
Abstract
Early myogenic events in regenerating whole muscle grafts were compared between transgenic MLC/mIGF-1 mice with skeletal muscle-specific overexpression of the Exon-1 Ea isoform of insulin-like growth factor-1 (mIGF-1) and control FVB mice, from day 3 to day 21 after transplantation. Immunocytochemistry with antibodies against desmin showed that skeletal muscle-specific overexpression of IGF-1 did not affect the pattern of myoblast activation or proliferation or the onset and number of myotubes formed in regenerating whole muscle grafts. Hypertrophied myotubes were observed in MLC/mIGF grafts at day 7 after transplantation, although such hypertrophy was transient, and the transgenic and control grafts had a similar appearance at later time points (days 10, 14, and 21). Immunostaining with antibodies to platelet endothelial cell adhesion molecule-1, which identifies endothelial cells, demonstrated no difference in the formation of new vascular network in grafts of transgenic and control mice. Skeletal muscle-specific overexpression of mIGF-1 does not appear to stimulate the early events associated with myogenesis during regeneration of whole muscle grafts.
T
Insulin-like growth factor (IGF-1) is a potent regulator of myogenesis (Florini et al. 1996) and skeletal muscle metabolism (Singleton and Feldman 2001), and there is increasing evidence to support autocrine/paracrine as well as endocrine effects of this growth factor in the regulation of skeletal muscle growth (reviewed in Le Roith et al. 2001; Adams 2002). Increased IGF-1 production is seen in exercising muscle (McKoy et al. 1999) and IGF-1 is undoubtedly implicated in compensatory growth (Singh et al. 1999). In injured muscles, the pattern of elevated IGF-1 mRNA expression in myoblasts closely corresponds to their proliferation (Edwall et al. 1989), indicating a critical role for IGF-1 in tissue repair.
Targeted overexpression of mIGF-1 locally in skeletal muscle by viral vector transfer (Barton-Davis et al. 1998, 1999) or in transgenic mouse models (Coleman et al. 1995; Musaro et al. 2001; Barton et al. 2002) results in muscle hypertrophy. IGF-1 signaling clearly has anabolic effects and plays a central role in muscle hypertrophy (Grounds 2002), and this is associated with stimulation of satellite cell replication (Barton-Davis et al. 1999; Fiorotto et al. 2003). Muscle-specific overexpression of mIGF-1 in transgenic MLC/mIGF-1 mice also has a positive effect on the pheno-type and function of dystrophic muscle (Barton et al. 2002), rescues aging skeletal muscle from atrophy (Musaro et al. 2001), and enhances muscle repair after cardiotoxin injury (Musaro et al. 2001).
To elucidate the mechanisms by which muscle-specific overexpression of IGF-1 enhances muscle regeneration (Musaro et al. 2001), we examined the early events of regeneration (myoblast activation and proliferation, myotube formation and revascularization) in whole muscle autografts in transgenic MLC/mIGF-1 mice. These mice are characterized by sustained muscle-specific overexpression of rat IGF-1 (exon1-Ea isoform) driven by the myosin light chain (MLC)-1/3 regulatory element, which is expressed only in postmitotic muscle cells after differentiation (mainly myo-tubes and myofibers) (Musaro and Rosenthal 1999), with higher expression in fast-twitch (2B, 2× and 2A) myofibers than in slow-twitch (type 1) myofibers (Donoghue et al. 1991). The expression of this transgene in myofibers in vivo clearly leads to hypertrophy (Musaro et al. 2001; Barton et al. 2002).
The whole muscle graft is a precise and reproducible model for studying early morphological events in muscle regeneration (Grounds and McGeachie 1999; Roberts and McGeachie 1992b). Transplantation causes necrosis of most muscle fibers throughout the length of the graft, and any transverse section typically illustrates the pattern of regeneration that occurs progressively from the periphery (marginal edge) of the muscle towards the necrotic central zone. This model allows precise detection of any subtle changes in the timing or pattern of inflammation, revascularization, myoblast activation and proliferation, and onset and extent of myotube formation (White et al. 2000, 2002). The present study tests the hypothesis that local overexpression of mIGF-1 increases the activation and proliferation of myoblasts, the formation of myotubes, and also promotes re-vascularization of the regenerating muscle tissue.
Materials and Methods
Animals
Experiments were performed on male and female 6–8-week-old MLC/mIGF-1 transgenic mice (Musaro et al. 2001). The background FVB strain was used as a wild-type control. The mice colonies were established at the Animal Resource Centre in Perth, Western Australia and were delivered under specific pathogen-free conditions. Mice were housed in standard animal cages and maintained at 12-hr light/dark regimen with free access to standard chow and drinking water. All experiments were conducted in strict accordance with the guidelines of the University of Western Australia Animal Ethics Committee and the National Health and Medical Research Council, Australia.
Genotyping of Transgenic MLC/mIGF-1 Mice
Presence of the MLC/mIGF-1 transgene was screened by polymerase chain reaction using tail digests. The oligonucleotide primers specific for rat IGF-1 used were forward 5′-GAG CTG ACT TTG TAG GCT TCA-3′ and reverse 5′-TTC CTG TCT ACA GTG TCT GTG-3′ (GeneWorks; Sydney, Australia) (fragment size 400 bp).
