
Abstract
Cells in tendons are conventionally identified as elongated tenocytes and ovoid tenoblasts, but specific markers for these cells are not available. The roles and interplay of these cells in tendon growth, remodeling, and healing are not well established. Therefore, we proposed to characterize these cells with respect to cell turnover, extracellular matrix metabolism, and expression of growth factors. Here we examined 14 healthy human patellar tendon samples for the expression of various proteins in tenocytes and tenoblasts, which were identified as elongated tendon cells and ovoid tendon cells, respectively. Matrix metalloproteinase 1 (MMP1), procollagen type I (procol I), heat shock protein 47 (hsp47), bone morphogenetic protein 12 (BMP12), 13 (BMP13), and transforming growth factor β1 (TGFβ1) were detected by immunohistochemistry (IHC). An image analysis of the IHC staining for proliferation cell nuclear antigen (PCNA) and apoptotic cells was performed to determine the proliferation index and the apoptosis index in elongated and ovoid tendon cells. The ovoid tendon cells expressed higher levels of procol I, hsp47, MMP1, BMP12, BMP13, and TGFβ1 than the elongated tendon cells. Both the proliferation index and the apoptosis index of ovoid tendon cells were higher than those of the elongated tendon cells. The results suggested that ovoid tendon cells, conventionally recognized as tenoblasts, were more active in matrix remodeling. The expression of BMP 12, BMP13 and TGFβ1 might be associated with the different cellular activities of tenoblasts and tenocytes.
T
Only a few studies have focused on the organogenesis of tendons in early development. A transcription factor, scleraxis, is a highly specific marker for all the connective tissues that mediate attachment of muscle to bone in chick and mouse (Schweitzer et al. 2001), and the development of tendon may involve signaling of BMP12 (Lou et al. 1999) as well as FGF8 (Edom-Vovard et al. 2002). However, the differentiation process of tendon in adults has not been studied. Tenoblasts and tenocytes may represent different statuses of differentiation with distinct roles in the tendon healing process in adults. They may contribute to the inherent healing potentials of tendons and the development of pathological conditions such as tendinosis (Rolf et al. 2001).
Cells undergo proliferation and apoptosis during tissue differentiation. Therefore, the proliferation index and apoptosis index of tenoblasts may be higher than those of tenocytes, which are presumably terminally differentiated. In addition, tenoblasts and tenocytes may also exhibit different capacities in matrix remodeling. Matrix remodeling involves localized matrix formation and degradation, which is important to tendon healing, adaptation to exercise, and tendon growth. We propose that tenoblasts, identified as round tendon cells, are responsible for matrix remodeling in healthy tendons, in contrast to tenocytes.
Here we collected healthy patellar tendon samples from patients undergoing anterior cruciate ligament reconstruction for histological examination. An image analysis protocol was designed to distinguish tenocytes and tenoblasts according to cell shape, and the percentages of proliferating cells and apoptotic cells were measured. The expression of procollagen type I (Procol I), heat shock protein 47 (hsp47), matrix metalloproteinase 1 (MMP1), transforming growth factor beta 1 (TGFβ1), and bone morphogenetic protein 12 (BMP12) and 13 (BMP13) were detected by immunohistochemistry (IHC).
Materials and Methods
All sampling procedures were approved by the Human Research Ethics Committee of the authors’ institution.
Sample Collection and Preparation
All subjects and controls were recruited from the authors’ institution. Fourteen samples (10 male and 4 female, with an average age of 28, ranging from 18 to 41) from patients with anterior cruciate ligament deficiency undergoing bone-patellar tendon-bone autograft were included in the current study. The subjects had no previous history or clinical signs of patellar tendon injury. Subjects were informed of the procedures and provided consent before surgery. A 0.5 × 0.2 × 0.2-cm piece of healthy patellar tendon was removed from the patellar tendon during anterior cruciate ligament reconstruction. All samples were taken within the deep layers of the tendon in its central portion. All specimens were cleansed in sterile saline, fixed in buffered formalin, and then used to prepare 5-μm-thick paraffin-embedded sections mounted on 3-aminopropyl-triethoxy-silane (Sigma-Aldrich; St Louis, MO) and dried overnight at 40C.
