
Abstract
Prion diseases are believed to develop from the conformational change of normal cellular prion protein (PrPc) to a pathogenic isoform (PrPsc). PrPc is present in both the central nervous system and many peripheral tissues, although protein concentration is significantly lower in non-neuronal tissues. PrPc expression is essential for internalization and replication of the infectious agent. Several works have pointed to the gastrointestinal (GI) tract as the principal site of entry of PrPsc, but how passage through the GI mucosa occurs is not yet known. Here we studied PrPc expression using Western blot, RT-PCR, and immunohistochemistry in rat GI tract. PrPc mRNA and protein were detected in corpus, antrum, duodenum, and colon. Immunoreactivity was found in scattered cells of the GI epithelium. With double immunofluorescence, these cells have been identified as neuroendocrine cells. PrPc immunostaining was found in subsets of histamine, somatostatin (Som), ghrelin, gastrin (G), and serotonin (5HT) cells in stomach. In small and large bowel, PrPc cells co-localized with subpopulations of 5HT-, Som-, G-, and peptide YY-immunolabeled cells. Our results provide evidence for a possible and important role of endocrine cells in the internalization of PrPsc from gut lumen.
T
PrPc is a membrane sialoglycoprotein expressed at high concentrations in the CNS (Moudjou et al. 2001) and at lower concentrations in many other peripheral tissues (Fournier 2001). In the case of acquired prion diseases, the infectious agent must first enter and then spread to the CNS. The lymphoreticular system plays an important role in the replication of the scrapie protein (Brown et al. 1999). On the other hand, the pathways of neuroinvasion are less known, although some routes have been proposed (Beekes et al. 1998; Aguzzi 2001). Many authors look to the GI tract as the principal site of entry for the infectious agent (Prinz et al. 2003), but how this passage through the GI mucosa occurs is one of the main questions. PrP is believed to be incorporated from the plasma membrane into endocytic organelles by constitutive transport, although most PrPc recycles back to the membrane without degradation. However, no agreement exists about the intracellular route. Reports on both transport in clathrin-coated vesicles (Shyng et al. 1995) and caveolae-mediated uptake exist (Vey et al. 1996; Peters et al. 2003). Several works have suggested that PrPc is concentrated in caveolae-like domains (CLDs), cell structures involved in crucial physiological processes (Kurzchalia and Parton 1999; Parton 2003). This is why CLDs appear to have an important role in the PrPc-to-PrPsc conversion process.
To understand the process of how and where the infectious agent enters, it is important to know the cell types that express host PrPc along the GI wall. However, few studies have investigated PrPc in the GI tract, and those few have been carried out in mouse (Ford et al. 2002), hamster, and human (Fournier et al. 1998,2000), with a lack of consistency in results. We have studied the rat GI tract because the only PrPc expression analysis in this species has been done with brain, placenta, uterus, and testis (Tanji et al. 1995).
The aims of the present work were to detect, by a combination of cellular and molecular techniques, the presence of PrPc in rat GI tract and to identify PrPc-producing cells.
Materials and Methods
Tissue Samples and Processing
Adult Wistar rats (n = 8) bred at Harlan-INTERFAUNAIBERICA (Barcelona, Spain) were used in this study. Animals were treated according to the guidelines of the Ethical Commitee of the University of Navarra. Rats were anesthetized intraperitoneally with a mixture of ketamine (75 mg/kg), xylacine (10 mg/kg), and atropine (0.05 mg/kg). The abdominal cavity was then opened and samples (stomach, duodenum, and colon) were dissected and immersed in 4% neutral buffered formalin for 24 hr. After fixation, the tissues were paraffin-embedded and 3-μm-thick sections were made. Some samples were immediately frozen in liquid N2 to carry out Western blotting and RT-PCR. In addition, rat brain tissue was used as a positive control in all techniques.
Antisera
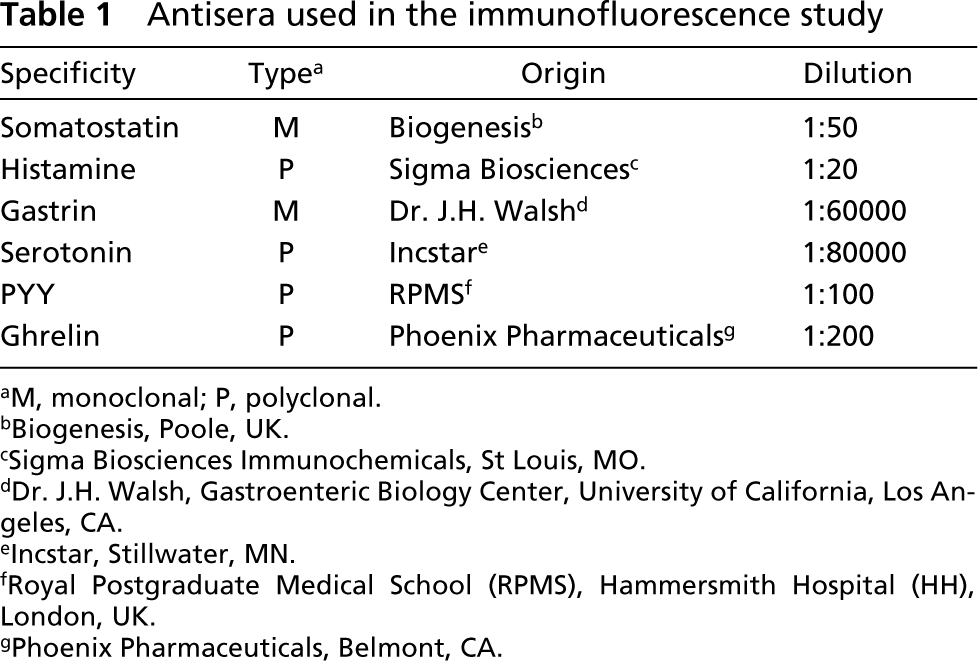
Two antisera against prion protein were used in this study: monoclonal 6H4 (Prionics; Zurich, Switzerland) and polyclonal IgG affinity-purified 91511 (Assay Designs; Ann Arbor, MI) antibodies. In addition, various antisera against some neuroendocrine cell markers (Table 1) were used for further identification of PrP-immunoreactive cells.
