mice was decreased in those with the SM22-PAI transgene. These results are consistent with our hypothesis that increased intramural PAI-1 promotes the evolution of atheroma particularly vulnerable to rupture and with the association between overexpression of PAI-1 in patients with diabetes and their high risk for acute coronary events (Sobel 1999; Sobel et al. 2003).
The objective of our study was to determine whether increased expression of PAI-1 altered the evolution of atheroma rather than a response to exogenous injury. This is because migration of VSMCs is likely to be pivotal during atherogenesis, whereas proliferation is potentially pivotal in response to exogenous injury. Nevertheless, our results are consistent with those of Carmeliet and colleagues, who found that mice in which expression of PAI-1 was increased exhibited decreased migration of VSMCs in response to electrically induced arterial mural injury (Carmeliet et al. 1997). By contrast, Lijnen and colleagues found that genetically increased expression of PAI-1 did not alter arterial VSMC content after severe electrical injury (Lijnen et al. 2004). Zhu and colleagues found that neointimal formation was greater in ApoE−-/– that were wild-type with respect to expression of PAI-1 compared with ApoE−-/– PAI-1−-/– mice when vascular injury was induced with ferric chloride (Zhu et al. 2001). Another important distinction between our study and the previous studies is the use of mice in our study in which the expression of PAI-1 was affected selectively rather than ubiquitously.
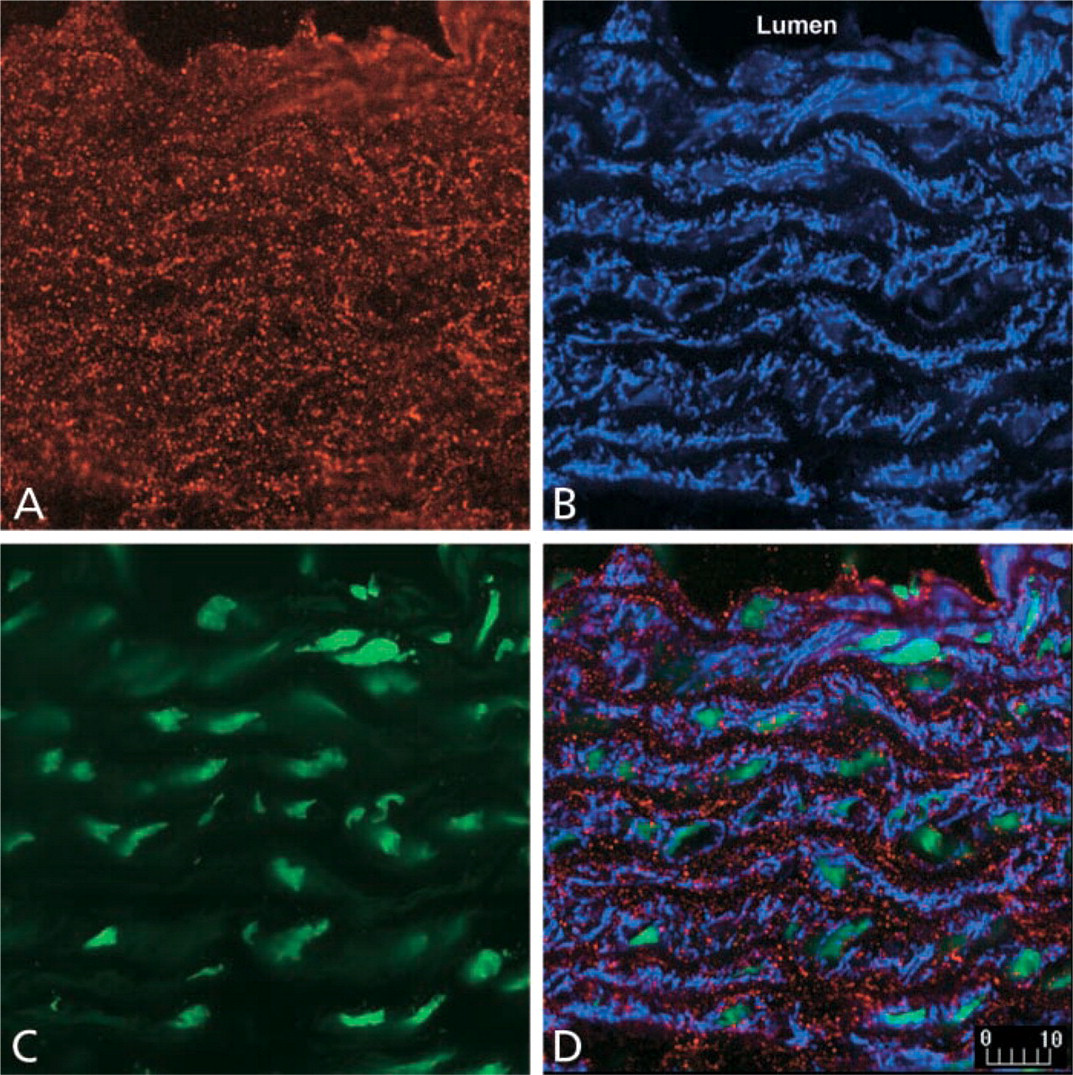
Expression of PAI-1 protein by smooth muscle cells in the aorta from SM22-PAI transgenic mice. Cryostat sections were incubated with antibodies against PAI-1 and α-smooth muscle actin as described in Materials and Methods. Single-channel images demonstrate PAI-1 immunoreactivity (
Our results are consistent with observations in diverse systems and with diverse cell types (O'Grady et al. 1981; Saksela and Rifkin 1988; Clowes et al. 1990; Redmond et al. 2001; Jackson and Reidy 1992; Bendeck et al. 1994; Stefansson and Lawrence 1996; Werb et al. 1997). Migration of human VSMC in vitro has been shown previously to be decreased by PAI-1 (Stefansson and Lawrence 1996; Redmond et al. 2001). Administration of a plasmin inhibitor, tranexamic acid, suppressed VSMC migration after injury of rat carotid arteries (Jackson and Reidy 1992). Activation of matrix metalloproteinases by plasmin generated from the ECM was inhibited by PAI-1 (O'Grady et al. 1981; Saksela and Rifkin 1988; Clowes et al. 1990; Bendeck et al. 1994; Werb et al. 1997), potentially constraining migration of many cell types.
