
Abstract
Brown adipose tissue (BAT) is richly provided with sympathetic noradrenergic nerves but is believed to lack a parasympathetic nerve supply. Acetylcholine is the predominant transmitter of postganglionic parasympathetic nerves. The vesicular acetylcholine transporter (VAChT) resides in synaptic vesicles of cholinergic nerve terminals and is used as a marker for peripheral cholinergic nerves. We sought cholinergic nerves in rat BAT using VAChT immunohistochemistry (IHC) on cryosections of interscapular, cervical, mediastinal, and perirenal depots. Mediastinal BAT was the sole depot provided with putative parasympathetic perivascular and parenchymal cholinergic nerves. The absence of vasoactive intestinal peptide-positive nerves suggested their nature as pure cholinergic fibers. By confocal microscopy, both cholinergic and noradrenergic nerves were detected in mediastinal BAT. Cold exposure and fasting led to increased density of VAChT-positive fibers and of noradrenergic sympathetic nerves at morphometry. The unexpected double innervation of mediastinal BAT may explain the inhibitory influence on thermogenesis observed after systemic injection of muscarinic antagonists in rats, and raises questions about the physiological role of its cholinergic nerve supply.
Keywords
I
Expression of the vesicular acetylcholine transporter (VAChT) has been demonstrated by in situ hybridization (ISH) in a variety of cholinergic neurons in the central and the peripheral nervous systems (Schafer et al. 1994). Subsequently, IHC studies have proved that VAChT is a reliable marker for the terminal fields of central and peripheral cholinergic nervous systems mainly by virtue of its high concentration in nerve terminals, which contain a number of small synaptic vesicles bearing this antigen (Schafer et al. 1995; Gilmor et al. 1996; Arvidsson et al. 1997; Schafer et al. 1998). Peripheral postganglionic cholinergic nerves are held to belong to the parasympathetic nervous system. The aim of the present work was therefore to establish whether rat subcutaneous and visceral BAT depots are provided with cholinergic, putatively parasympathetic, nerves. To address this question, we investigated by IHC the occurrence and distribution of VAChT-positive nerves in interscapular, cervical, mediastinal, and perirenal BAT depots of rats kept at different environmental temperatures and of fasted rats. The results showed that only mediastinal BAT is provided with cholinergic nerves and that they are modulated by both cold and fasting.
Materials and Methods
Animals and Tissues
Male Sprague-Dawley rats 9 weeks of age were obtained from Morini Laboratories (S. Polo d'Enza, Italy). They were singly caged and randomly assigned to the following four experimental groups: five rats with free access to food (65% carbohydrates, 11% fat, 24% protein, w/w) and water were kept at 22C (control group); five animals were kept at 4C for 2 days (cold-exposed group); five rats were kept at 28C for 10 days (warm-acclimated group), and five rats were fasted for 4 to 5 days (fasted group). Lights were on for 12 hr daily (0700 to 1900 hr). Animal care was in accordance with institutional guidelines.
All animals were anesthetized [100 mg/kg ketamine (Ketavet; Farm. Gellini, Aprilia, Italy) in combination with 10 mg/kg xylazine (Rompum; Bayer AG, Leverkusen, Germany)] and transcardially perfused with 4% paraformaldehyde in 0.1 M phosphate buffer, pH 7.4 (PB). Mediastinal, cervical, interscapular, and perirenal BAT and control tissues (see below) were dissected and postfixed by overnight immersion in the same fixative at 4C. After a brief wash in PB, tissues were cryoprotected in a solution of 30% sucrose in PB for 24 hr at 4C. Then a glass beaker containing 2-methylbutane (isopentane) was cooled in liquid nitrogen and the specimens, embedded in OCT medium, were frozen. Finally, tissues were stored at −80C.