Surgical Procedures for Whole Muscle Autografts
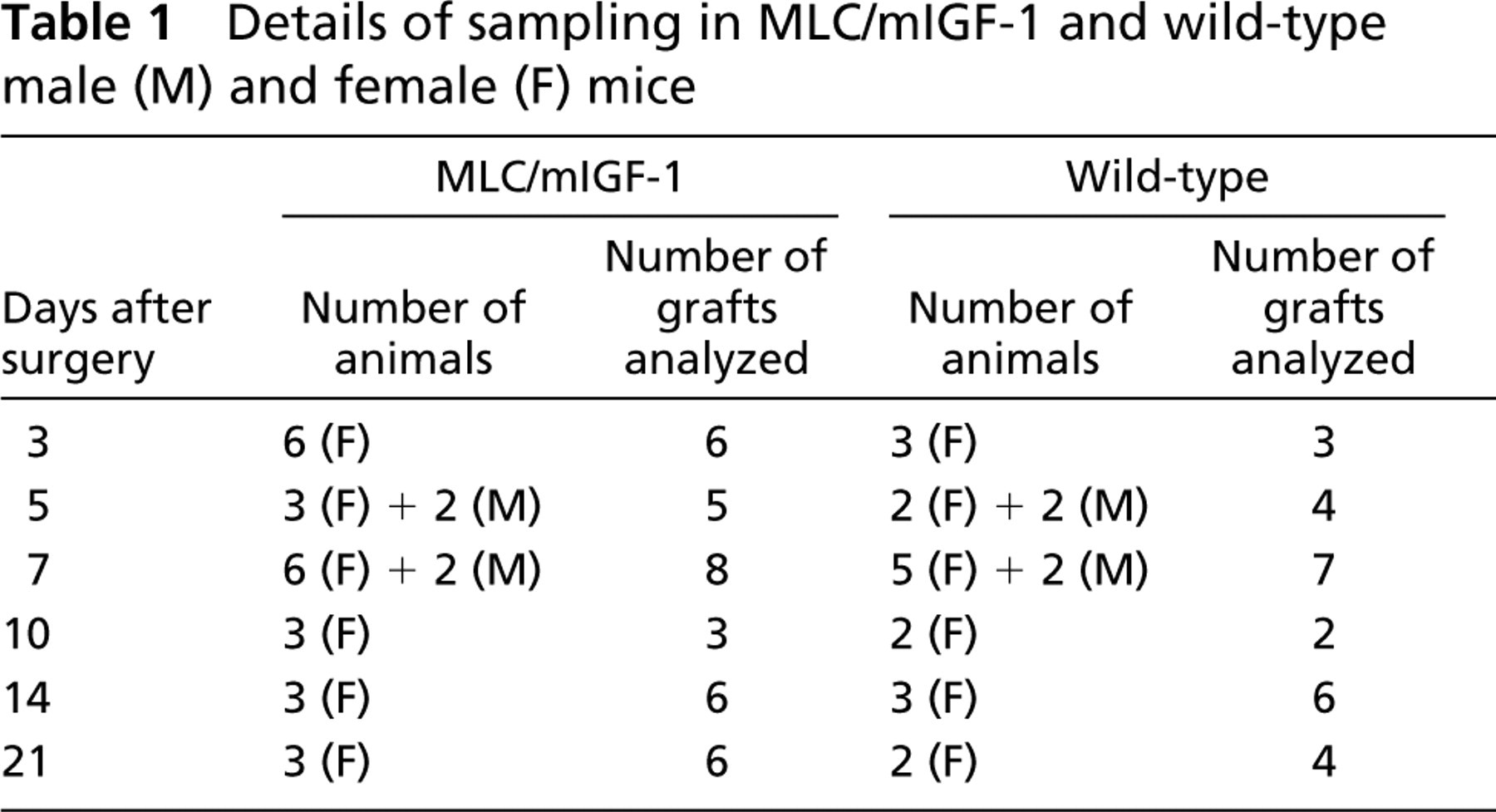
Mice were anesthetized with Halothane and whole muscle autograft transplantation was performed as described previously (Roberts and McGeachie 1992a; White et al. 2000). Briefly the extensor digitorum longus (EDL) muscle was removed from its anatomic bed by cutting both tendons, relocated over the tibialis anterior (TA) muscle of the same leg, stretched to the resting length, and fixed in this position by suturing the tendons. All neurovascular connections to the transplanted EDL were severed during this procedure. Skin was closed and the autografts were allowed to regenerate. Numbers of animals and muscles sampled at each time point for each strain are summarized in Table 1.
Tissue Sampling Procedure
Between 3 and 21 days after grafting, mice were anesthetized and sacrificed by cervical dislocation. Day 3, 7, 10, 14, and 21 grafts were paraffin-embedded or frozen. For paraffin processing, whole EDL muscle grafts with underlying TA muscles were removed, immediately fixed in 4% (w/v) paraformaldehyde (pH 7.6) for 30 min, transferred to 70% ethanol, and processed in a Shandon automatic tissue processor. Muscle grafts were dissected in the mid-region, embedded with both cut surfaces at the top of the paraffin block, and transverse sections (5 μm) collected on silanated glass slides. Several grafts were dissected out with the underlying TA muscle, cut transversely in the mid-region, mounted on cork with tragacanth gum, and frozen in isopentane quenched in liquid nitrogen. Transverse frozen sections (8 μm) were collected on silanated glass slides, air-dried, and stored at −20C. Paraffin and frozen sections were used for hematoxylin and eosin (H and E) staining and immunocytochemistry as described below. One section from each paraffin-embedded and frozen block was stained with H and E for overall morphological assessment of the samples and to confirm the orientation of the grafts.
Details of sampling in MLC/mIGF-1 and wild-type male (M) and female (F) mice
Immunocytochemistry
Polyclonal rabbit anti-desmin (DAKO; Carpinteria, CA) primary antibody was used to identify muscle cells (myoblast and myotubes) in regenerating grafts. Blood vessel endothelial cells were identified using the monoclonal rat antibody to platelet endothelial cell adhesion molecule-1 (PECAM-1) (PharMingen; Sydney, Australia), also known as CD31 (Vecchi et al. 1994). The primary antibodies were detected using biotinylated secondary antibodies donkey anti-rabbit IgG (Jackson ImmunoResearch Laboratories; West Chester, PA) and rabbit anti-rat IgG (Vector Laboratories; Burlin-game, CA), respectively, in conjunction with avidin D–peroxidase (Vector Laboratories) and streptavidin Alexa488 (Molecular Probes; Eugene, OR).
