
Abstract
Serum amyloid A (SAA) is an acute phase protein which is expressed primarily in the liver as a part of the systemic response to various injuries and inflammatory stimuli; its expression in ovarian tumors has not been described. Here, we investigated the expression of SAA in human benign and malignant ovarian epithelial tumors. Non-radioactive in situ hybridization applied on ovarian paraffin tissue sections revealed mostly negative SAA mRNA expression in normal surface epithelium. Expression was increased gradually as epithelial cells progressed through benign and borderline adenomas to primary and metastatic adenocarcinomas. Similar expression pattern of the SAA protein was observed by immunohistochemical staining. RT-PCR analysis confirmed the overexpression of the
Serum amyloid a (SAA) is an acute phase protein, expressed primarily in the liver, whose level in the blood is elevated to 1000-fold in response to trauma, infection, inflammation, and neoplasia (Urieli-Shoval et al. 2000). The liver is the major site of SAA expression; however, extrahepatic expression has been described including in human atherosclerotic lesions (Meek et al. 1994), in synovial tissues from rheumatoid arthritis patients (Kumon et al. 1999), and in many histologically normal human tissues (Urieli-Shoval et al. 1998). The role of SAA in health and diseases is not well understood. In addition to its role as the precursor protein in AA-type amyloidosis and a role in cholesterol metabolism and transport, being an HDL-associated apolipo-protein, SAA is viewed as a modulator of inflammatory processes. Several biologic activities/effects of SAA have been described in relevance to inflammation. These include the following: inducing adhesion, migration, and tissue infiltration of inflammatory cells (Badolato et al. 1994; Xu et al. 1995); enhancing activity of matrix-degrading enzymes, matrix metalloproteinases (MMPs) (Migita et al. 1998; Lee et al. 2005), and plasminogen activators (PAs) (Michaeli et al. 2008); enhancing expression of inflammatory cytokines, chemokines, and growth factors (IL-1β, IL-6, TNFα, IL-8, G-CSF, COX-2) (Furlaneto and Campa 2000; Jijon et al. 2005; Koga et al. 2008); stimulating angiogenesis (Lee et al. 2006; Mullan et al. 2006); stimulating activity of inflammation-associated nuclear factor NFκB and signaling pathways mediated by mitogen-activated protein kinases ERK1/2, p38, and JNK (Jijon et al. 2005; Mullan et al. 2006).
In studies performed in the 1970s and 1980s, SAA was suggested as a nonspecific serum biomarker for neoplastic activity. SAA serum levels were found elevated in a wide range of malignancies, were higher in patients with distant metastases, decreased in response to therapy, and inversely correlated with patients' survival (Rosenthal and Sullivan 1979; Weinstein et al. 1984; Biran et al. 1986). Studies performed more recently, using modern mass spectrometry and proteomic technologies, confirmed these earlier studies and identified SAA as a major serum protein differentiating patients with cancer, including those with lung, ovarian, and pancreatic carcinomas (Moshkovskii et al. 2005; Yokoi et al. 2005; Liu et al. 2007) and melanoma (Findeisen et al. 2009). Whether the SAA is expressed locally in cancerous tissues was for many years not known. We had reported that SAA is locally expressed in colon carcinomas and that its expression increases in correlation with the progressive stages of colonic neoplasia (Gutfeld et al. 2006; Michaeli et al. 2008). More recently, overexpression of SAA in uterine serous papillary, renal cell, and endometrial carcinomas was reported by others (Cocco et al. 2009, 2010; Paret et al. 2009). Expression of SAA in ovarian tumors has not been described.