Western Blotting
Tissue samples were submerged in lysis buffer (10 mM Tris, pH 7.4, 150 mM NaCl, 1% Triton-X-100, 1% deoxycholate, 0.1% SDS, 5 mM EDTA) with a protease inhibitor cocktail (Roche; Basel, Switzerland) and homogenized with a Polytron PT-K (Kinematica; Lucerne, Switzerland). After centrifugation the supernatant was removed and distributed into different tubes. The final protein concentration was determined with the BCA kit (Pierce; Rockford, IL). Protein extracts were electrophoretically separated on a 12% SDS-PAGE gel and run at 130 V for 90 min under reducing (5% β-mercaptoethanol) conditions. One hundred μg of protein from the different organs was loaded into the sample wells. For brain extracts used as positive controls, 25 μg of protein was loaded. Proteins were transferred to 0.2 μm pore diameter nitrocellulose membranes (Bio-Rad; Hercules, CA) at 400 mA for 1 hr. The membranes were blocked for 1 hr in 10% PBS-milk and incubated overnight at 4C with the primary 6H4 anti-PrP antibody at a 1:6000 dilution. Membranes were washed four times in 0.1% Tween-20 in PBS and incubated with horseradish peroxidase-conjugated anti-mouse antibody at a 1:6000 dilution for 1 hr at room temperature. After four washes in 0.1% Tween-20 in PBS, immune complexes were detected with Lumi-LightPLUS Western Blotting Substrate (Roche) according to the supplier's instructions.
Antisera used in the immunofluorescence study
aM, monoclonal; P, polyclonal.
bBiogenesis, Poole, UK.
cSigma Biosciences Immunochemicals, St Louis, MO.
dDr. J.H. Walsh, Gastroenteric Biology Center, University of California, Los Angeles, CA.
eIncstar, Stillwater, MN.
fRoyal Postgraduate Medical School (RPMS), Hammersmith Hospital (HH), London, UK.
gPhoenix Pharmaceuticals, Belmont, CA.
RNA Extraction and RT-PCR
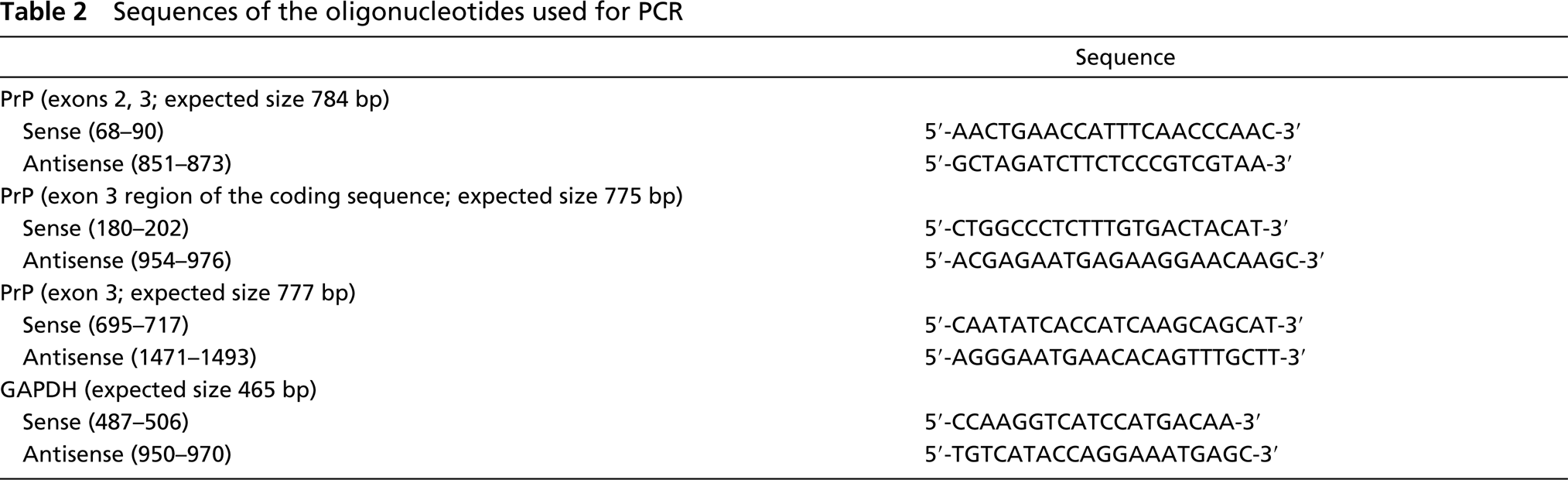
Total RNA was obtained with the Ultraspec RNA Kit (Biotecx; Houston, TX), according to the manufacturer's instructions. The RNA concentration was spectrophotometrically determined. RNA was retrotranscribed with SuperScript II RNase-free Reverse Transcriptase (Invitrogen; Carlsbad, CA). PrP primers (Table 2) were designed based on the published rat sequence [GenBank accession numbers D50092 (exons 1, 2) and D50093 (exon 3)] and synthesized by Invitrogen. PCR was first performed to rule out genomic DNA contamination, employing a pair of primers including exons 2 and 3 of the PrP gene. Then PCR was carried out with two sets of primers that amplified the exon 3 region that encodes for PrP protein. PCR was performed in a total volume of 50 μl by using 2 μl of cDNA, 5 μl of 10 × buffer, 1.5 μl of MgCl2 (50 mM), 1 μl of dNTPs (10 mM), 1 μl of each primer (10 μM), 38 μl of ddH2O, 0.5 μl of BioTaq DNA polymerase (5 U/μl; Bioline, Canton, MA). After incubation at 94C for 10 min in a P-E Thermocycler-2400 (Perkin-Elmer; Foster City, CA), 30 cycles were performed for 30 sec at 94C, 30 sec at 59C, and 1 min at 72C, followed by 10 min at 72C. Water blank and non-RT samples were used as negative controls. To assess RNA integrity, rat GAPDH was amplified on the same samples. The primers used for GAPDH are shown in Table 2. PCR was performed in a total volume of 50 μl by using 1 μl of cDNA, 5 μl of 10 × buffer, 2.5 μl of MgCl2 (50 mM), 2.5 μl of dNTPs (10 mM), 2 μl of each primer (20 ng/μl), 34.5 μl of ddH2O, 0.5 μl of BioTaq DNA polymerase (5 U/μl; Bioline). After incubation at 95C for 5 min, 30 cycles were performed for 1 min at 95C, 1 min at 54C, and 1 min at 72C, followed by 10 min at 72C. Images were processed with Scion Image software (Scion; Frederick, MD) and captured in TIFF format.