Immunohistochemical Staining
IHC staining was performed as described previously (Fu et al. 2003). In brief, after removal of paraffin and dehydration, two consecutive paraffin sections from each sample were quenched with 3% H2O2 in methanol for 20 min and transferred to warm 10 mmol/liter citrate buffer solution (pH 6) for 10 min. After brief digestion with 0.05% trypsin for 5 min, the sections were incubated with primary antibody in a humid chamber at 4C overnight. The primary antibodies and the titers used are listed in Table 1. Negative staining controls were prepared by omitting the primary antibody.
After thorough washing with PBS, the sections were incubated with a secondary antibody, biotinylated anti-mouse/ rabbit IgG (DAKO; Glostrup, Denmark) or biotinylated anti-goat IgG (Chemicon; Temecula, CA). The classic antibody-binding complex method was used to amplify the specific binding signal. Finally, 3,3'-diaminobenzidine tetrahydrochloride (DAB) was used to develop color in the presence of H2O2. The sections were rinsed in distilled water, counterstained in Mayer's hematoxylin, dehydrated through graded alcohol to xylene, and mounted with p-xylene-bis-pyridinium bromide (DPX) permount (Sigma-Aldrich).
Antibodies used for immunohistochemistry
TdT-mediated dUTP Nick-end Labeling (TUNEL)
TUNEL identifies cells undergoing apoptosis by labeling nuclear DNA fragments that have been cleaved during apoptosis (Gavrieli et al. 1992). Apoptotic cells were detected on paraffin-embedded sections using an Apoptosis Detection Kit (Intergen; Purchase, NY). After removal of paraffin and rehydration of the sections, sections were pretreated with 20 μg/ml protease K (Sigma Chemical; St Louis, MO) and quenched with 