Ovarian cancer is one of the most aggressive cancer types in women, and epithelial ovarian cancer is the most common cause of mortality among gynecologic malignancies (Cannistra 2004). Primary ovarian epithelial tumors are divided into benign, borderline, and carcinoma, which are further classified into histological subtypes, including serous carcinoma - the most common subtype. Two pathways were proposed for the tumorigenesis of epithelial ovarian cancer. In one, low-grade carcinomas develop by a stepwise progression from ovarian surface epithelia to benign lesions and then to borderline tumors and to invasive carcinomas. In the second, high-grade carcinomas develop by transformation of the ovarian surface epithelia without intermediate stages (Singer et al. 2002; Landen et al. 2008). The pathogenesis of ovarian cancer has been closely linked to inflammatory processes associated with ovarian folliculogenesis and ovulation, and incessant ovulation has been suggested as a risk factor for development of ovarian cancer (Fleming et al. 2006). Most patients with ovarian cancer are diagnosed at advanced stages when conventional therapy is less effective, necessitating the need for identifying new diagnostic and therapeutic targets in ovarian cancer.
To date, there has been no report on the local expression of SAA in ovarian tumors. In this study, we investigated the expression pattern of SAA in human ovarian epithelial benign and malignant tumors, in ovarian carcinoma cell line, and the serum levels of SAA in patients with ovarian carcinoma in comparison with CA-125 and C-reactive protein (CRP). The data suggest the involvement of SAA in ovarian tumorigenesis.
Materials and Methods
Tissues and Sera
Conventional serial sections of routinely processed formalin-fixed and paraffin-embedded archival ovarian tissues from patients operated between 1998 and 2005 were obtained from the Department of Pathology, Hadassah-Hebrew University Medical Center. Ovarian tumors included the following: 10 benign epithelial tumors (8 serous cystadenomas, 2 serous cystadenofibromas; mean patient age 48 years, range 20-71 years), 9 serous borderline tumors (8 serous tumors, 1 mucinous tumor; mean age 28 years, range 20-42 years), 14 carcinomas (11 serous carcinomas, 9 high grade and 2 low grade; 1 mixed serous and endometroid carcinoma, high grade; 1 mixed clear-cell and transitional carcinoma, high grade; 1 undifferentiated, high grade; mean age 58 years, range 51-66 years), and 6 serous carcinoma metastases (3 to omentum, 2 to colon, 1 to stomach wall; mean age 62 years, range 52-72 years). Sections of normal ovaries were obtained from eight women (mean age 50 years, range 39-71 years). Freshly frozen ovarian biopsies, four normal and five carcinomas (three serous carcinomas, one endometroid carcinoma, one mixed mucinous and endometroid carcinoma) from nine patients (mean age 50 years, range 41-64 years), operated between 1997 and 2006, were obtained from the Department of Pathology, Hadassah-Hebrew University Medical Center.
Sera of patients with ovarian pathology were obtained from the Department of Clinical Biochemistry during 2005-2006. Ovarian pathology included the following: epithelial carcinomas—active disease (47 patients; mean age 63 years, range 39-85 years); epithelial carcinomas—under remission (12 patients; mean age 68 years, range 37-76 years); benign cysts (23 patients; mean age 46 years, range 19-68 years). The patient group with carcinoma—active disease included the following: 37 patients postsurgery and during adjuvant chemo treatment, 9 patients after histology confirmation of disease and before treatment, and 1 patient with active disease receiving only palliative care. Treatment for patients with carcinoma included mainly taxol and carboplatin as a first-line treatment and various other chemo agents as second-, third-, and fourth-line treatments. Studies were approved by the Human Subjects Research Committee of the Hadassah-Hebrew University Medical Center.