Immunocytochemistry
Paraffin sections were mounted on slides coated with Vecta-bond (Vector Laboratories; Burlingame, CA). Tissue sections were deparaffinized, rehydrated to water, and endogenous peroxidase was blocked with 3% H2O2 for 10 min. Slides were washed with distilled water for 5 min, placed in citrate buffer 0.01 M (pH 6), and heated in a microwave (Balay W-2112, 1150–700 W; Madrid, Spain) for 10 min at maximal power and 10 min at minimal power. Background blocking was performed with normal goat serum (DAKO; Glostrup, Denmark) before incubation with specific antiserum. Tissue sections were incubated overnight at 4C with 6H4 monoclonal or 91511 polyclonal antiserum diluted 1:800 for stomach, 1:400 for duodenum, and 1:200 for colon. After rinsing in Tris-HCl 0.05 M buffer, 0.5 M saline, pH 7.6 (TBS), single immunohistochemical staining for prion protein was performed using the EnVision System (DAKO) as previously described (Sanchez and Burrell 2002), the only difference being that the peroxidase activity was revealed according to the manufacturer's recommendations. All experiments included rat brain sections as positive controls. Adsorption controls were performed to test the specificity of immunostaining. Antisera to PrPc were preincubated for 12 hr at 4C with their respective synthetic peptides at a concentration of 0.1 nmol/ml of optimally diluted primary anti-serum before application to tissue sections.
Sequences of the oligonucleotides used for PCR
Double Immunofluorescence
Tissue sections were deparaffinized, rehydrated, and microwave-preheated as described above. Slides were incubated with 2% bovine serum albumin (BSA; Sigma, St Louis, MO) in TBS for 30 min and incubated overnight at 4C with a mixture of the specific antisera (monoclonal and polyclonal) at their optimal dilutions. The slides were washed in TBS and incubated with a mixture of Alexa Fluor 568 TRITC–goat anti-mouse and Alexa Fluor 488 FITC–goat anti-rabbit antisera (Molecular Probes; Eugene, OR) for 45 min. Secondary antibody dilutions were 1:200 for PrPc and 1:400 for the rest of the primary antisera. Then slides were washed six times in 0.1% Tween-20 in TBS. Finally, slides were mounted in PBS–glycerol. Each secondary reagent was confirmed to be species-specific by secondary antibody crossover (mouse or rabbit primary antibody followed by anti-rabbit or anti-mouse secondary, respectively). Sections were examined in a Nikon Eclipse E800M microscope equipped with filters giving excitation at a wavelength of 510–560 nm for TRITC and 465–495 nm for FITC, and images were captured in TIFF format with Analysis Soft Imaging System Gmbh software.
Results
Western Blot
Western blot analysis revealed an immunoreactive band corresponding to PrPc at the expected molecular weight of ~33 kD in all regions of the GI tract (Figure 1). The level of PrPc detected in the rat GI tract was considerably lower than in brain, considering that four times more protein was loaded. Differences in the intensity of expression of the protein between regions were also found, being higher in the stomach than in the duodenum and colon. Both antibodies 6H4 and 91511 gave the same results, although band intensity was greater with the former.
Immunocytochemistry
In rat tissues, labeling for PrPc was found throughout the GI tract (Figure 2). Immunoreactivity was found in scattered cells of the GI mucosa. Both antibodies 6H4 and 91511 gave similar immunocytochemical results, although immunostaining was more intense when the latter was applied. In the stomach, PrPc-immunoreactive cells were scattered in both the fundic (Figure 2A) and pyloric (Figures 2B and 2C) glands and were preferentially located in the lower region of the glands. Immunoreactive cells showed round or oval morphology and sometimes presented slender cytoplasmic processes reaching the apical surface or extending towards neighboring cells (Figure 2C, arrows). Immunoreactivity was preferentially localized in the basal region of the cell, around the nucleus, but was also present within the cytoplasmic processes. PrPc-immunoreactive cells were more numerous in the pyloric (Figure 2B) than in the fundic (Figure 2A) mucosa. In the duodenum, immunostained cells appeared scattered in the epithelium of the villi (Figure 2D) and the crypts, and were less numerous than in the stomach. Some of these cells, mainly those of the villi, had the typical morphology of intestinal open endocrine cells with long apical processes reaching the lumen (Figure 2E). The labeling for PrPc extended throughout the cytoplasm reaching the luminal processes. The lowest amount of PrPc-positive cells was found in the colon. Cells appeared to be dispersed in the epithelium of the crypts (Figure 2F), and immunoreactivity was mainly located in the basal region of the cells (Figure 2G). In all GI regions, adsorption controls confirmed the specificity of the immunostaining (Figure 3).