3% H2O2 in PBS for 5 min. The fragmented DNA in the apoptotic cells was labeled with digoxigenin-11-dUTP by terminal deoxinucleotidyl transferase (TdT) for 2 hr. Negative controls were prepared by replacing the enzyme with reaction buffer only. The digoxigenin-labeled cells were treated with the peroxidase-conjugated anti-digoxigenin antibody for 30 min and diaminobenzidine (DAB) was added for color development (Ito et al. 1999). The sections were rinsed in distilled water, counterstained in Mayer's hematoxylin, dehydrated through graded alcohol to xylene, and mounted with p-xylene-bis-pyridinium bromide (DPX) permount (Sigma-Aldrich).
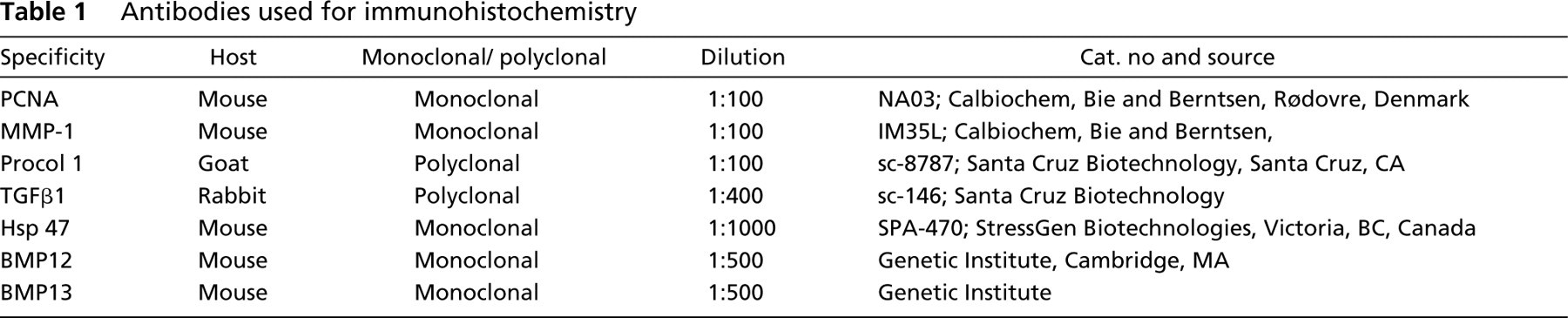
Round tendon cells (
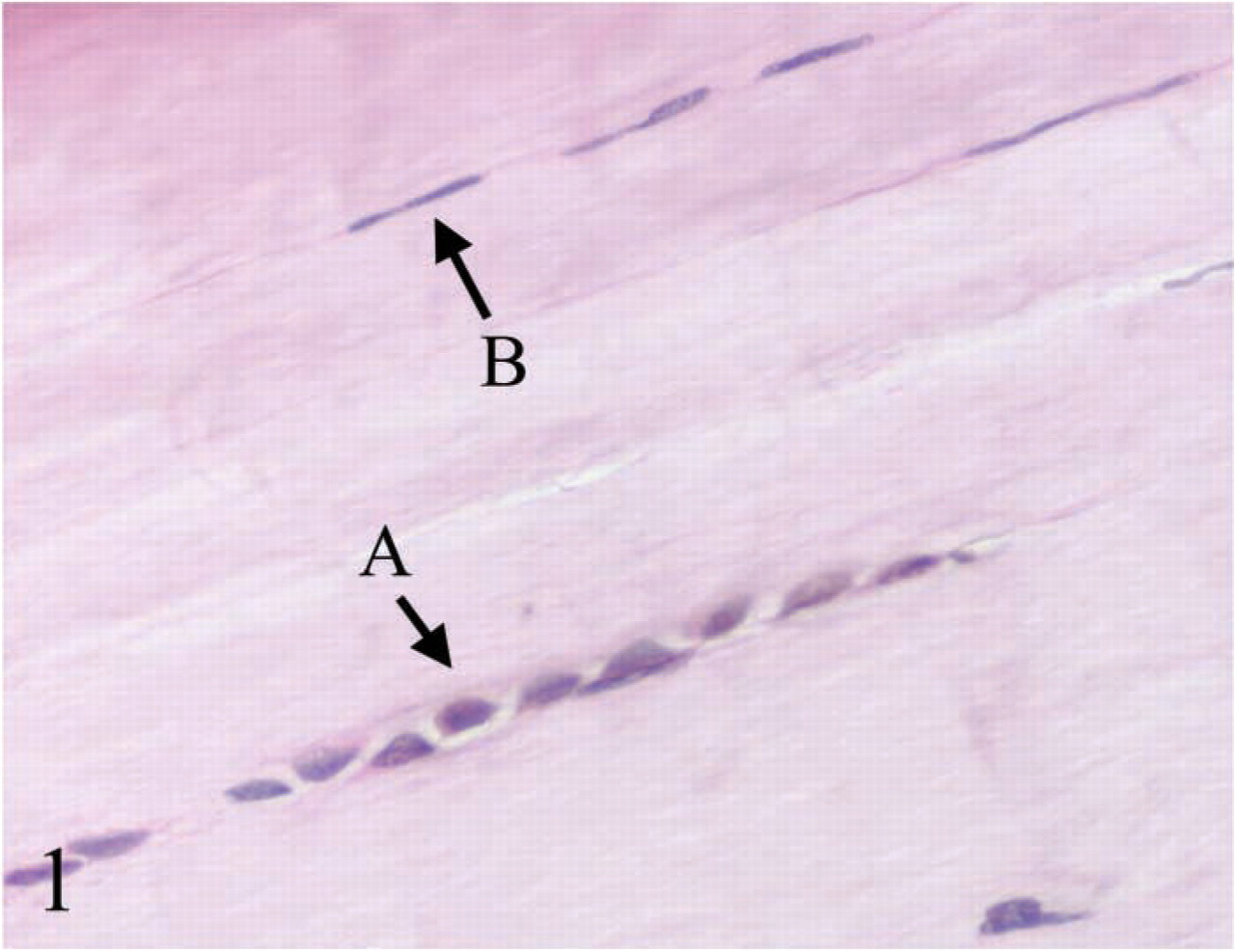
More proliferating cells were observed in (
Image Analysis
After IHC staining, micrographs were taken from each slide using a CCD camera-assisted microscope (Leica Cambridge; Cambridge, UK) and analyzed with Metamorph image analysis software (Universal Imaging; Downingtown, PA). Under ×400 magnification, 20 viewing fields (0.038 μm2) were systematically sampled to avoid allocation bias. Using classifying parameters including length, elliptical form factor, and standard area, tenoblasts and tenocytes can be distinguished on the basis of their morphology. Because cell morphology is the only discriminating feature, borderline cases between tenoblasts and tenocytes were excluded from further analysis, i.e., only “typical tenocytes” and “typical tenoblasts,” defined by a non-overlapping set of classifying parameters, were included. Ovoid tendon cells and elongated tendon cells were distinguished using length and elliptical form factor. Length is defined as the span of the