In Situ Hybridization
Non-radioactive ISH was performed as we previously described (Urieli-Shoval et al. 1998; Gutfeld et al. 2006). Briefly, SAA probe was prepared from pGEM transcription vector that contained a 110-bp sequence of mouse
Immunohistochemistry
IHC was performed using the Histostain-Plus SP kit (Zymed Laboratories; San Francisco, CA) as we previously described (Urieli-Shoval et al. 1998; Gutfeld et al. 2006). Briefly, two different anti-SAA monoclonal antibodies were used: clone mcl (Dako; Carpinteria, CA) and clone mc29 (provided by RP Linke). The preparation and specificity of these antibodies were previously described (Linke 1984; Linke et al. 1991; Urieli-Shoval et al. 2002). Antibodies were diluted 1:20 (mcl) and 1:600 (mc29) in 0.1 M Tris-HCl (pH 7.6), and incubation lasted for 2 hr at room temperature. The two anti-SAA antibodies yielded a similar staining pattern. For negative control, the primary antibodies were replaced by normal mouse isotype-matched IgG (IgG2a, κ; Dako), resulting in a diminished signal.
RT-PCR Analysis
RNA was extracted from freshly frozen ovarian biopsies, using TRI Reagent (Sigma-Aldrich; St Louis, MO), and cDNAs were synthesized using random hexamer primer and Superscript II RNase H- Reverse Transcriptase (Invitrogen by Life Technologies; Paisley, UK). The cDNAs were amplified using SuperTherm DNA polymerase (JMR Holdings; London, UK) and primers specific for the four known human SAA genes,
Detection of SAA Expression in OVCAR-3 Cells
OVCAR-3 cells were maintained in DMEM supplemented with 10% FBS (Biological Industries; Kibbutz Beit Haemek, Israel). Cells were subjected to RNA extraction and RT-PCR as described earlier and to protein extraction using RIPA lysis buffer containing protease and phosphatase inhibitors (UPSTATE; Lake Placid, NY). Concentrations of SAA protein and total protein in cell extracts were determined using SAA-specific ELISA (BioSource International; Camarillo, CA) and a modified Lowry protein assay (Sigma-Aldrich), respectively. Secreted SAA protein was detected upon concentration of conditioned media by Amicon Ultra centrifugal filters (Millipore; Billerica, MA) before SAA determination. Cell-associated and secreted SAA proteins were also determined in cells grown with cytokines (IL-β + IL-6 at 10 ng/ml each), as we described (Michaeli et al. 2008).
Measurement of Serum Levels of SAA, CA-125, and CRP
SAA serum levels were determined by the SAA-specific ELISA (BioSource International). Serum CA-125 levels were determined using Elecsys CA125 II immunoassay kit in an Elecsys 2010 analyzer (Roche Diagnostics; Indianapolis, IN). Serum CRP levels were measured by particle-enhanced turbidimetric test (COBAS INTEGRA 800 analyzer; Roche Diagnostics). Assays were performed according to the manufacturers' instructions.
Results
Expression of SAA in Ovarian Epithelial Tumor Tissues
To investigate SAA mRNA expression, non-radioactive ISH was applied on ovarian tissue sections representing normal, benign, borderline, carcinoma, and metastasis. IHC was applied on selected sections to document that the presence of SAA mRNA is accompanied by SAA protein synthesis. Our findings are described below. Representative results are demonstrated in Figures 1 and 2. Summary of SAA mRNA expression in ovarian tumor epithelium is given in Table 1.