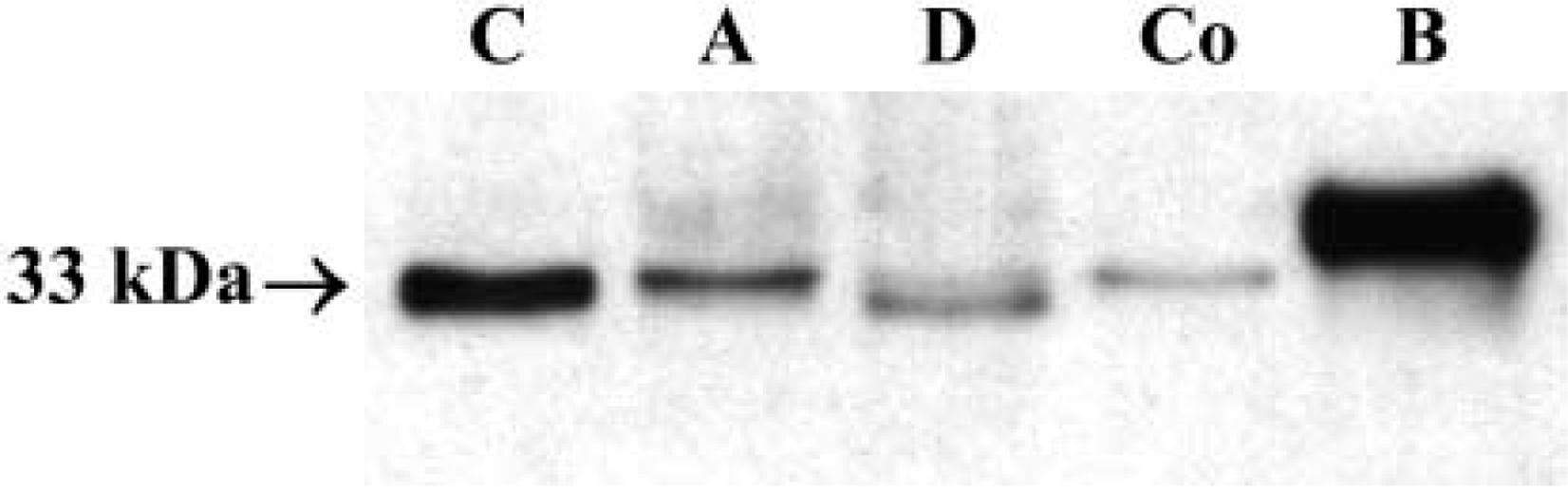
Western blot of rat GI tract. An immunoreactive band of ~33 kD (the expected molecular weight) was obtained in all the regions of the GI tract. The level of PrPc detected in GI samples was significantly lower than in brain, considering that four times more protein was loaded. C, corpus; A, antrum; D, duodenum; Co, colon; B, brain.
Double Immunofluorescence
The first identification of PrPc-labeled cells as neuroendocrine cells was made on the basis of their morphology. Then, to determine the particular endocrine cell type displaying PrPc immunoreactivity, a double-immunofluorescence technique was performed (Figures 4 and 5). In the corpus (Figures 4A–4F), a few histamine-immunoreactive cells (Figure 4A) were also immunolabeled for PrPc (Figure 4B). In the same way, PrPc (Figure 4C) was detected in subpopulations of ghrelin-positive (Figure 4D) and Som-positive cells (Figures 4E and 4F). In the antrum (Figures 4G–4L), most G-cells (Figure 4G) were immunoreactive for PrPc (Figure 4H), but occasional G-cells (Figure 4I) did not stain for PrPc (Figure 4J). In addition, a few PrPc-immunostained cells were identified as 5HT-(Figure 4K) and Som- (Figure 4L) producing cells. In duodenum (Figures 5A–5H), some PrPc-immunolabeled cells (Figures 5A and 5C) displayed immunoreactivity for 5HT (Figures 5B and 5D), while others (Figure 5E) were detected as Som cells (Figure 5F). However, not all the 5HT and Som cells were also stained for PrPc. In addition, co-localization between some PrPc-labeled cells (Figure 5G) and a subpopulation of G-cells (Figure 5H) was also found. Finally, in colon (Figures 5I–5P) partial co-localization between 5HT (Figures 5I and 5K) and PrPc (Figures 5J and 5L) was detected. Some PrPc-immunoreactive cells (Figure 5M) showed immunoreactivity for PYY (Figure 5N), but not all PYY cells stained for PrPc. As in duodenum, only a subset of Som cells were also positive for PrPc (Figures 5O and 5P).