longest chord through the selected object, while the elliptical form factor is defined as the ratio of length to breadth. Breadth is defined as the caliper width that is perpendicular to the length. The reference values for elongated tendon cells are length >30 and elliptical form factor >1.7. The reference values for ovoid tendon cells are length <25 and elliptical form factor <1.5. Positive immunostaining signals were detected by color thresholding on the brown color of DAB peroxidation as described previously (Fu et al. 2002). For proliferation index and apoptosis index, the percentages of proliferating tenoblasts and apoptotic tenoblasts in all tenoblasts were calculated and the same was done for tenocytes. For the evaluation of protein expression in tenocytes and tenoblasts, an adapted scoring scheme was used (no detectable expression = 0; detectable expression with the percentage of positive cells less than 10% = 1; significant expression with the percentage of positive cells much higher than 10% = 2) (Gilardoni et al. 2003), and the results were presented as median scores. Because a paired comparison was employed, detectable expression in score 1 and significant expression in score 2 also referred to a direct comparison in staining intensity, i.e., if an obvious difference in staining intensity between elongated tendon cells and ovoid tendon cells was consistently noted, the higher staining intensity was scored 2 and the lower staining intensity was scored 1, even when the percentages of positive cells were similar.
Statistical Analysis
Statistical analysis was done using the Statistical Package for Social Science (SPSS) 11.0 (SPSS; Chicago, IL). A paired t-test was employed to compare the proliferation index and the apoptosis index after checking for normal distribution by the Kolmogorov-Smirov test. A Wilcoxon sign-ranked test was used to compare the scores of protein expression in tenoblasts and tenocytes in a paired manner for all samples. Significant difference was determined at p<0.05.
Expression levels of proteins in rounded tendon cells and elongated tendon cellsa
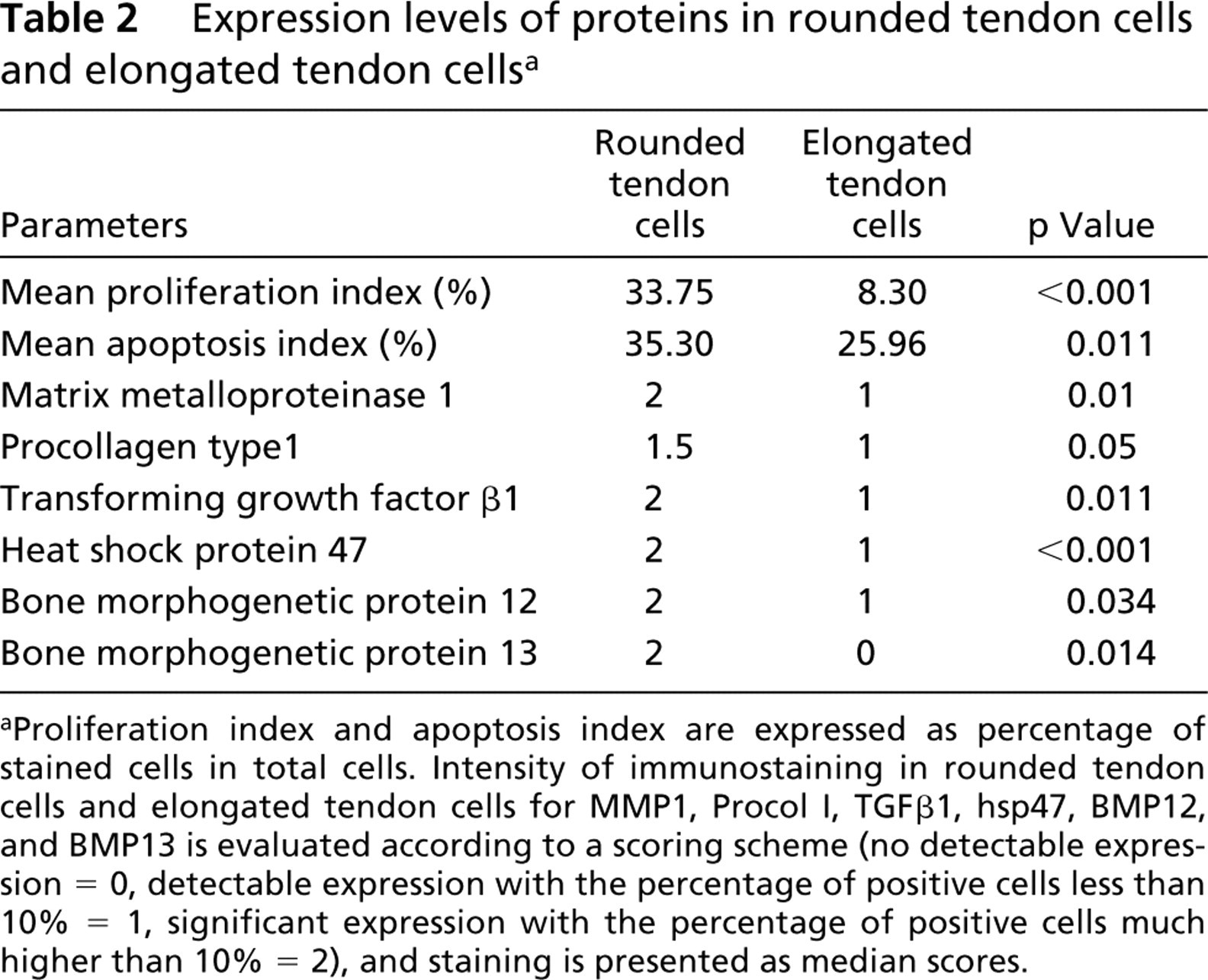
Proliferation index and apoptosis index are expressed as percentage of stained cells in total cells. Intensity of immunostaining in rounded tendon cells and elongated tendon cells for MMP1, Procol I, TGFβ1, hsp47, BMP12, and BMP13 is evaluated according to a scoring scheme (no detectable expression = 0, detectable expression with the percentage of positive cells less than 10% = 1, significant expression with the percentage of positive cells much higher than 10% = 2), and staining is presented as median scores.
Results
Typical elongated tendon cells and round tendon cells are shown in Figure 1. The results of IHC analysis are summarized in Table 2. Both the proliferation index and apoptosis index in ovoid tendon cells were significantly higher than those of elongated tendon cells (p<0.05), as shown by the staining of PCNA (Figure 2) and the results of the TUNEL assay (Figure 3).
All ovoid tendon cells were stained positively with MMP1, procollagen type I, TGFβ1, and BMP13, whereas only a few elongated tendon cells showed detectable expression of these proteins. Significantly higher expression of hsp47 and BMP12 was observed in ovoid tendon cells compared with elongated tendon cells. Representative micrographs for IHC staining of TGFβ1 and BMP13 are shown in Figures 4 and 5, respectively.