Figure 1 and 2
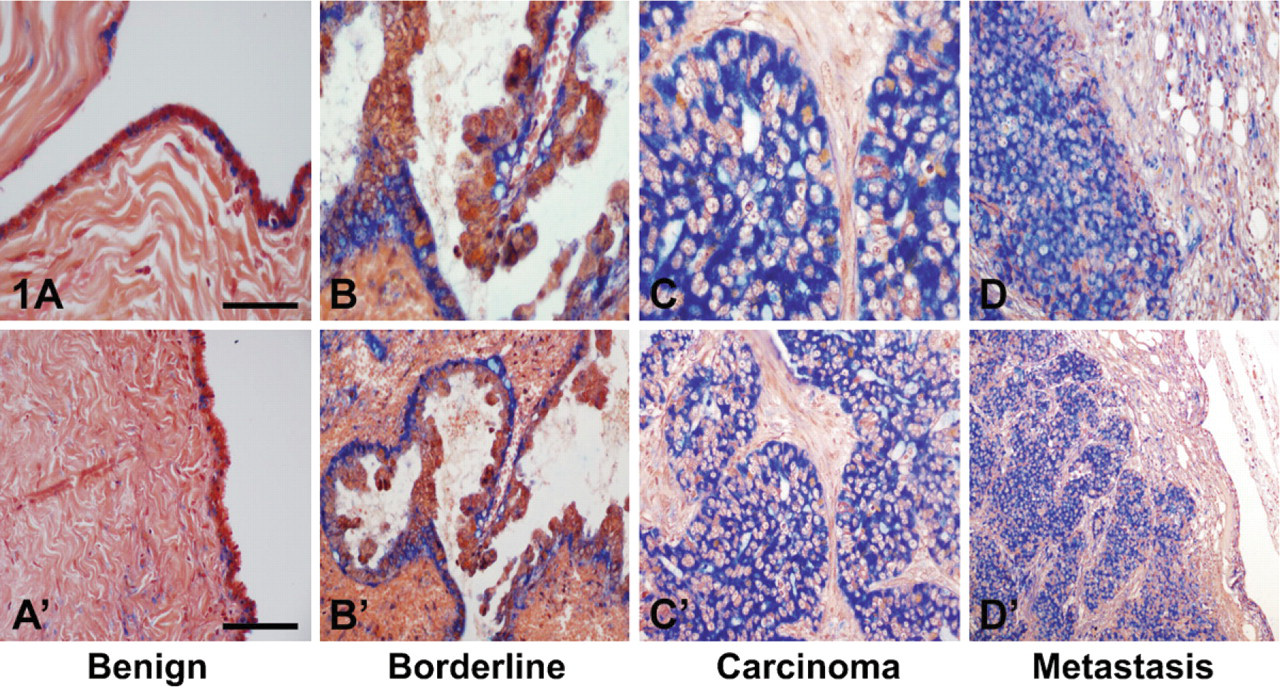
ISH demonstrating SAA mRNA expression in ovarian tumors. The blue cytoplasmic staining represents positive SAA mRNA signal; counterstaining is red-pink. (
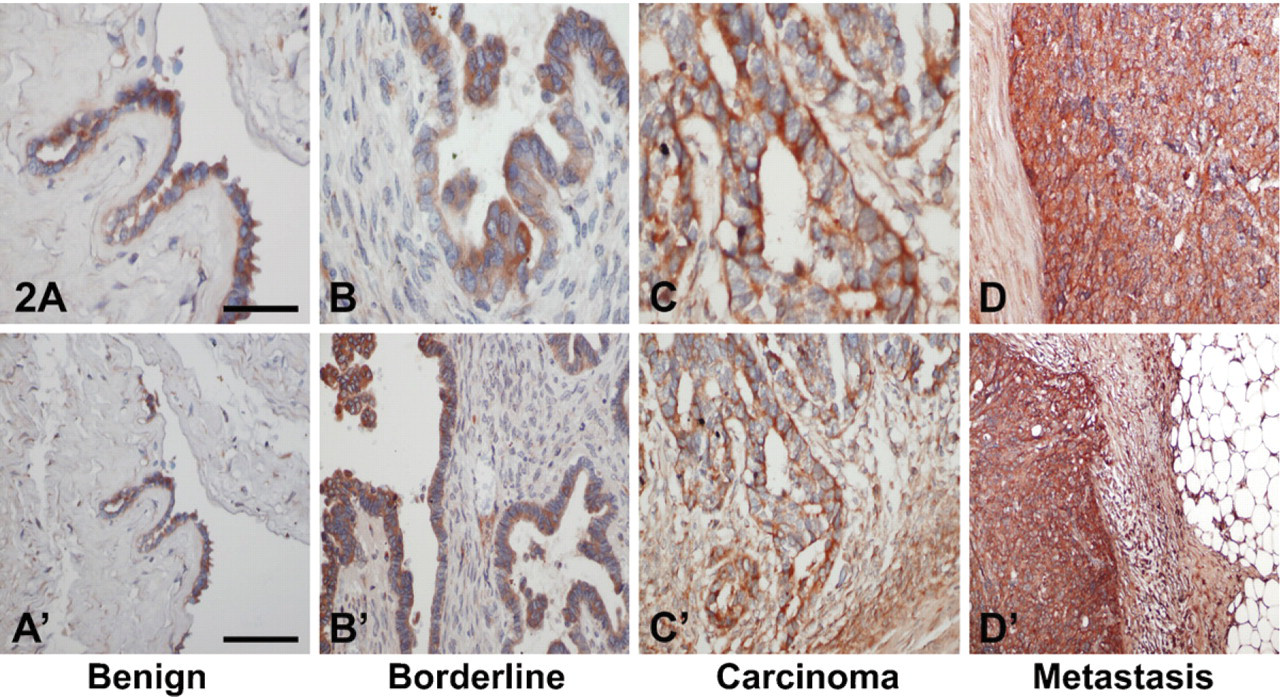
IHC demonstrating SAA protein expression in ovarian tumors. The sections were immunostained with monoclonal anti-SAA antibodies. The reddish-brown staining represents positive SAA protein signal; counterstaining is