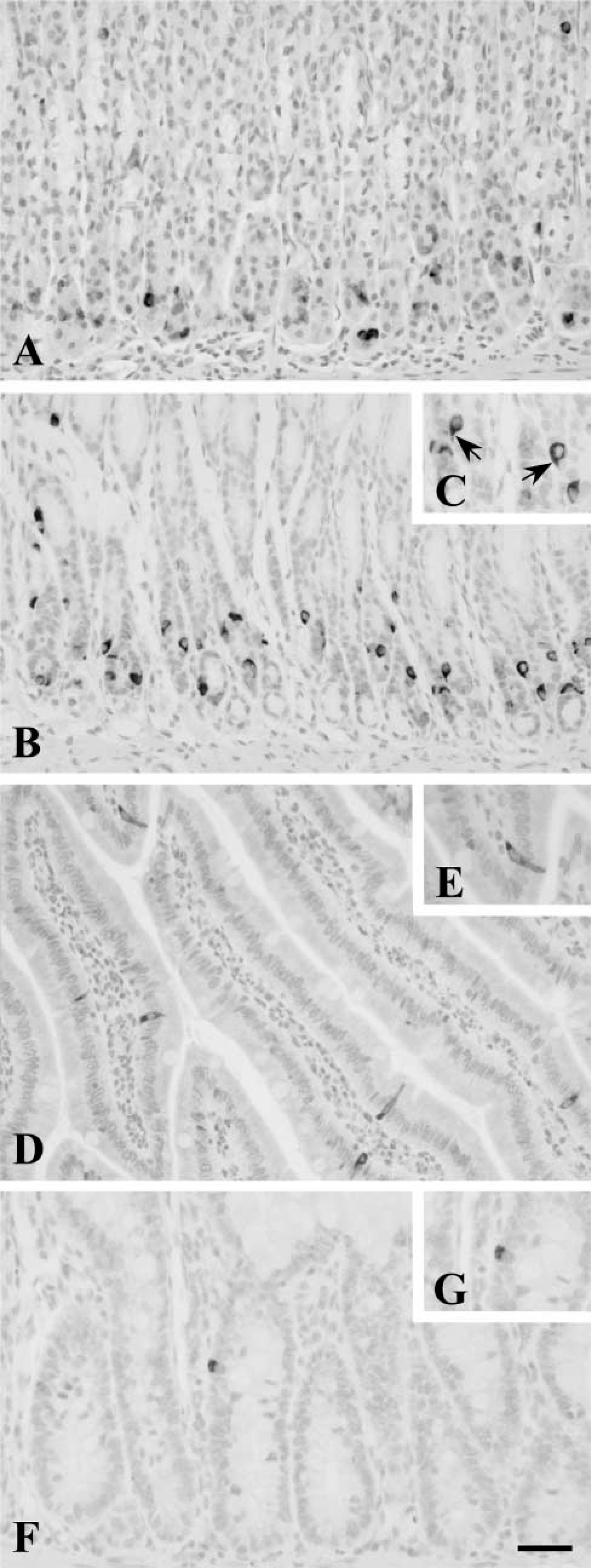
PrPc immunoreactivity in the rat GI tract. PrPc-immunoreactive cells appear scattered in the basal region of fundic (
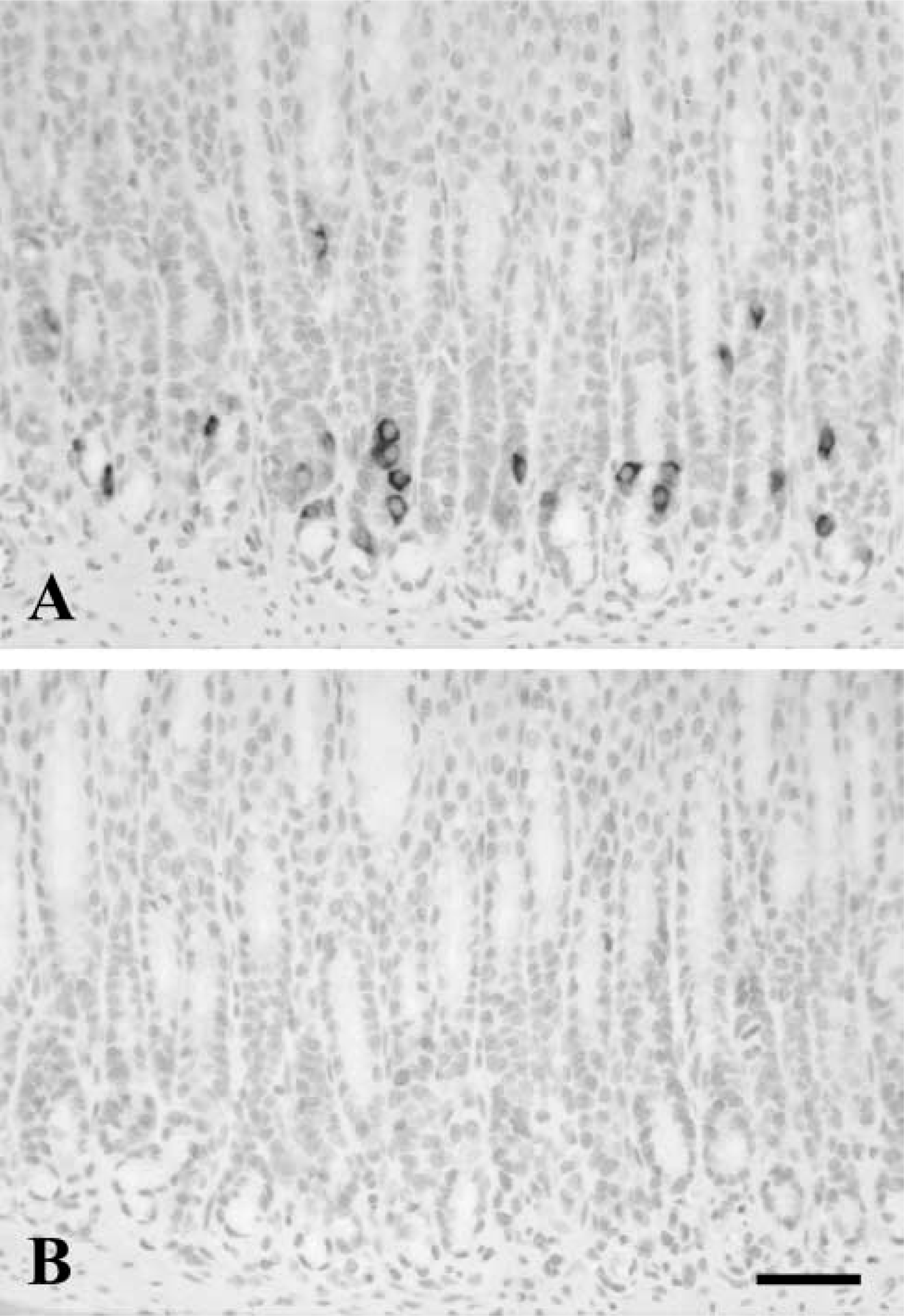
Adsorption control in rat pyloric mucosa. Immunostaining for PrPc in neuroendocrine cells (
RT-PCR
Using the three different sets of primers (Table 2) to detect PrPc mRNA, the expected 784-, 775-, and 777-bp PCR products were found (Figure 6A) in control tissue (brain) and in the different regions of the rat GI tract (stomach, duodenum, and colon). Control transcript GAPDH from all samples was obtained (Figure 6B).
Discussion
Here we demonstrate that PrPc is expressed in rat GI tract, as determined by several analytical techniques. PrPc expression has previously been found in mammalian GI tract, but this is the first report in rat GI tract. We have found that PrPc immunoreactivity is located in endocrine cells and that immunoreactive cell number decreases from the stomach to the small and large bowels.