Desmin immunocytochemistry was performed either on paraffin or on frozen sections. Immunofluorescent detection (Alexa 488) on frozen sections was used on day 3 grafts because this was more sensitive than peroxidase color development of desmin antibody (used on paraffin sections) for identifying early activated myoblasts. Paraffin sections were de-waxed, rehydrated, and antigen retrieval was performed before immunostaining for desmin as described previously, with color development using peroxidase substrate DAB (Roche Diagnostics; Sydney, Australia) and counterstaining with hematoxylin (White et al. 2000). All slides were analyzed using a Leica PM RBE microscope.
PECAM-1 immunocytochemistry was performed on frozen sections. After blocking nonspecific antibody binding with 5% (v/v) fetal calf serum, anti-PECAM-1 antibodies were applied at a dilution of 1:70 in 0.5% (w/v) BSA/PBS/0.1 M glycine and incubated at 4C overnight. Sections were sequentially incubated in biotinylated donkey anti-rat secondary antibody and streptavidin–Alexa488. Between incubations, sections were washed three times in 0.5% (w/v) BSA/PBS/0.1 M glycine for 5 min. Slides were mounted with PVA aqueous medium, sealed with nail polish, and stored at 4C until analysis under a fluorescent microscope as described above. Immunochemistry omitting incubation with primary antibodies was used as negative control.
Tissue Analysis
Histological analysis, cell counts, morphometric analysis, and assessment of the percentage vascularity were performed on digital images of the tissues. Slides were placed on a Leica PM RBE microscope connected to a personal computer and a video camera (Hitachi HV-C20M). Non-overlapping images of the entire grafts were taken and tiled together using ImagePro Plus 4.0 (Microsoft) software and an automated microscope stage movement mechanism. Although the pattern of regeneration of the whole muscle grafts is similar throughout its length (unpublished data), all analyses were based on sections from the mid-region of each graft. The pattern of regeneration of the grafts was assessed on paraffin-embedded and frozen transverse sections by calculating the area of the zone occupied by newly formed myotubes (days 5, 7, and 10 grafts) and also by counting total number of myotubes (large desmin-positive cells with well-pronounced cytoplasm and centralized nuclei) in all grafts. Because wide biological variation occurred in the area of the regenerating zone, the myotube counts were also normalized against 0.5-mm2 area of the regenerating zone. Owing to the high biological variation in myotube numbers between individual grafts, the data are presented as histograms showing the total number of myotubes for each individual graft (White et al. 2000). Cross-sectional areas (CSAs) of all myotubes were also measured using ImagePro Plus 4 imaging software by outlining each individual myotube.
The percentage vascularity of the grafts was assessed using a M168 Weibel grid as the total number of points falling on vascular tissue (i.e., within a blood vessel lumen or on a PECAM-1-stained blood vessel wall) were counted. This process was repeated for all fields and the percententage vascularity for each graft was calculated using the equation:
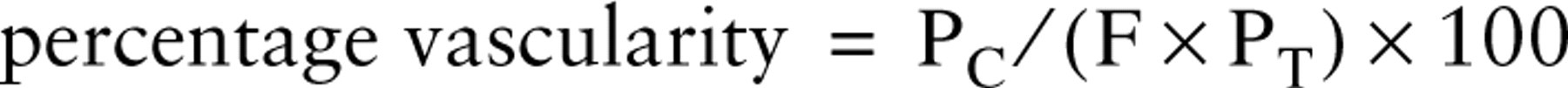
where PC = the total number of points falling on vascular tissue (combined for all field counts in each graft), F = the total number of field counts for each graft, PT = the total number of points on the grid. Data for the percentage vascularity of the grafts are presented for each graft individually.
Results
Whole Muscle Graft Regeneration
Detailed histological examination was carried out on entire transverse sections of 60 grafts at six different times after transplantation. The pattern of whole muscle graft regeneration from day 3 to day 21 after transplantation in transgenic MLC/mIGF-1 and wild-type control FVB mice broadly corresponds to the well-described sequence of histological events (Grounds 1991; Grounds and McGeachie 1999). At day 3 after transplantation, inflammatory cells were not observed in transgenic or wild-type grafts, although desmin-positive cells (myoblasts) were present at the periphery of all grafts (Figures 1A and 1B). This contrasts with other models of muscle damage, such as cardiotoxin (Couteaux et al. 1988) or crush injury (Mitchell et al. 1992), in which the vascular supply is relatively intact and inflammatory cells are abundant among necrotic myofibers by day 1. Semi-quantitative and detailed morphological analysis performed on six transgenic and three control grafts at day 3 did not reveal any differences in the number or distribution of the desmin-positive cells (myoblasts), and there was no evidence of myotube formation at this time in either strain. By day 5, events of degeneration and regeneration were well pronounced and the entire area of all nine grafts was infiltrated with inflammatory cells. The day 5 transgenic and control grafts were very similar, consisting of a necrotic core with disintegrating persisting myofibers and a well-pronounced peripheral regenerating zone. There was no difference in the numbers of desmin-positive myoblasts (recently activated or replicating) or myotubes between the two strains. Newly formed myotubes were present, with larger myotubes clustered at the marginal zone of the graft. By day 7, the regeneration zone was more advanced towards the center of grafts (Figures 1C and 1D). In most cases, the necrotic zone was completely removed, with necrotic myofibers present in only a minority of grafts. The overall morphological appearance of all grafts (both transgenic and wild-type) was similar. By day 10, all grafts were composed entirely of myotubes with very few inflammatory cells. By day 14, all grafts consisted of densely packed clusters of young myofibers, separated by connective tissue, and at day 21 (Figures 1E and 1F), all grafts (in both strains) were composed of small myofibers with centralized nuclei and had a similar appearance. The overall histological analysis of regenerating grafts did not reveal any major differences between transgenic and control animals at any time point.