Immunohistochemistry
Four-μm-thick cryosections obtained with a Leica CM1900 cryostat (Leica Microsystems; Vienna, Austria) were collected and air-dried overnight at room temperature (RT). Immunoreactivity was assessed according to the avidin-biotin-peroxidase (ABC) method. After two washes in 0.05 M PBS, pH 7.4, for 15 min each, sections were incubated with 0.3% H2O2 in methanol for 30 min at RT to block endogenous peroxidase, washed twice in PBS (15 min each), and incubated in 1:75 v/v normal goat serum [Vector Laboratories, Burlingame, CA; VAChT and vasoactive intestinal peptide (VIP) schedule] or in 1:75 v/v normal rabbit serum [Vector; tyrosine hydroxylase (TH) schedule] in PBS for 20 min at RT to block nonspecific sites. Sections were incubated overnight at 4C with the primary antibody against VAChT (polyclonal rabbit; Sigma–Aldrich, St Louis, MO) at a concentration of 1 μg/ml, with the primary antibody against VIP (polyclonal guinea pig; Peninsula Laboratories, San Carlos, CA) at a concentration of 2.5 μg/ml, or with the primary antibody against TH (polyclonal sheep; Chemicon International, Temecula, CA) at a concentration of 0.2 μg/ml; washed twice with PBS (15 min each), incubated in 1:200 v/v biotinylated IgG anti-rabbit goat serum (Vector; VAChT schedule), 1:200 v/v biotinylated IgG anti-guinea pig goat serum (Vector; VIP schedule), or 1:200 v/v biotinylated IgG anti-sheep rabbit serum (Vector; TH schedule) in PBS for 30 min at RT, washed twice with PBS (15 min each), incubated in ABC reagent (Vector) in PBS for 1 hr at RT, washed twice with PBS (15 min each) incubated in 0.02% H2O2 and 0.075% diaminobenzidine (Sigma) in 0.05 M Tris buffer (pH 7.6), kept for 5 min in a dark room, rinsed in distilled water, and counterstained with hematoxylin. Sections were finally dehydrated and mounted in Entellan. The ability of the antibodies to specifically detect the antigens was evaluated in sections (prepared as described above) of tissues known to contain the antigens (heart, intestine, and skeletal muscle for VAChT; central nervous system and intestine for VIP; central nervous system and heart for noradrenergic nerves). Negative controls were obtained in each instance by omitting the primary antibody and using preimmune instead of primary antiserum. The images were stored as TIFF files. Brightness and contrast of the final images were adjusted using the PHOTOSHOP 6 software (Adobe Systems; Mountain View, CA).
Morphometry and Statistical Analysis
For semiquantitative evaluations, VAChT and TH IHC reactions were performed under standardized conditions for all samples. The number of spots indicating specific immunostaining in parenchymal cholinergic and noradrenergic nerves and the area of these nerves were evaluated on three representative sections collected every 200 μm for each animal. Five rats were studied for each experimental condition, and an amount of mediastinal BAT containing ∼500 brown adipocytes was measured for each experimental condition. The number of nerve spots and the area (μm2) occupied by the specific brownish precipitate, indicating both presence and amount of VACht or TH, were measured at high power using a morphological imaging system (LUCIA, Version 4.5; Nikon Instruments, Rome, Italy). To compensate for the relative increase in nerve density due to cell shrinkage during fasting and, to a lesser extent, during cold exposure, results are presented as mean nerve spot number/50 brown adipocytes ± SE and as mean nerve area/50 brown adipocytes ± SE for each group. Group means were compared by two-way ANOVA. Significance was defined as 95% confidence levels.
Immunofluorescence and Confocal Microscopy
Immunofluorescence was performed on cryosections obtained according to the procedure described above. After two washes in PB for 15 min each, sections were incubated in 1:75 v/v normal donkey serum (Jackson ImmunoResearch; West Grove, PA) in PB for 20 min at RT to block nonspecific sites. Sections were incubated overnight at 4C with the mixture containing the primary antibody against VAChT (polyclonal rabbit; Sigma) at a concentration of 1.3 μg/ml and the primary antibody against TH (polyclonal sheep; Chemicon) at a concentration of 0.3 μg/ml, washed twice with PB (15 min each), and incubated in 1:100 v/v FITC donkey anti-rabbit (Jackson) and TRITC donkey anti-sheep (Jackson) antibody in PB for 30 min at RT. Sections were subsequently washed twice with PB, air-dried, and cover-slipped using the Vectashield mounting medium (Vector). Fluorescence was detected with a BioRad (Hercules, CA) Microradiance confocal laser scanning microscope equipped with an argon and He/Ne mixed gas laser. FITC and TRITC were excited with the 488- and 543-nm lines, respectively, imaged separately (emissions were separated using 515/30- and 570-nm filters), and then merged using the LaserSharp Processing Bio-Rad software (version 3.2). Sections were viewed in an Eclipse E600 Nikon microscope with a ×60 plan apochromat objective and 1.4 numerical aperture. The images (512 × 512 pixels) were then obtained sequentially from two channels using a confocal pinhole of 2–3. The images were stored as TIFF files. Brightness and contrast of the final images were adjusted using the Photoshop 6 software (Adobe Systems).