Immunoreactivity for PrPc in epithelial cells of the GI tract had already been described in human (Fournier et al. 1998,2000; Pammer et al. 2000), mouse (Ford et al. 2002), and hamster (Fournier et al. 1998,2000). However, there is no agreement on which cell type expresses PrPc. In rat (present work), human (Pammer et al. 2000), and mouse (Ford et al. 2002) GI tract, PrPc immunoreactivity has been detected in endocrine cells. On the contrary, other authors have described PrPc immunolabeling in mucous and parietal cells in humans (Fournier et al. 1998,2000; Pammer et al. 2000) and in mucous, parietal, and goblet cells in hamsters (Fournier et al. 1998,2000). Recently, PrPc has been localized in cell–cell junction domains of human enterocytes (Morel et al. 2004). These apparent discrepancies may be due to differences in the antisera, the fixation process, or the pretreatments used, or to changes in the pattern of expression of the protein among species. In fact, different subcellular locations for PrPc in the same cell type have been reported with the use of different antibodies (Fournier et al. 2000). In transgenic mice expressing the green fluorescent protein reporter gene under the control of bovine PrP promoter, correlation between fluorescence and PrPc immunohistochemistry has been found but no fluorescence has been observed in gut epithelial cells (Lemaire-Vieille et al. 2000). In the same way, other authors did not find PrPc in gut epithelium of neonatal mice (Okamoto et al. 2003). These findings suggest that efficiency of the antisera and tissue processing are critical factors in detection of PrPc.
Our immunocytochemical findings are in agreement with the Western blot results because the more intense bands were found in GI regions (antrum and corpus) in which a high number of PrPc-positive cells were detected. In addition, demonstration of PrPc mRNA by RT-PCR further supports the expression of prion protein in the rat GI tract.
Few works have demonstrated the nature of PrPc positive cells as endocrine cells using phenotypic markers. The first report related to PrPc immunoreactivity in endocrine cells was in human antrum (Pammer et al. 2000). Using consecutive sections, PrPc-positive cells were identified as G-cells, although no images demonstrating this fact were shown. Similarly, in rat antrum we