Myotube Counts in Regenerating Grafts and Pattern of Myotube Formation
Desmin immunostaining on paraffin-embedded and frozen sections was used to visualize myoblasts and newly formed myotubes in regenerating whole muscle grafts. Quantitation of myotubes in grafts at days 5, 7, and 10 after transplantation did not reveal any significant difference in either the onset of myotube formation or the numbers of myotubes between transgenic and control mice, and this was also the case when the data were standardized to take into account variations in the size of the regenerating area of the grafts (Figures 2A and 2B). The only apparent morphological difference between the two strains was hypertrophy of myotubes at day 7 in transgenic MLC/mIGF-1 grafts. Measurement of the CSA of the newly formed myo-tubes in paraffin-embedded day 7 grafts revealed more myotubes with larger CSA in transgenic vs wild-type grafts (Figure 2C). Some hypertrophic myotubes with CSA >800 μm2 were observed in the MLC/mIGF-1 grafts, whereas myotubes of this size were never seen in the wild-type grafts.
Vascular Network of the Whole Muscle Grafts
An antibody against the cell surface marker PECAM-1 was used to visualize the blood vessel network in regenerating grafts (Figure 3), and percentage vascularity was calculated at day 3 and 7 after transplantation. There was no difference between the percentage vascularity of the transgenic vs wild-type grafts at either time point (Figure 4).
Discussion
Early Events in Skeletal Muscle Regeneration Are Unaffected by Targeted Overexpression of IGF-1 in Differentiated Muscle Cells
The pattern of muscle regeneration in whole muscle autografts in young transgenic MLC/mIGF-1 mice was not affected by local overexpression of mIGF-1. These observations do not support any aspect of our initial hypothesis that, in whole muscle grafts, overexpression of mIGF-1 increases muscle regeneration by enhancing myoblast activation and proliferation, the onset of myoblast fusion to form myotubes, number of myotubes formed, and the speed of revascularization of the regenerating tissue. The MLC/mIGF-1 transgene used in the present study was constructed using the rat IGF-1 Ea (Exon 1) isoform driven by the MLC promoter to restrict expression only to differentiated skeletal muscle cells (Barton-Davis et al. 1998; Musaro et al. 2001; Barton et al. 2002). In muscles of these transgenic mice, total levels of IGF-1 protein are almost doubled (16 ng/g vs 9 ng/g) compared with non-transgenic controls (Barton et al. 2002), and it was concluded that mIGF-1 protein remains in the muscle bed and does not enter the circulation (Musaro et al. 2001). Therefore, many of the benefits on regeneration reported after cardiotoxin injury (Musaro et al. 2001) and specifically the early events embodied in the hypothesis would necessitate a paracrine effect by the IGF-1 or else an indirect influence resulting from other factors produced by the transgenic muscle. Although the mIGF-1 (Exon-1 Ea isoform) may not enter the bloodstream, there is evidence that overexpression of this transgene in vivo upregulates other endogenous IGF-1 isoforms that can potentially enter the circulation (Winn et al. 2002), although the biological role and mechanism of action of these isoforms in MLC/mIGF-1 transgenic mice have yet to be elucidated. Furthermore, the possibility of a systemic effect of mIGF-1 overexpression is supported by recruitment of c-kit+ stem cells to the site of cardiotoxin injury (Musaro et al. 2004). Various reasons for the differences between the earlier study using cardiotoxin injury (Musaro et al. 2001) and the present results using the graft model are discussed below.