Results
Mediastinal BAT Is the Only Brown Fat Depot Provided with Cholinergic Nerves
In rats, mediastinal BAT is found in the upper and posterior mediastinum. It is composed of many different-sized lobules lying among the large mediastinal blood vessels, heart, trachea, esophagus, and descending aorta. In control rats, VAChT-positive nerves provided surrounding viscera. Occasionally they were also found in the mediastinal BAT lobules, sometimes around blood vessels, more often arteries (Figure 1A), but more frequently running among brown adipocytes in the parenchyma (Figure 1B). The density of VAChT-positive nerves varied among BAT lobules regardless of their size or anatomic position. The origin of these cholinergic nerves was probably the postganglionic neurons contained in the parasympathetic ganglia located close to the mediastinal viscera (Figure 2A). Cholinergic nerves usually co-localize VIP (Lindh and Hökfelt 1990). Nevertheless, with a specific antibody against VIP (Figure 2B), VIP-positive nerves were never found in mediastinal BAT in control rats or in any other experimental condition investigated (see below). Double labeling experiments in mediastinal BAT sections showed the presence of both cholinergic and noradrenergic nerves, which ran parallel and in close proximity to one another both around blood vessels and in the parenchyma under all conditions examined (Figure 3).
VAChT-positive nerves were never found in interscapular, cervical, and perirenal BAT depots in control animals or in any of the experimental conditions investigated (see below). Mediastinal BAT is therefore the only BAT depot in rats to be provided with both sympathetic noradrenergic and putatively parasympathetic cholinergic innervation.
Cholinergic Nerve Density Increases After Cold Exposure in Mediastinal BAT
To verify whether cold, the physiological stimulus for BAT heat production and recruitment (Trayhurn and Nicholls 1986), modulates the cholinergic nerve supply to mediastinal fat, we compared by IHC and morphological analysis the density of TH- and VAChT-positive nerves in mediastinal BAT in rats kept at temperatures close to thermoneutrality (28C) and in animals exposed to low temperatures (4C) for 2 days. As expected, TH-positive nerves were sparse in the former animals, whereas their density significantly increased after cold exposure (Figures 4A and 4B). Notably, very few vascular and parenchymal VAChT-positive nerves were found in mediastinal BAT in rats kept in a thermoneutral condition. After cold exposure, VAChT immunoreactivity was more evident at perivascular sites and the density of VAChT-positive nerves among brown adipocytes increased significantly (Figures 4A and 4B).
Cholinergic Nerve Density Increases During Fasting in the Mediastinal BAT
Fasting is associated with complex metabolic and morphological changes in BAT (Cinti 1999). Food reduction and starvation are generally believed to inhibit BAT heat production (Trayhurn and Jennings 1988), probably to save lipids for the general metabolism, but brown adipocytes may retain the ability to produce heat to maintain a constant body temperature under certain conditions (Champigny and Ricquier 1990). Compared with controls, rats fasted for 4 or 5 days (which showed a homogeneous weight reduction of ∼25 ± 1.5%) exhibited a more evident TH immunoreactivity around blood vessels and a significant increase in parenchymal noradrenergic nerve density (Figures 4C and 4D). In parallel, vascular and parenchymal VAChT-positive nerves increased, albeit not significantly, largely because of the extreme variability of their density among the lobules (Figures 4C and 4D).
Discussion
Among the four larger BAT depots of rats (interscapular, cervical, mediastinal, and perirenal), only mediastinal brown fat lobules are provided with cholinergic nerves. In this study we used VAChT to detect cholinergic nerves (Arvidsson et al. 1997). VAChT belongs to the family of vesicular transporters, whose function is to concentrate neurotransmitters into synaptic vesicles through proton exchange (Bravo and Parsons 2002). In cholinergic neurons, acetylcholine is synthesized in the cytoplasm of nerve endings by choline acetyl transferase and is transported by VAChT into synaptic vesicles, where it is stored until release (Prado et al. 2002). Therefore, by detecting the presynaptic pool of cholinergic vesicles, VAChT IHC allows visualization of the sites of neurotransmitter release along axons (Schafer et al. 1995; Gilmor et al. 1996; Arvidsson et al. 1997; Schafer et al. 1998). In mediastinal BAT, VAChT-positive nerves were observed at perivascular sites and, more often, among brown adipocytes. This pattern of immunostaining lends support to the hypothesis that acetylcholine is indeed released by both perivascular and parenchymal cholinergic nerves and that it exerts an effect on blood vessels and brown adipocytes.