have also found co-localization between G and PrPc but a few 5HT- and Som-positive cells also showed PrPc labeling. Furthermore, human fundic glands were reported to be negative (Pammer et al. 2000). On the contrary, in rat corpus we have found a high number of PrPc-positive cells in the basal region of the glands that co-localized with histamine, ghrelin, or Som. For small and large intestine, our results agree with those of Ford et al. (2002) in relation to the co-localization between 5HT and PrPc. Nevertheless, the present study demonstrated that only a subset of 5HT cells were PrPc-immunoreactive and that staining for PrPc was also present in subpopulations of G, Som, and PYY cells. The presence of PrPc in subpopulations of particular neuroendocrine cell types suggests a tightly regulated expression of this protein.
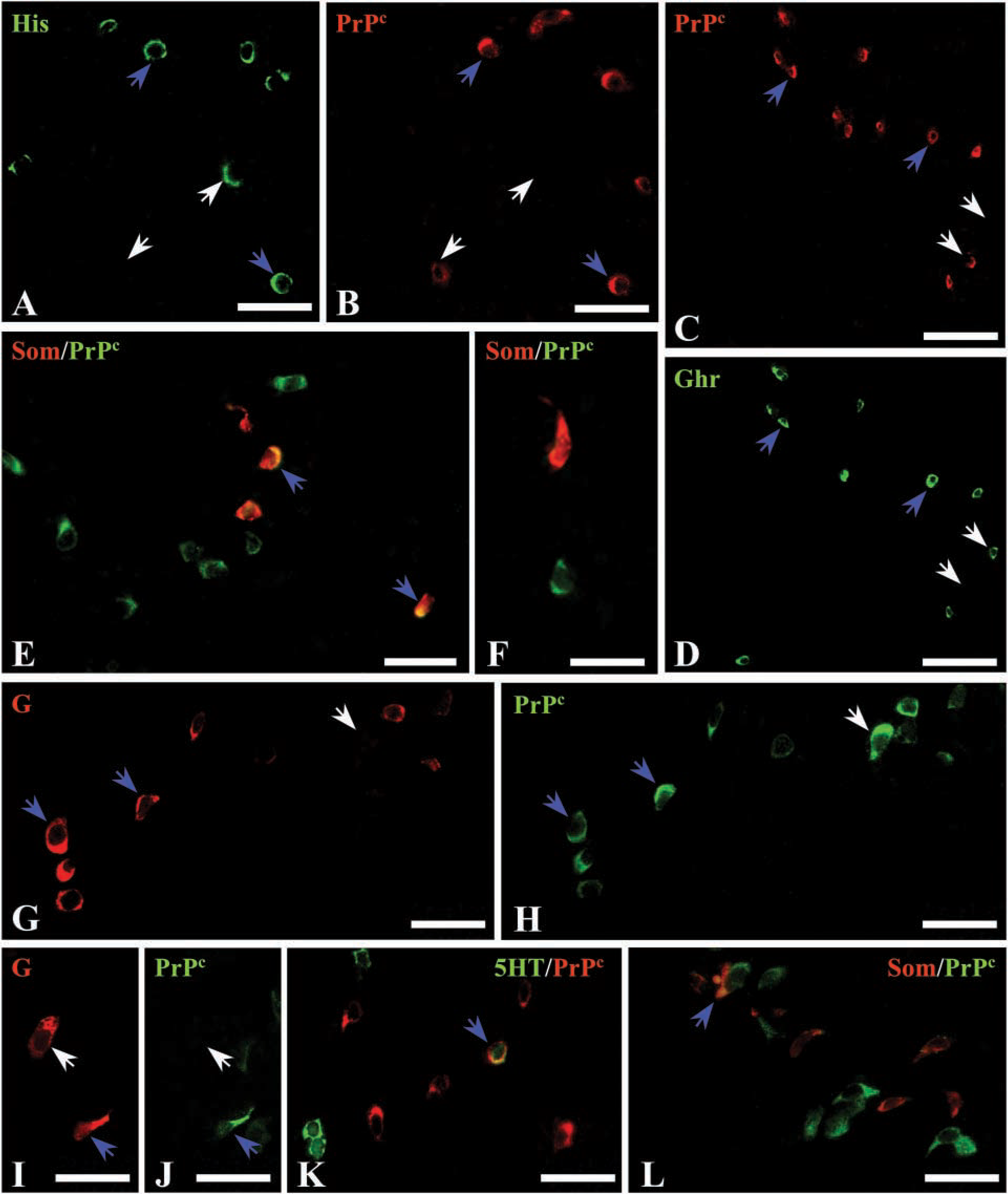
Double immunofluorescence for PrPc and neuroendocrine markers in rat stomach. (
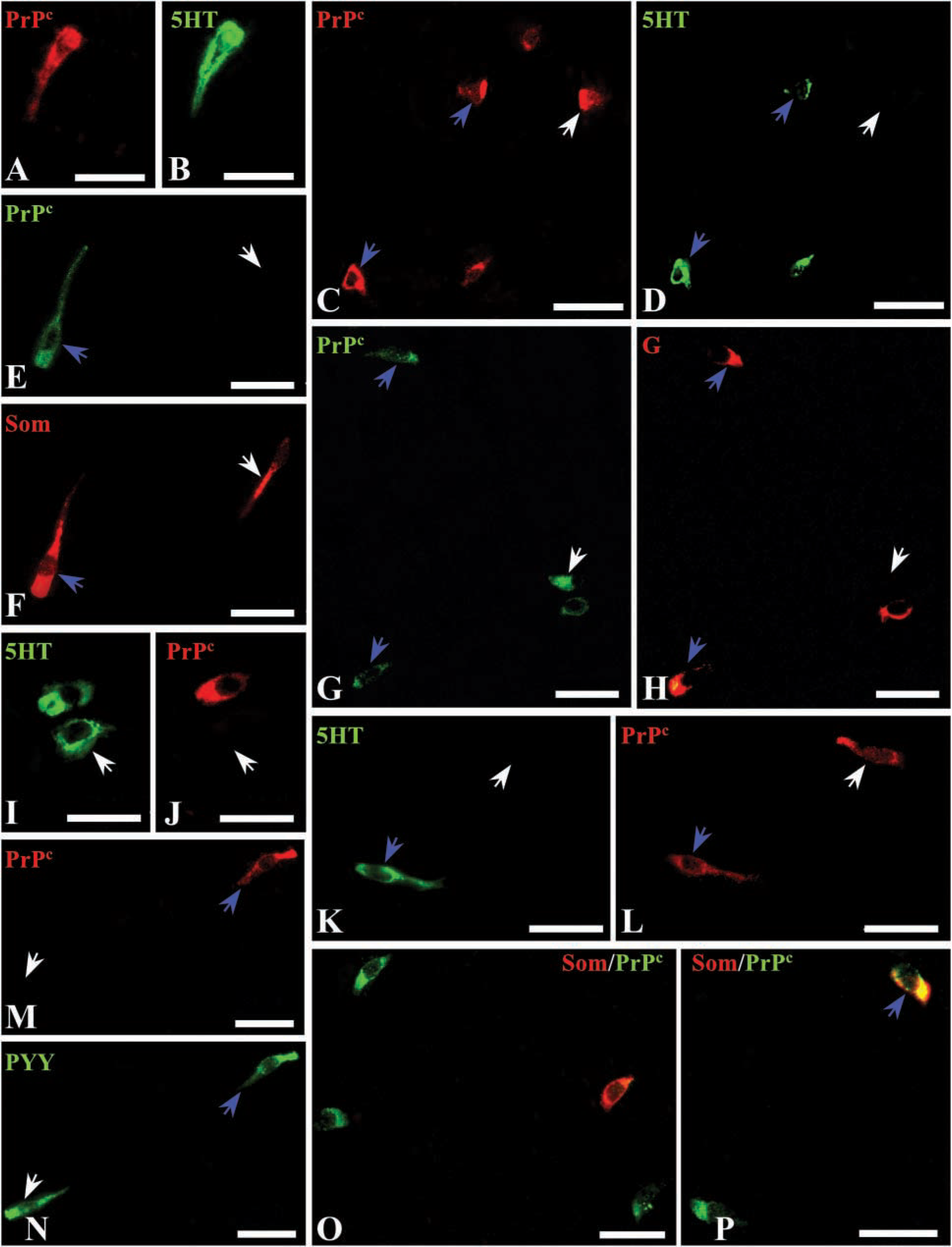
Double immunofluorescence for PrPc and neuroendocrine markers in rat gut. (
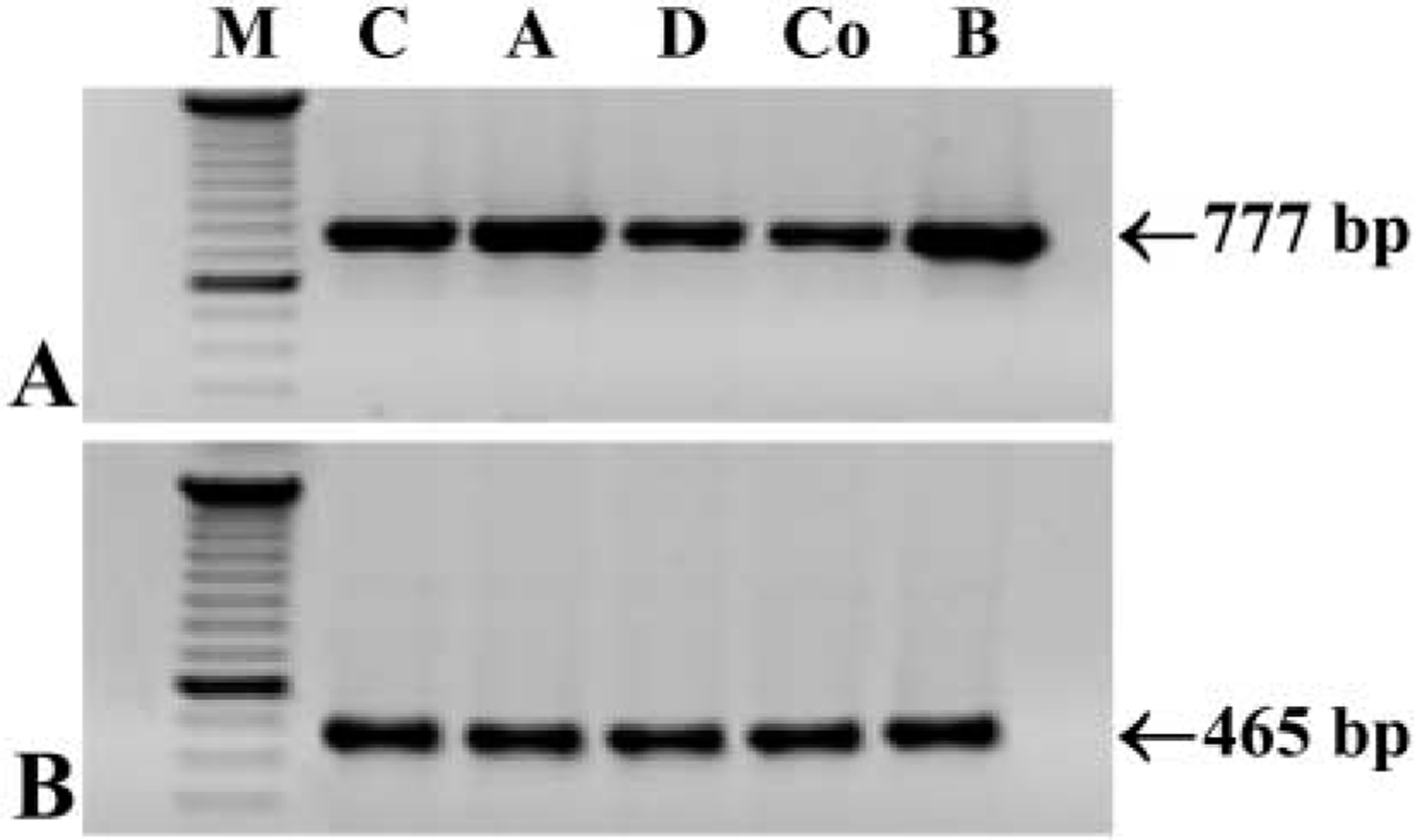
RT-PCR showing mRNA expression in all regions of the GI tract. (
The functional significance of the PrPc present in the neuroendocrine cells is not yet known. How the infectious agent crosses the gut wall and reaches germinal centers of lymphoid tissues or the peripheral nervous system also remains cryptic. The incorporation of PrPsc as a host PrPc-dependent process is a widely accepted idea because PrP knockout mice do not develop infection (Bueler et al. 1992). This is probably why some authors have pointed to enteroendocrine cells and other cell types, such as dendritic cells and intraepithelial lymphocytes, as the most probable PrPsc target cell populations (Ford et al. 2002).
With regard to PrPsc interacting molecules, the infectious agent could interact with PrPc expressed by GI epithelial cells, forming a large molecular complex that would later be incorporated (Horiuchi et al. 1995; Fournier et al. 1998). On the other hand, evidence has also pointed to the 37-kD laminin receptor precursor (LRP) as a PrPsc receptor (Rieger et al. 1997; Shmakov et al. 2000; Ghosh 2002). LRP expression in human intestinal brush border (Shmakov et al. 2000) and LRP–PrPsc interaction (Rieger et al. 1997) have been reported. PrPsc-receptor