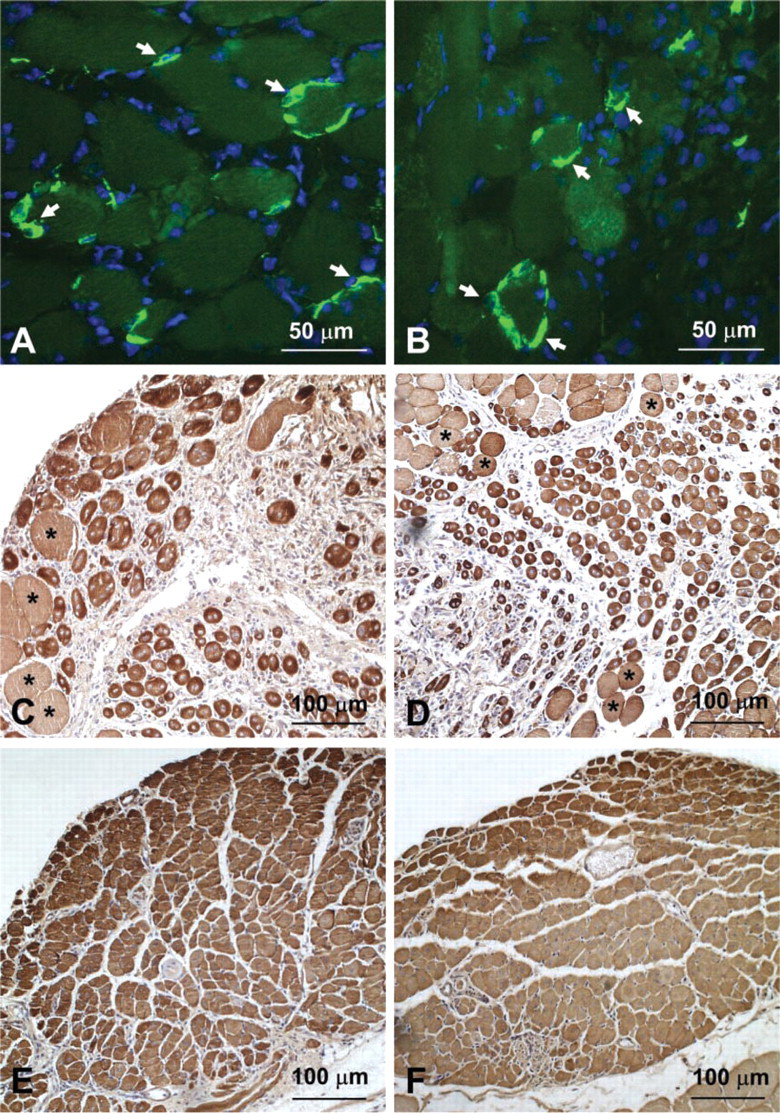
Desmin immunocytochemistry on transverse sections of MLC/mIGF-1 (
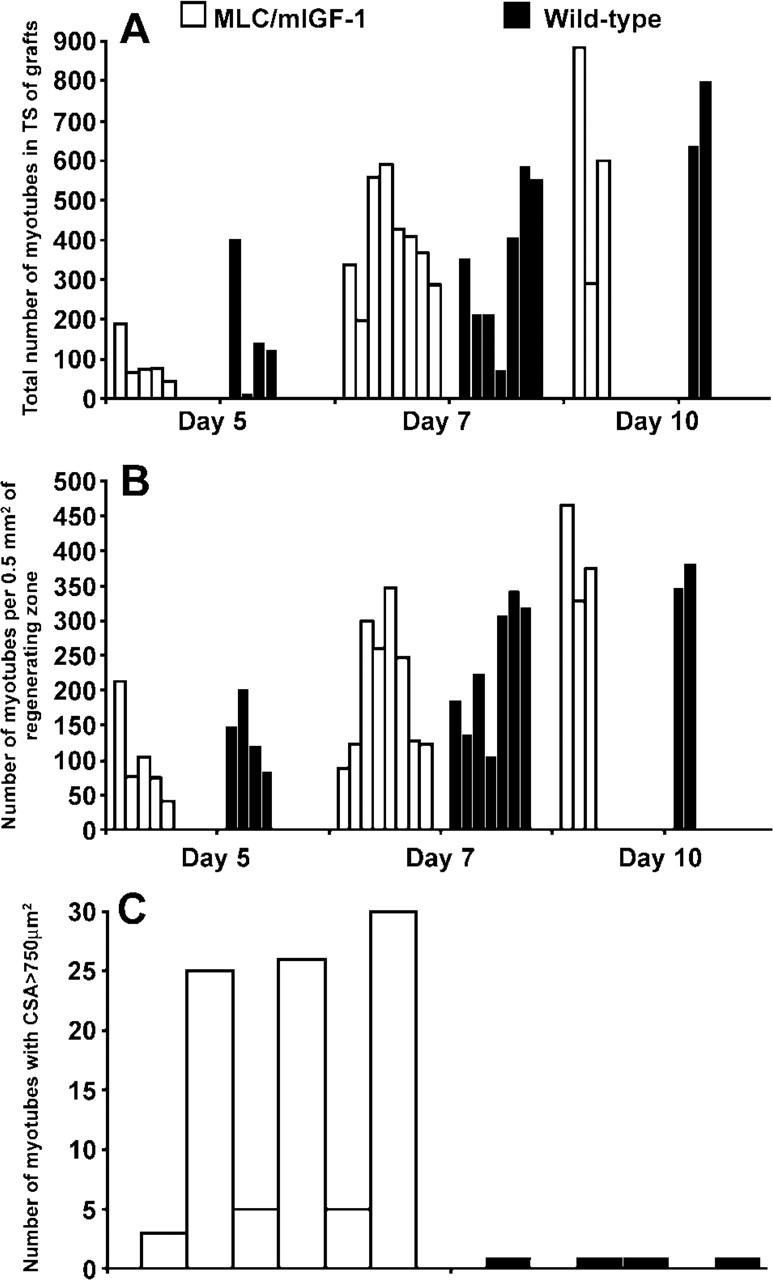
Numbers and sizes of myotubes in the regenerating grafts. (
Differences Between Regeneration in Grafts and Cardiotoxin-injured Muscles
There are fundamental differences between the experimental models of muscle regeneration resulting from whole muscle graft transplantation (used in the present study) and cardiotoxin injection (Couteaux et al. 1988) used by Musaro et al. (2001,2004) and others (Asakura et al. 2002; Yan et al. 2003). Cardiotoxin causes degeneration of the sarcoplasm of most myofibers. However, basal lamina sheaths, vascular supply, and innervation, as well as satellite cells, remain relatively intact (Couteaux et al. 1988). Even when cardiotoxin is injected at high concentration (10 μM), some of the myofibers in the deepest regions of the muscle and near their attachments may escape necrosis (Couteaux et al. 1988). In response to cardiotoxin injury, it is not clear whether regeneration occurs homogeneously through the entire mass of the necrotic tissue or advances progressively from the outside edges of the damaged tissue towards the necrotic central zone as occurs after crush injury (Mitchell et al. 1992). In whole muscle grafts, necrosis occurs throughout the entire length of individual myofibers within the relatively intact basement membrane, although some myofibers persist at the periphery of the graft (Roberts and McGeachie 1992b). In both situations, transgenic mIGF-1 protein presumably disappears from necrotic myofibers and is available only in healthy or surviving muscle cells.
For whole muscle grafts, the intact EDL, which in mice consists of 60% type 2B myofibers (Barton-Davis et al. 1998), is removed and all neuro-vascular connections are severed, in marked contrast to the situation in cardiotoxin-injured muscle. Blood vessel growth into the graft starts around day 3 after transplantation (Roberts and McGeachie 1990, 1992b) in contrast with cardiotoxin injury, where pre-existing blood vessels remain in intimate contact with the damaged muscle (Couteaux et al. 1988). Availability of the vascular supply may be critical in different models. A recent study of cardiotoxin injury (Musaro et al. 2004) in young (3-month-old) MLC/mIGF-1 mice demonstrates that overexpression of mIGF-1 enhances the recruitment of bone marrow-derived c-kit+ stem cells to the damaged site, and these may be converted into myogenic cells. Such recruitment of circulating myogenic stem cells into regenerating whole muscle grafts might be limited by the initial absence of the vascular supply and the barrier of epimysial connective tissue around the graft. Although bone marrow-derived stem cells can certainly contribute muscle nuclei to muscle fibers, this is normally an exceedingly rare event in vivo (Grounds et al. 2002; Morgan and Partridge 2003; White and Grounds 2003). However, enhanced recruitment of such circulating stem cells to muscle formation in vivo in response to mIGF-1 or other factors is highly desirable.