Discussion
Although cell heterogeneity in tendons has been recognized for years, the identities of various cell types have not been studied in detail. Here we found that round tendon cells, or tenoblasts, were present in 89% of healthy patellar tendon samples, predominantly in clustered forms. As shown by the proliferation index, tenoblasts are actively proliferating cells (33%), whereas elongated tendon cells, also known as tenocytes, are not (3.64%). It is consistent with the conventional view that tenocytes are terminally differentiated cells with very limited proliferative capacity. The apoptosis index was also higher in tenoblasts, indicating a high cell turnover that would limit the progressive accumulation of tenoblasts. Apoptosis, or programmed cell death, is usually triggered in tissue differentiation, and it has been demonstrated that apoptosis was evident in mesenchymal cells destined to develop into tendons (Sulik et al. 2001). This finding supports the view that tenoblasts are precursor cells for tenocyte differentiation. With respect to the origins, tenoblasts may be remnants from embryonic development (Jozsa and Kannus 1997), generated from activation of tenocytes in response to tendon injury (Kakar et al. 1998) or derived from connective tissue progenitor cells (Muschler and Midura 2002). Although conventional characterization of tendon healing indicates that tenocytes may still retain the capacity for back-differentiation, it is more plausible that the activation of vascular pericytes in endotenons and pre-existing tenoblasts plays a more important role in the generation of tenoblasts (Salingcarnboriboon et al. 2003).
In contrast to tenocytes, which are scattered evenly on the collagen fibers in tendons, tenoblasts often appear in clusters with a localized pericellular region devoid of collagen fiber anchorage. This may explain why the expression of MMP1 in tenoblasts was higher, because MMP1 is one of the major extracellular proteases that can cleave native type I collagen fibers in tendons, and its activity is associated with enhanced cell migration and tendon response to injuries (Stamenkovic 2003). In addition to MMP1, we also found that the expression of hsp47 and procollagen type I was high in these clusters. Because hsp47 is a collagen-specific molecular chaperone that assists in collagen synthesis and production (Nagata 1998), clustered tenoblasts may participate in active collagen matrix remodeling. The presence of localized matrix