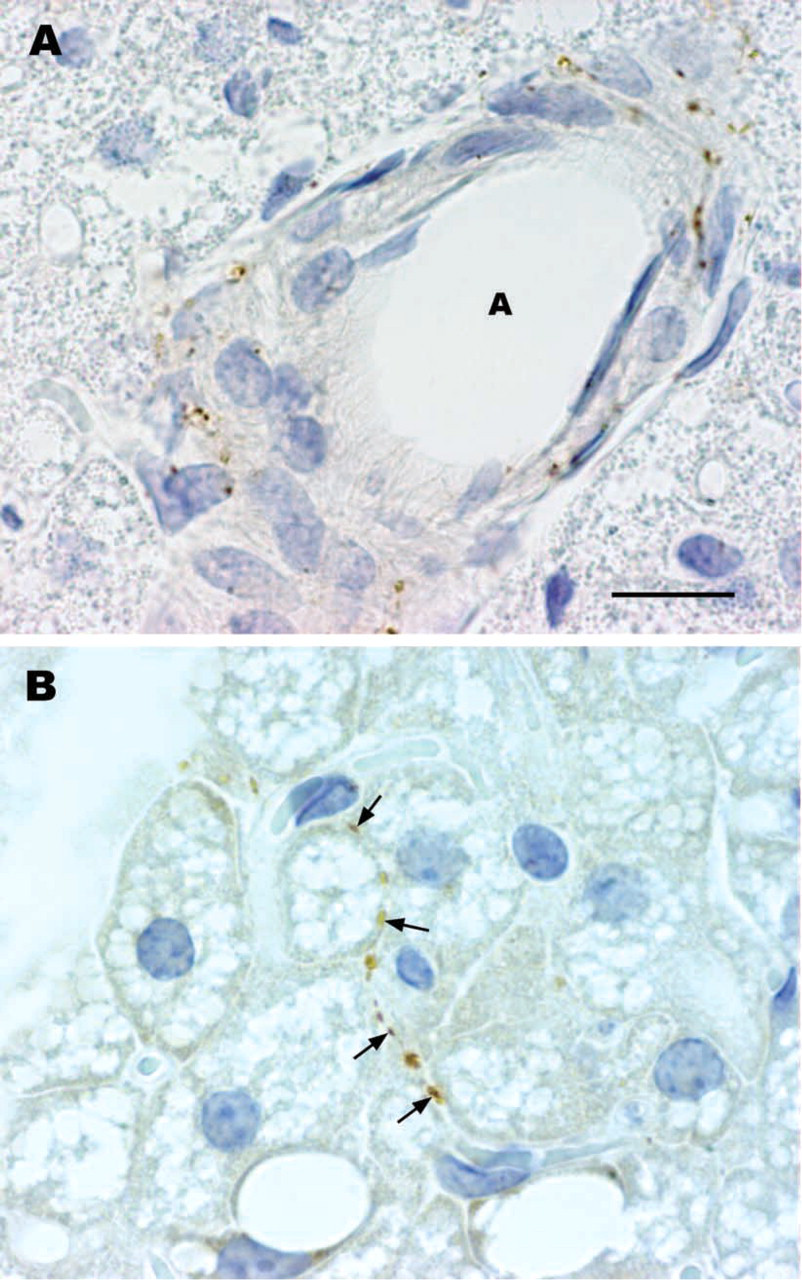
Vesicular acetylcholine transporter IHC in rat mediastinal brown adipose tissue. In control animals, cholinergic nerves are found in the adventitia of intralobular blood vessels (
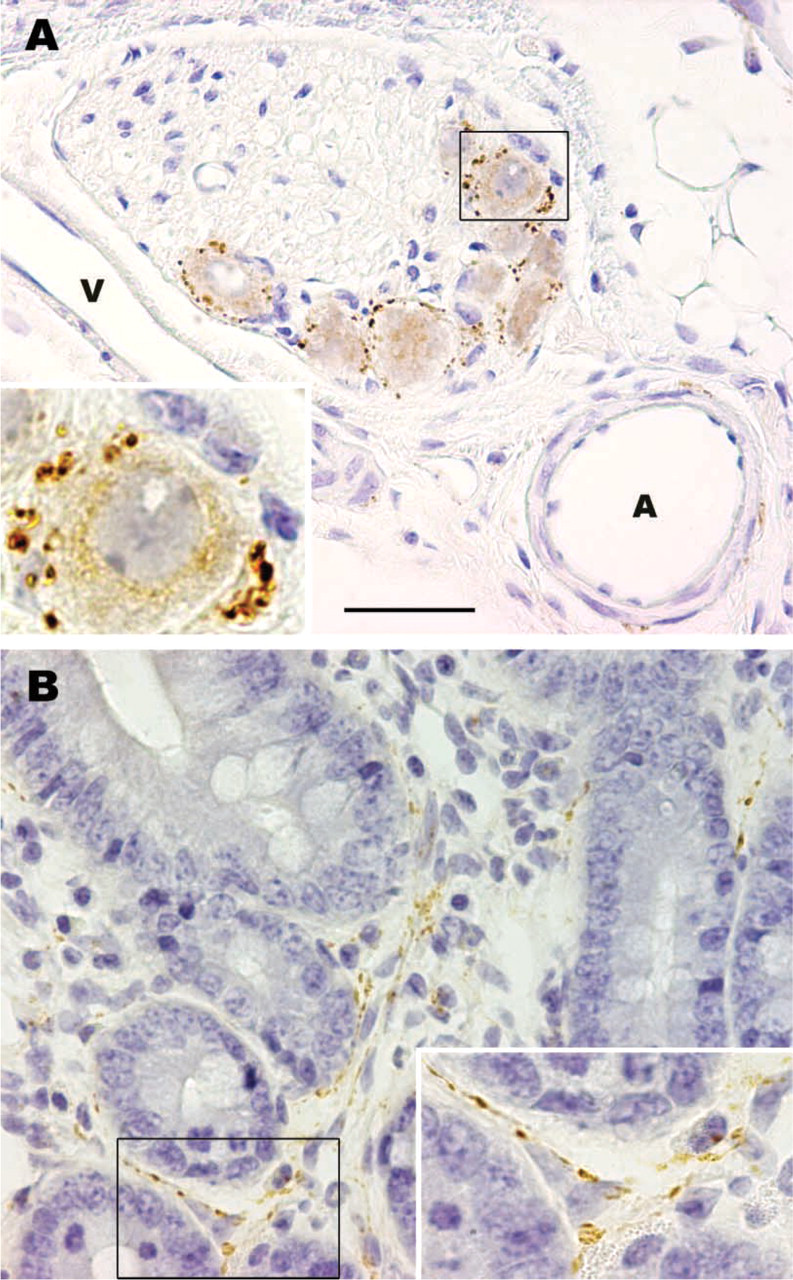
(
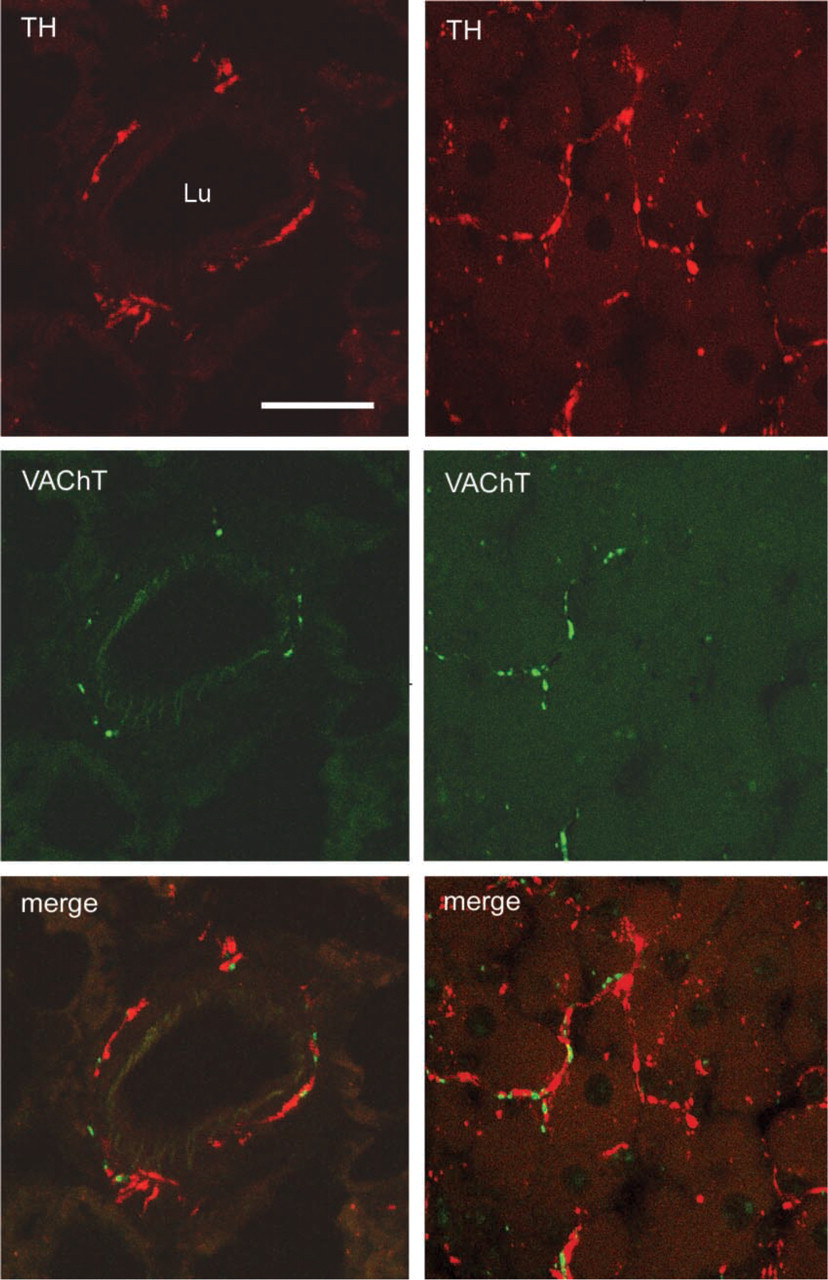
Indirect immunofluorescence in rat mediastinal brown adipose tissue. Confocal microscopy reveals the presence in the same section of both noradrenergic (TH, red) and cholinergic (VAChT, green) nerves at perivascular (left panels) and parenchymal (right panels) sites. Lu, lumen of an artery. Bar = 15 μm.
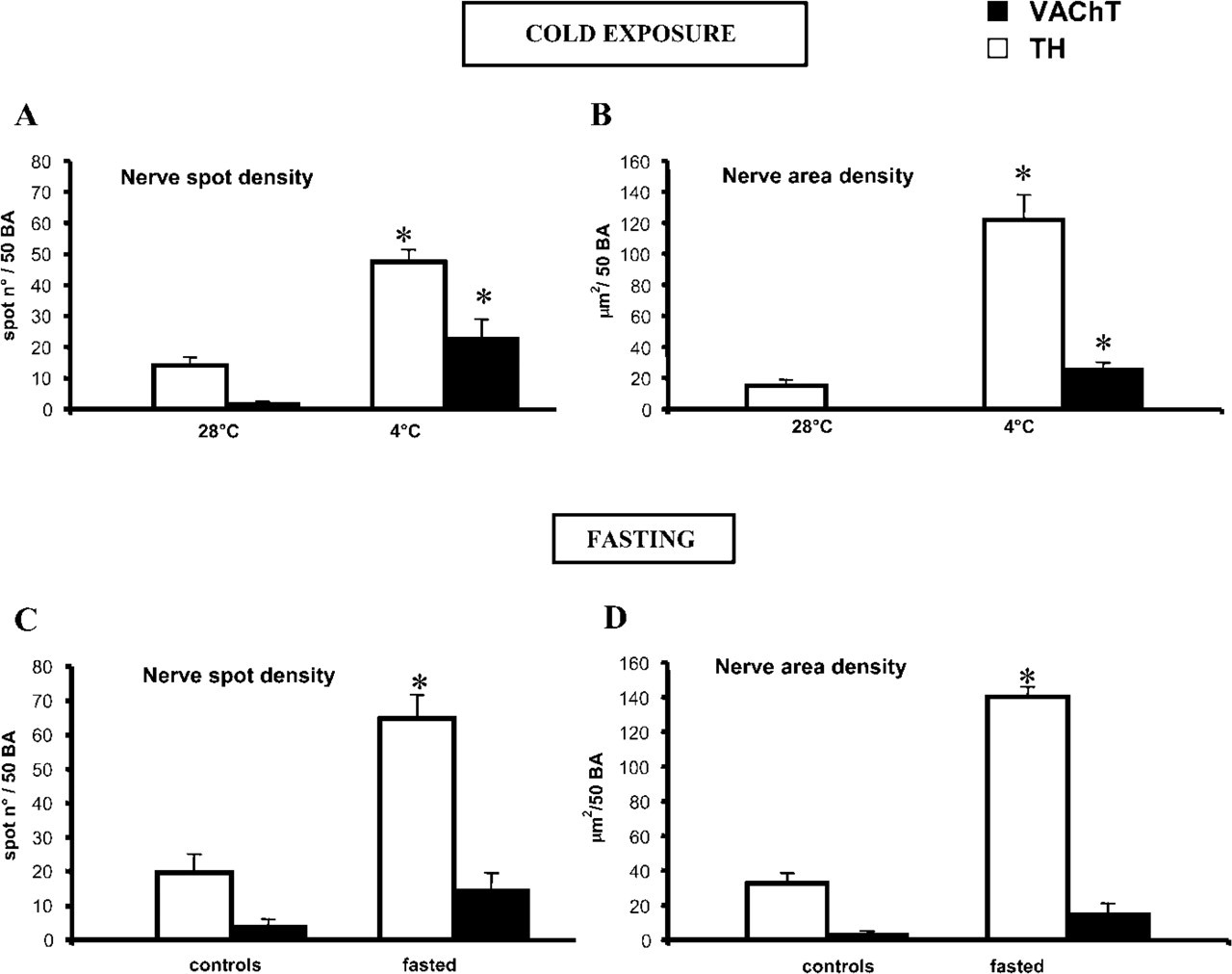
Mediastinal brown adipose tissue. Morphometry. (
A possible cholinergic control over mammalian thermogenesis has been hypothesized. In rats, injection of the muscarinic receptor antagonist atropine sulfate enhances the acute thermogenic