Our results are consistent also with findings in patients that implicate PAI-1 in the potentiation of evolution of atherosclerotic plaques prone to rupture. Elevated concentrations of PAI-1 in blood are seen in patients with coronary artery disease, unstable angina, myocardial infarction, and stroke (Paramo et al. 1985; Juhan-Vague and Alessi 1993; Padro et al. 1995). Increased PAI-1 expression is evident in sections of atheroma (Schneiderman et al. 1992; Lupu et al. 1995; Padro et al. 1995). In people with diabetes the incidence of myocardial infarction, generally a complication of plaque rupture, is increased fourfold (Uusitupa et al. 1990). These patients have increased PAI-1 not only in blood but also in atheroma and arterial walls (Sobel et al. 1998; Pandolfi et al. 2001). Our results indicate that increased vessel wall expression of PAI-1 by VSMCs is likely to decrease their migration. Therefore, they are consistent with a predisposition towards the evolution of atherosclerotic plaques particularly prone to rupture in patients with increased VSMC expression of PAI-1 in vessel walls.
Diminution of increased intramural PAI-1 may favorably alter the evolution of atheroma with respect to cellular composition, thereby rendering them less vulnerable to rupture and less prone to precipitate an acute coronary syndrome. Diminution is an attractive target for prophylactic and therapeutic pharmacological intervention. Insulin sensitizers reduce elevated concentrations of PAI-1 in blood in patients with insulin resistance and those with type 2 diabetes mellitus. They attenuate evolution of atherogenesis, as judged from studies with duplex carotid ultrasound (Minamikawa et al. 1998; Koshiyama et al. 2001). Increased PAI-1 may inhibit migration of VSMCs disproportionately to macrophage and T-lymphocyte migration, thereby accounting for the observed close correlation between increased predominance of macrophages and increased PAI-1 in atheroma from patients with unstable angina or acute myocardial infarction compared with those from patients with stable angina pectoris (Shindo et al. 2001). A limitation of the current study is the absence of specific quantification of VSMCs and inflammatory cell content. Subsequent studies are planned to use confocal scanning laser microscopy to quantify the number of VSMCs, macrophages, and T-lymphocytes in vascular lesions of ApoE−-/– mice compared with ApoE−-/–SM22-PAI+ mice.