remodeling processes in normal tendons may be attributed to the cellular response to micro-injuries from daily activities. In fact, these sites may serve as nuclei for the intrinsic healing response as well as the origins of tendon non-healing. It is believed that tenoblasts normally undergo proliferation, matrix remodeling, apoptosis, and finally differentiation into tenocytes to resolve tendon healing. If this process is blocked or if tenoblasts deviate from their normal destination as tenocytes, active tenoblasts would accumulate at the expense of the extracellular matrix. This may result in progressive matrix disturbance and hence a longstanding status of non-healing known as tendinosis. Therefore, studies focused on active remodeling sites and tenoblasts will lead not only to a better understanding of tendon healing and tendon growth but also of pathological conditions such as overuse injuries and tendinosis.
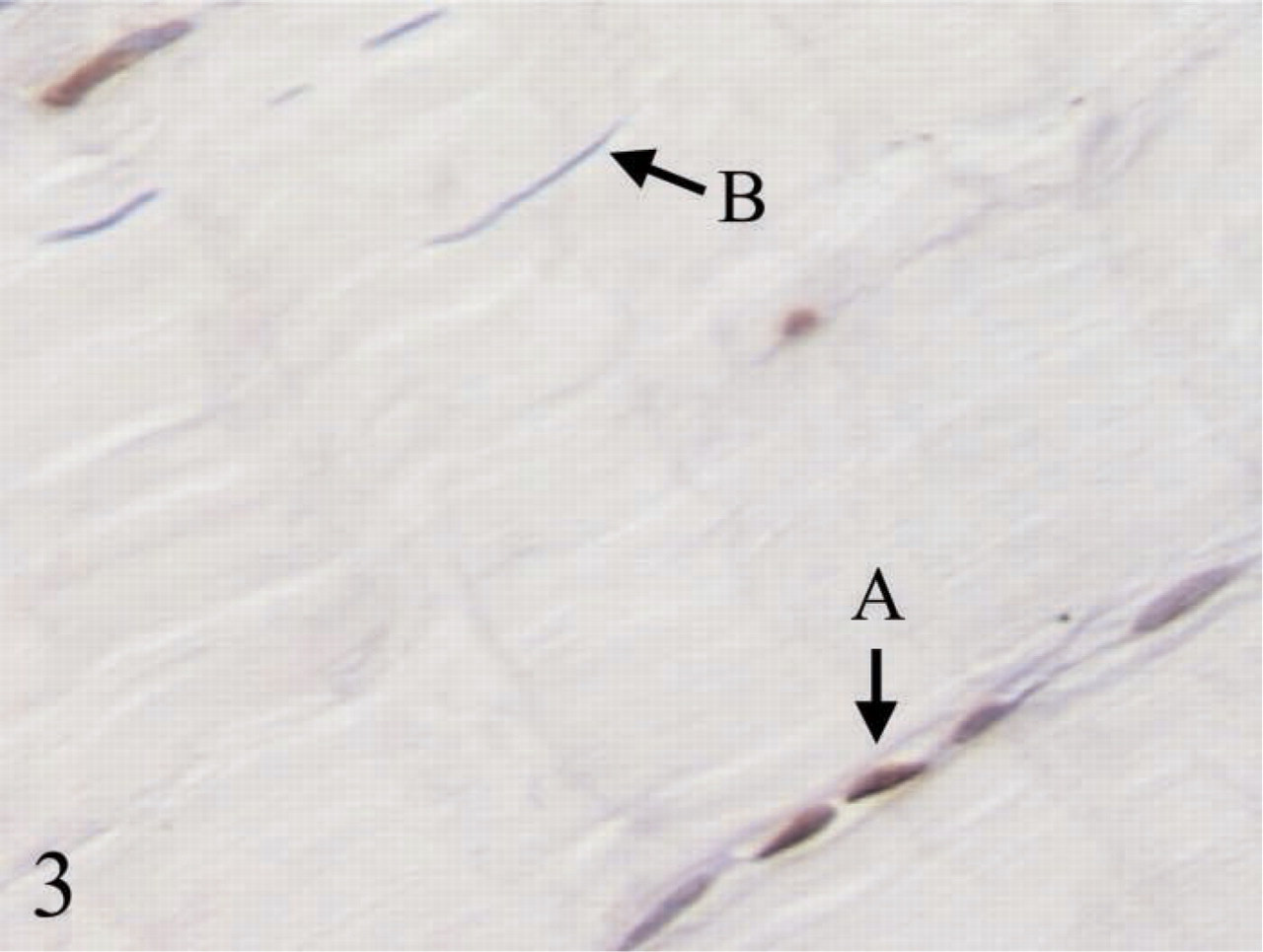
More apoptotic cells were observed in (
Tenoblasts expressed higher levels of TGFβ1, BMP12, and BMP13 compared with tenocytes. With respect to the ability to induce tendon formation (Wolfman et al. 1997) and promote tendon healing (Fu et al. 2003), BMP12 and BMP13 may be involved in the activation of connective tissue progenitors and might trigger the differentiation processes of tenoblasts and tenocytes. It is likely that conversion between tenoblasts and tenocytes is regulated by the expression of these bone morphogenetic proteins. TGFβ1 is one of the key regulatory cytokines for tendon healing (Molloy et al. 2003). It can relay the signals from mechanical loading to modulate the adaptive response in tendons, including collagen synthesis (Heinemeier et al. 2003) and proteoglycan synthesis (Robbins et al. 1997). Therefore, the significant expression of TGFβ1 in tenoblasts suggests that these cells are responsible for the maintenance of matrix integrity in response to injuries and mechanical stimuli. Increased TGFβ1 levels associated with tendon adhesion (Khan et al. 2000) and non-healing tendinosis (Fu et al. 2002) may therefore indicate an accumulation of tenoblasts that leads to augmented collagen and proteoglycan deposition, respectively. Further characterization of tenoblasts, tenocytes, and active remodeling sites may help to explain how tendon respond to the needs of matrix degradation and deposition, such as exercise involving strain and stress, overuse injury, and pathological processes such as tendinosis.