response to a meal, an effect that is particularly evident in genetically obese Zucker rats, which normally show defective diet-induced thermogenesis (Rothwell et al. 1981). Furthermore, injection of atropine or surgical vagotomy also prevents metabolic rate decline after glucopenia induced by central or peripheral injections of 2-deoxy-
Cold exposure and fasting led to increased density of VAChT-positive fibers in addition to noradrenergic sympathetic nerves. Therefore, any influence of cholinergic nerves on brown adipocyte thermogenesis and metabolism increases during cold-dependent and fast-dependent heat production and/or lipid secretion.
Acetylcholine is usually found in the parasympathetic system throughout the body. Therefore, the cholinergic nerves found in mediastinal BAT are probably postganglionic parasympathetic nerves pertaining to the vagal nerve and arising from the many parasympathetic ganglia located close to the mediastinal viscera. Nevertheless, sympathetic postganglionic cholinergic nerves have been described in sweat glands (Landis and Keefe 1983; Leblanc and Landis 1986), arterial microvasculature of skeletal muscle (Bolme and Fuxe 1970), and periosteum (Asmus et al. 2000). Therefore, it cannot be excluded that the cholinergic axons supplying mediastinal brown fat lobules could derive from the cholinergic neurons of the sympathetic thoracic chain (Schafer et al. 1998).
We did not find cholinergic innervation in inter-scapular, cervical, and perirenal BAT depots. However, it should be noted that non-cholinergic nitric oxide-, VIP-, and/or calcitonin gene-related peptide-containing parasympathetic nerves have recently been described in human airways (van der Velden and Hulsmann 1999) and in rat heart (Onuoha et al. 1999). Therefore, our results do not exclude a non-cholinergic parasympathetic nerve supply to other BAT depots. Of note is that a dual sympathetic and parasympathetic innervation has recently been hypothesized for rat white adipose tissue (Kreier et al. 2002).
On the other hand, if further studies were to exclude the presence of a parasympathetic nerve supply to the other BAT depots, mediastinal BAT would be the sole BAT depot provided with both a sympathetic and a parasympathetic nerve supply. Such double control may be connected with a closer and more subtle modulation of brown adipocyte metabolism and heat production in the mediastinum, possibly in a still unknown relationship to cardiac activity.
Footnotes
Acknowledgements
Supported by grants from Polytechnic University of Marche (Contributi Ricerca Scientifica) and the Ministry of University (Cofinanziamento Bando 2002) to S.C. and M.C.