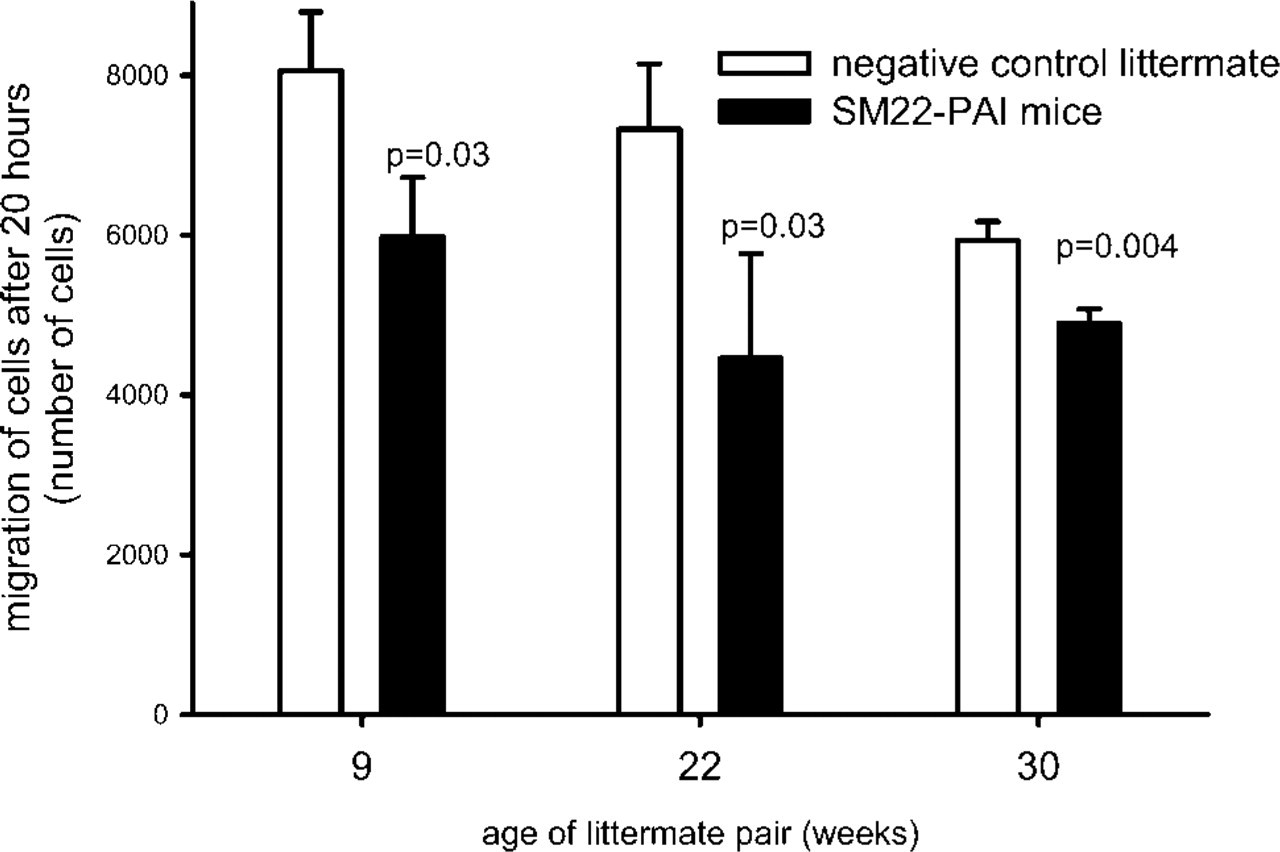
Migration of VSMCs through Matrigel over 20 hr. After hydration of the Matrigel membrane for 2 hr, 50,000 VSMCs were seeded into the upper chamber. Rat platelet-derived growth factor-BB (100 ng/ml) was added to the lower chamber as a chemoattractant. After 20 hr, Matrigel and cells were removed from the upper aspect of the membrane and VSMCs that had migrated to the basal aspect of the membrane were quantified with laser scanning cytometry. Results are the means ± SD from three wells each with VSMCs explanted from the aortas of SM22-PAI mice and negative control littermates. VSMCs had been maintained in culture for 4 (from 22- and 30-week littermates) through 7 (for 9-week littermates) passages. Migration of VSMCs from SM22-PAI mice was consistently attenuated.