IHC staining of TGFβ1 in healthy human patellar tendon showed that the immunopositivity in (
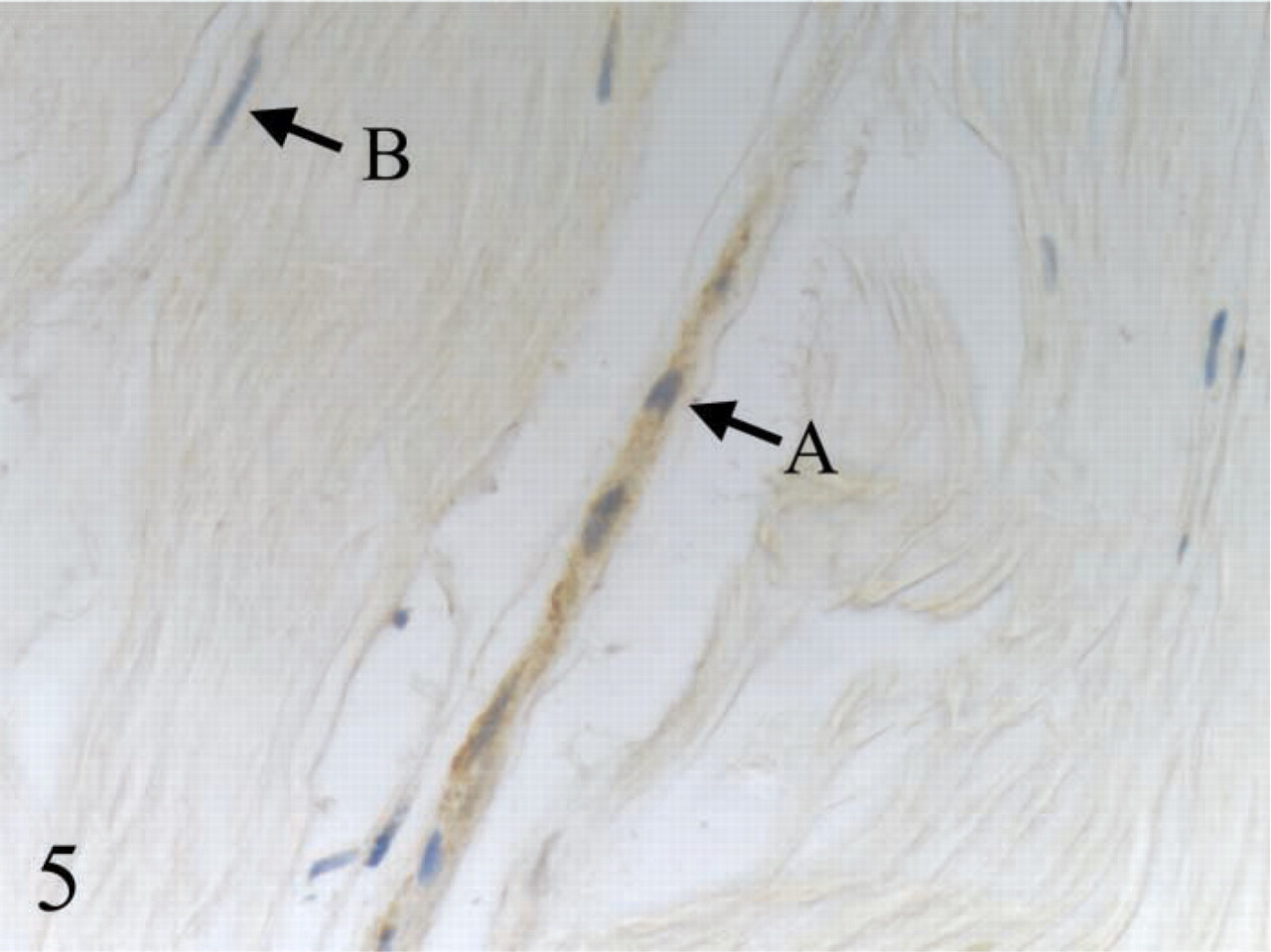
IHC staining of BMP13 in healthy human patellar tendon showed that the immunopositivity (