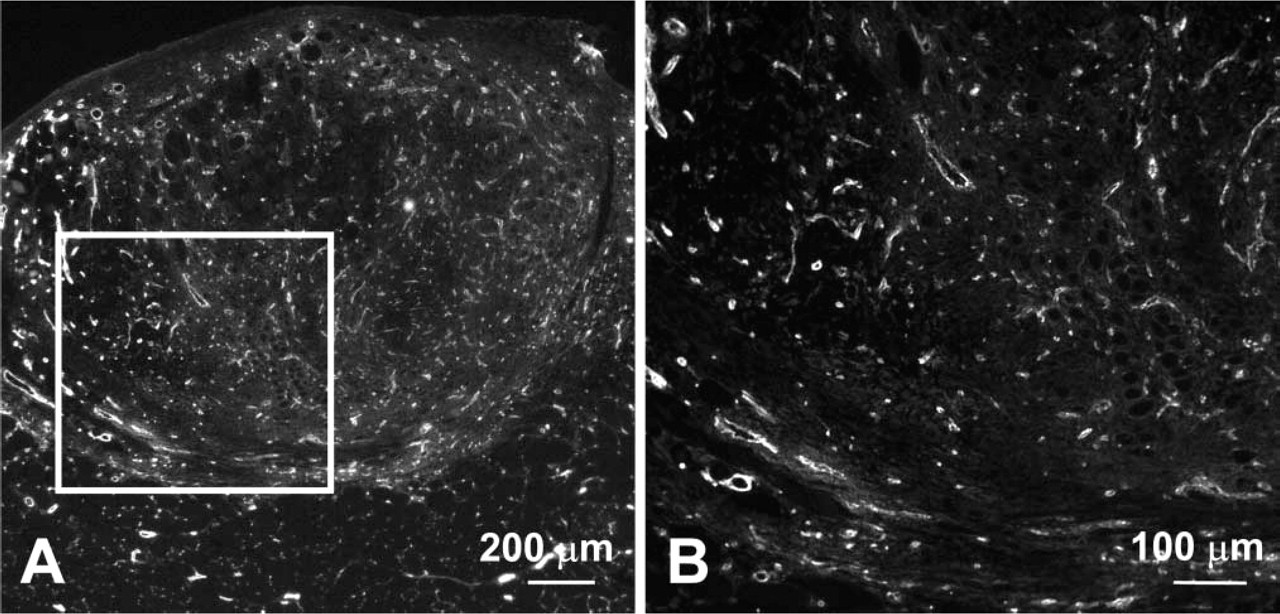
PECAM-1 immunofluorescence on transverse section of a day 7 MLC/mIGF-1 graft. Low (
The early study using cardiotoxin injury in aged (22-month-old) mice (Musaro et al. 2001) concluded that MLC/mIGF-1 transgenic muscle has increased regenerative capacity, based on two sets of observations. The first was the presence of myofibers with central myonuclei that expressed neonatal MyHC isoform in transgenic but not in control muscle. The persistence for many months of central myonuclei is widely used as a marker to identify regenerated myofibers in mature postnatal mouse muscle (McGeachie et al. 1993). However, another possible explanation for central myonuclei within myofibers is denervation, which is known to occur in aging muscles (Larsson 1995). Such internalization of the nuclei has been observed in experimentally denervated muscles of guinea pigs (Tomanek and Lund 1973), rats (Lu et al. 1997), and highly atrophic denervated human muscle fibers (Engel and Banker 1986). As observed in long-term (up to 18 month) experimentally denervated EDL muscles of rats, central nucleation becomes increasingly common with time after denervation and is probably related to decreased CSA of myofibers (Lu et al. 1997). The situation in the cardiotoxin-injured old muscles is further complicated by the fact that upregulation of the embryonic and neonatal MyHC isoforms used as a marker of regenerating muscle (Musaro et al. 2001) has also been demonstrated within pre-existing myofibers after denervation (Schiaffino et al. 1988). Therefore, by either of these criteria it can be difficult to distinguish between denervated (resulting from aging) compared with regenerated myofibers in old muscle.