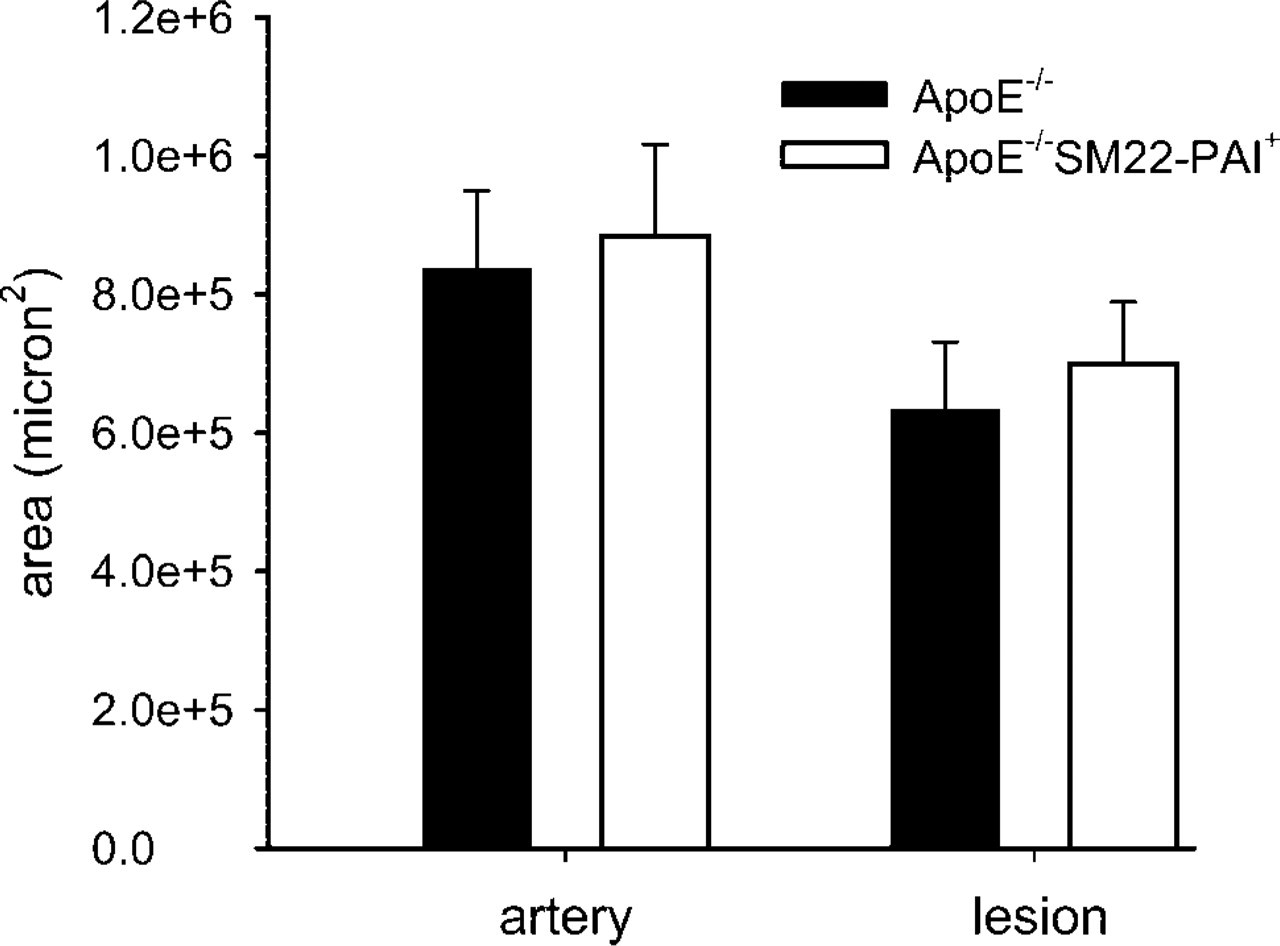
Artery and lesion size were assessed quantitatively with oil red O staining of cryostat sections. Both the average size of the artery and that of the lesion were similar in ApoE−-/– and ApoE−-/– SM22-PAI + male mice (n = 5 for each genotype) that were fed a high-fat diet after weaning and euthanized at 20 weeks of age.