complexes would further reach the lymphoreticular system (van Keulen et al. 1996; Hill et al. 1999; Hilton et al. 2002) and peripheral nerves (Groschup et al. 1996; McBride and Beekes 1999) before spreading to the CNS. As a possible infection transfer mechanism between cells, an intercellular traffic of glycosylphosphatidylinositol-anchored PrPc has been described (Liu et al. 2002). This could explain PrPsc dissemination towards the nervous system once it crosses the epithelial barrier, because many nerve endings are present in tight contact with epithelial cells.
Only one theory exists about the role of enteroendocrine cells in the pathogenesis of spongiform encephalopathies. This theory postulates that changes in the properties of the GI epithelium might activate cells so that PrPsc could enter into them (Argenzio 1997). Here, PrPsc could replicate and then be secreted to reach the enteric nervous system or other non-enteric target cells throughout the bloodstream. In fact, a secretory form of PrPc has been reported (Hay et al. 1987). In addition, PrPc expression in secretory cells, such as neuroendocrine cells, supports the possible involvement of this protein in the regulated secretory pathway, as has been suggested by other authors who also found PrPc in exocrine cells (Bendheim et al. 1992; Fournier et al. 1998). Nevertheless, PrPc expression in only specific neuroendocrine cell subpopulations is an intriguing point. The physiological role of PrPc might be connected to regulatory substances secreted by some neuroendocrine cells, but further research needs to be conducted.
In summary, PrPc is present in some but not all neuroendocrine cell types along the rat GI tract. This specific pattern indicates both a highly regulated expression and a particular role of this protein in these cells. Moreover, our results suggest that neuroendocrine cells may play an important role in the internalization of the PrPsc from gut lumen. Nevertheless, further studies are needed to verify this hypothesis.
Footnotes
Acknowledgements
Supported by Departamento de Educación y Cultura del Gobierno de Navarra (and by a grant to Z.M.), the University of Navarra (PIUNA), and partially funded through the “UTE project CIMA.”
We thank Teresa Sabata, Paz Zamora, Silvia González, and Ainhoa Urbiola for technical assistance.