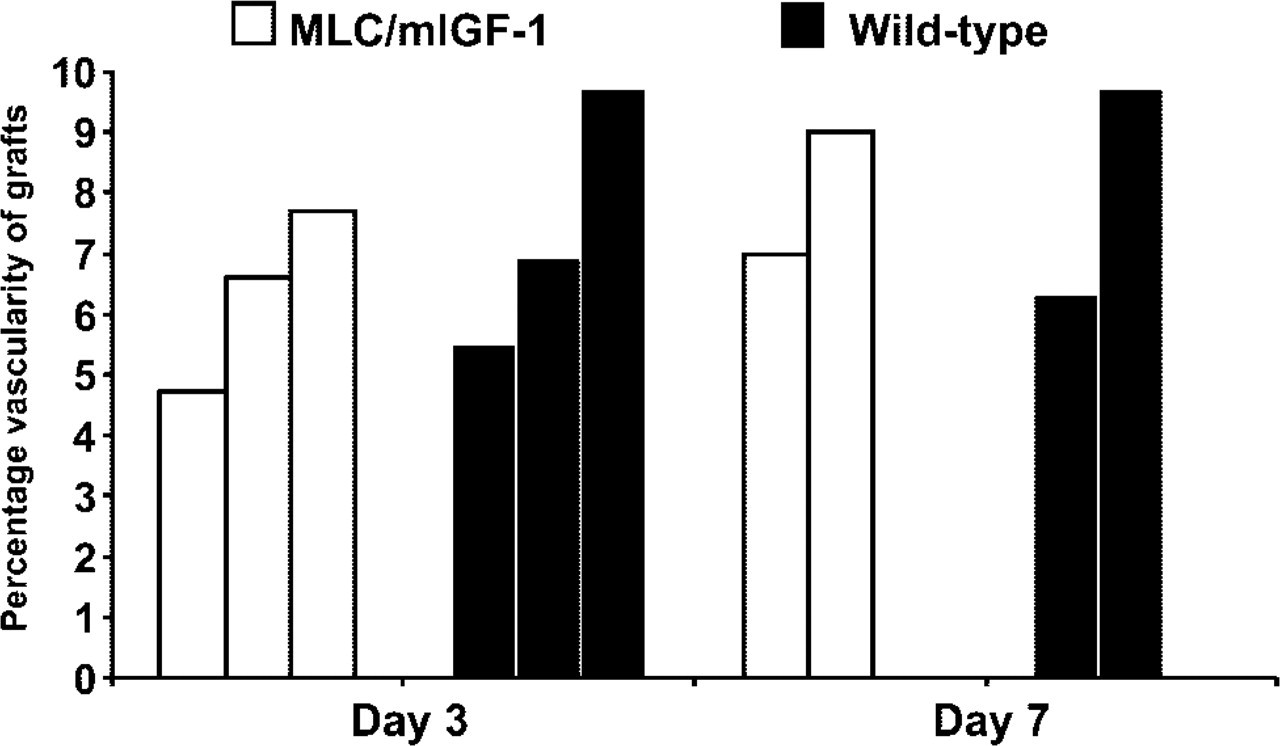
Percentage vascularity of whole muscle grafts. Percentage vascularity was calculated from transverse sections of MLC/mIGF-1 and wild-type grafts immunostained for PECAM-1. Each bar represents an individual graft. There was no significant difference in average percentage vascularity between day 3 (assessed by t-test) or day 7 grafts for transgenic and wild-type mice (total n = 5 per each strain). Similarity between the percentage vascularity of day 3 and day 7 grafts is accounted for by the original (PECAM-1-positive) vasculature persisting in the day 3 grafts.
The study using old (22-month-old) mice also implied that regeneration was normally impaired in old muscle (Musaro et al. 2001). However, although it is often stated that muscle regeneration is impaired or exhausted with aging, this does not appear to apply to early myogenesis and the capacity to form new muscle in vivo. The process of satellite cell activation and new muscle formation seems remarkably robust in normal aging muscle (reviewed in Grounds 1998), although overall functional recovery of regenerated old muscle can certainly be severely compromised due to impaired re-innervation (Carlson et al. 2001) (reviewed in Shavlakadze and Grounds 2003). The same study by Musaro et al. (2001) used intramuscular cardiotoxin injection to injure muscles of 22-month-old MLC/mIGF-1 mice that had been crossed with transgenic desmin/nls-LacZ mice (to readily identify activated myogenic cells), and concluded that muscle regeneration was superior in the transgenic animals compared with controls. Unfortunately, this interpretation is complicated by the fact that there was no apparent injury or regenerative response (as identified by LacZ expression) resulting from cardiotoxin injection into the control muscles.
In summary, the present study found no striking benefit of mIGF-1 overexpression on the early histological events of new muscle formation in whole muscle grafts in young (2-month-old) transgenic mice, in contrast to the enhanced expression of the molecular markers of myogenesis seen after cardiotoxin injury in young (3-month-old) (Musaro et al. 2004) transgenic mice and the original report in old (22-month-old) MLC/mIGF-1 mice (Musaro et al. 2001). This may be accounted for by different features of the transplantation and cardiotoxin injury models of regeneration.
Formation of hypertrophic myotubes in regenerating MLC/mIGF-1 grafts at day 7 in the present study agrees with earlier reports on the ability of this transgene to induce hypertrophy of cultured L6 rat myocytes (that otherwise lack IGF-1) (Musaro et al. 1999) and of myofibers in mature transgenic mice (Musaro et al. 2001) and mdx mice (Barton et al. 2002). However, in regenerating whole muscle grafts such hypertrophy was not apparent at 10 days. This may reflect a normal delay in re-innervation of the grafts, which is required for full maturation of myofibers although not for myotube formation (McGeachie and Grounds 1989). In rats, contact of axons with new muscle fibers in grafts is first detected 18–20 days after transplantation (Hansen-Smith 1983) and myofiber maturation and fiber types are evident by 60 days after grafting (Carlson and Gutmann 1975; Hansen-Smith and Carlson 1979). Rescue of the phenotype of aging skeletal muscle by overexpression of the MLC/mIGF-1 transgene (Musaro et al. 2001) may largely reflect the beneficial effects of IGF-1 on preventing myofiber atrophy (Grounds 2002) and preservation of fast-type myofibers (Barton-Davis et al. 1998), possibly also involving the maintenance of healthy innervation (reviewed in Shavlakadze and Grounds 2003). Although satellite cell proliferation is closely associated with IGF-1-induced hypertrophy (Barton-Davis et al. 1999) and is usually a response to muscle growth, other data obtained from transgenic animals suggest that proliferation can precede hypertrophy (Engert et al. 1996; Fiorotto et al. 2003). The autocrine hypertrophic effects of the MLC/mIGF-1 transgene certainly appear to play an important role in the reduced pathology seen in dystrophic mdx/mIGF-1 mice (Barton et al. 2002), since we have recently shown that mIGF-1 specifically protects dystrophic mdx muscle from the acute onset of muscle breakdown normally seen at 3 weeks of age (unpublished observations).