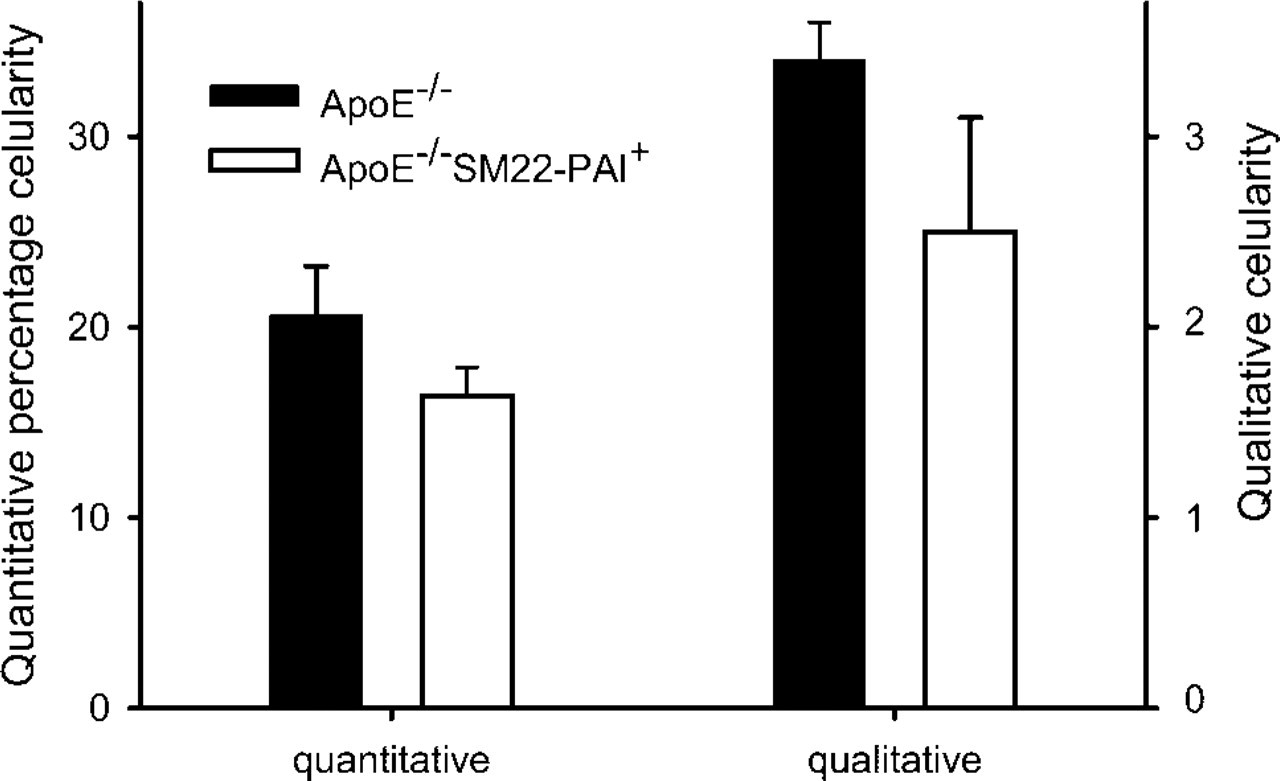
Neointimal cellularity of male ApoE−-/– and ApoE−-/– SM22-PAI+ mice were assessed both qualitatively and quantitatively. Mice (n = 5 for each genotype) were fed a high-fat diet after weaning and euthanized at 20 weeks of age. For the quantitative analysis, results are the percentage of the entire lesion occupied by cells identified by the staining of their nuclei. For qualitative analysis, lesions were compared with a set of reference images that had been analyzed quantitatively. A score of 1 reflected the lowest extent of cellularity or lipid content and 5 the greatest. Each increment reflected an approximate increase of 20%. Parallel results were obtained with qualitative and quantitative assessment. Greater cellularity was apparent in the ApoE−-/– mice (p=0.01 for both comparisons).
In summary, our results indicate that overexpression of PAI-1 by VSMCs reduces their capacity to migrate through the ECM and to populate the neointima. Therefore, overexpression of PAI-1 during the development of atherosclerotic lesions does not necessarily change the size of the lesion but may promote formation of a plaque that is more prone to rupture. Accordingly, inhibition of vessel wall PAI-1 overexpression may facilitate evolution of atherosclerotic plaques that are less prone to rupture and precipitate acute coronary events.