IGF-1 and Muscle Regeneration
Expression of IGF-1 is normally elevated in regenerating muscles of young (Jennische and Hansson 1987; Jennische et al. 1987; Edwall et al. 1989; Marsh et al. 1997; Hill and Goldspink 2003) rodents, and neutralization of IGF-1 by antibodies in mice decreases the proliferation of myoblasts and delays their fusion into myotubes (Lefaucheur and Sebille 1995). The fact that the process of regeneration is very similar in the MLC/mIGF-1 transgenic and wild-type grafts in the present study might be due to sufficient (optimal) levels of native IGF-1 already being produced by regenerating muscles in this model in normal young mice. In many situations, the availability of IGF-1 or other growth factors may not be the main limiting event in the complex in vivo environment of normal muscle. Instead, the presence of specific receptors and binding molecules may be major players in controlling the mitogenic response of myoblasts (reviewed in Grounds and McGeachie 1999).
With aging, there are reports that the endogenous IGF-1 system is downregulated. For example, the impaired ability to upregulate the endogenous IGF-1 isoform known as mechano growth factor in response to overload has been reported in muscles of old mice (Owino et al. 2001) and humans (Hameed et al. 2003; Harridge 2003). Age-related downregulation of the IGF-1 system is also observed at the IGF-1 receptor level (Martineau et al. 1999). Under such circumstances, sustained overexpression of mIGF-1 may have a particularly marked impact on aging muscle in old transgenic mice (Barton-Davis et al. 1998; Musaro et al. 2001). In muscles of old (24-month-old) rats, upregulated expression of endogenous IGF-1 mRNA has also been reported after bupivacaine injection, similar to the response of young animals. However, high levels of IGF-1 mRNA expression were sustained in old muscles, suggesting some failure in the IGF-1 signal transduction pathway (Marsh et al. 1997). Senescent muscles in rodents show resistance to IGF-1 associated with a dramatic decrease in IGF-1 receptor number (Dardevet et al. 1994; Willis et al. 1997), and a similar situation has been shown for FGF receptors (Johnson and Allen 1995). Although quiescent satellite cells in old muscle may be slower to respond to growth factor stimulation, this appears to have no overall adverse affect on the capacity to form new muscle in response to tissue damage (reviewed in Grounds 1998). The more robust regenerative response in senescent MLC/mIGF-1 mice (Musaro et al. 2001) might be due to the preservation of the healthy muscle phenotype with aging (Barton-Davis et al. 1999; Musaro et al. 2001) by overexpression of the transgenic mIGF-1 directly or through its effects on the endogenous IGF-1 isoforms (Winn et al. 2002) imposed upon a normally age-associated downregulation of the IGF-1 system (Owino et al. 2001; Hameed et al. 2003).
In young growing rats, anabolic hormones (growth hormone, testosterone, and tri-iodothyronine) did not influence protein synthesis at the early stage of regeneration in muscle grafts (Schwartz et al. 1986) and, similarly, there was no difference in early events of regeneration in response to ischemic necrosis, although muscle mass was increased by 4 weeks (Ullman et al. 1989). These observations that early events of muscle regeneration are not altered by exogenous growth hormone might be due to the fact that expression of muscle-specific isoforms of IGF-1 is regulated by local rather then systemic mechanisms, in contrast to liver-specific expression of (circulating) IGF-1 that is induced by growth hormone. Muscle formation can appear quite normal by 1 week even when myotube formation is delayed by several days (White et al. 2000; Smythe et al. 2001), emphasizing the importance of detailed observations throughout the first week of regeneration to detect any alteration to the kinetics of myoblast proliferation or fusion.
IGF-1 and Revascularization
IGF-1 is a potent signal for the growth of many tissues and in regenerating muscles will affect a variety of cell types. Blood vessel growth in particular is responsive to IGF-1 levels (Hellstrom et al. 2001), and activation of the Akt signaling pathway, one of the main pathways downstream from IGF-1 receptor binding, stimulates vascular endothelial growth factor (VEGF) production in cultured C2C12 myotubes and in muscle in vivo (Takahashi et al. 2002). The efficiency of blood vessel formation is a crucial event for the successful regeneration of transplanted grafts (Hansen-Smith and Carlson 1979; Hansen-Smith et al. 1980; Roberts and McGeachie 1990, 1992b), and increased speed of revascularization of transplanted muscle by targeted overexpression of VEGF results in more newly formed myotubes (Smythe et al. 2002). In the present study, we quantitated the percentage vascularity of the regenerating grafts at days 3 and 7 after transplantation using immunocytochemistry with PECAM-1 antibodies. PECAM-1 identifies blood vessel endothelial cells and does not distinguish between newly formed and existing blood vessels. Because the growth of new blood vessels into whole muscle grafts starts around 3 days after transplantation (Roberts and McGeachie 1990, 1992b), immunostaining with PECAM-1 reveals newly formed blood vessels as well as surviving blood vessels (Faulkner et al. 1983) at this time. Therefore, quantitation of the percentage vascularity of the grafts represents total vascularity rather than only neoangiogenesis. There was no difference in the vascularity of the grafts between strains at either of the time points. Therefore, our hypothesis that overexpression of mIGF-1 would affect the pattern of revascularization (in a paracrine manner) is not supported in this muscle autograft model of regeneration.
Footnotes
Acknowledgements
Supported by funding from the Muscular Dystrophy Association of the USA (MG) and by an International Postgraduate Research Scholarship, Australia (TS).
The MLC/mIGF-1 mice were a generous gift from Prof. Nadia Rosenthal, EMBL, Monterotondo, Italy, and we deeply appreciate her constructive criticisms and contribution to this work.
The expert assistance of Esfir Slonimsky (EMBL) in initial mouse breeding and characterization is gratefully acknowledged.