light blue. (A,A') Serous cystadenoma. The monolayer serous epithelium stained weak and focal. (
Summary of ISH staining of ovarian tumor epithelium
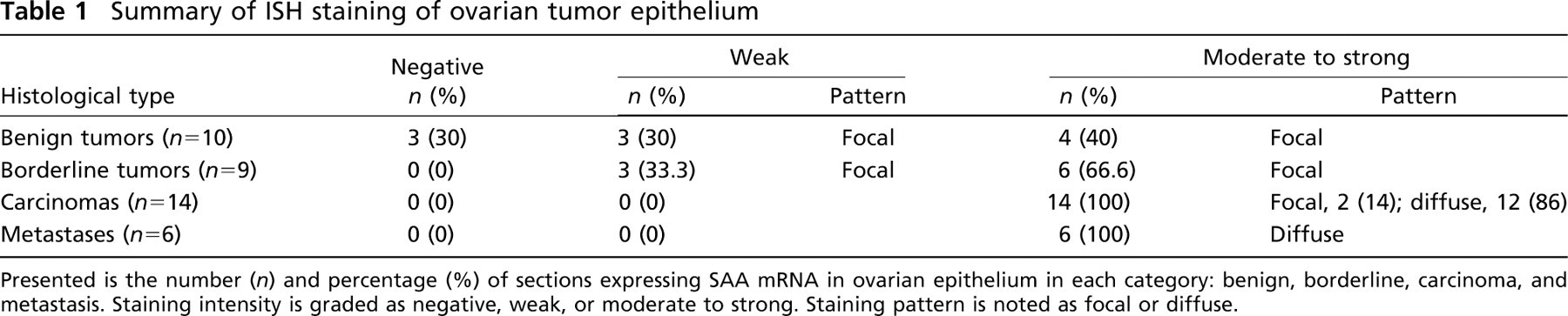
Presented is the number (
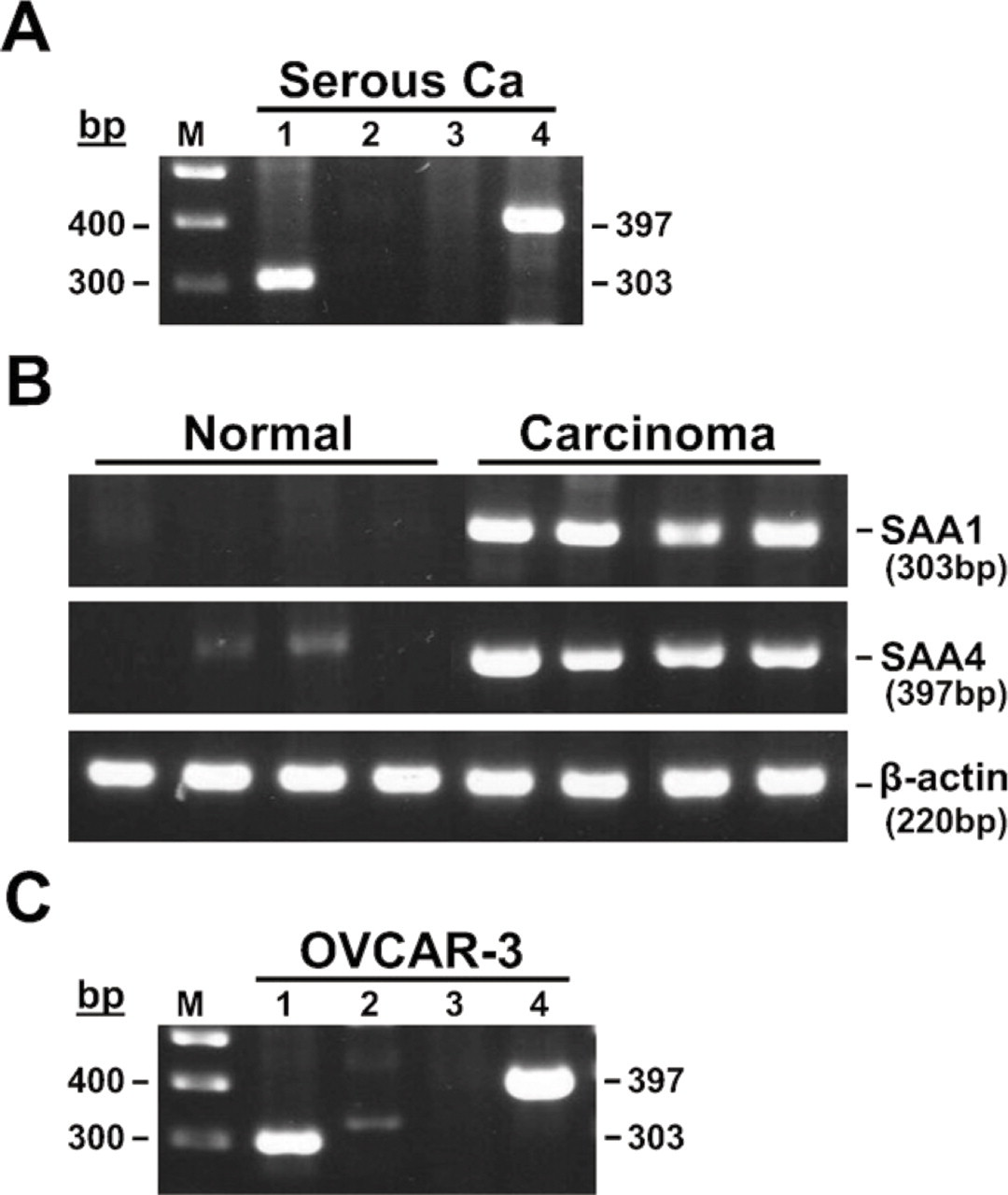
RT-PCR analysis of ovarian tissues and OVCAR-3 cells. (
RT-PCR of Ovarian Tissues
To further support the ISH findings and to determine which of the four known human SAA genes are expressed, we performed RT-PCR analysis on RNA extracted from freshly frozen ovarian biopsies. In Figure 3A, results obtained from ovarian serous carcinoma are shown. Fragments of the predicted sizes were amplified when the
Expression of SAA in Ovarian Cancer Cell Line OVCAR-3
We investigated the expression of SAA in ovarian carcinoma cell line OVCAR-3. RT-PCR analysis revealed expression of
SAA Serum Levels in Patients With Ovarian Carcinoma
We measured the SAA concentration in the sera of patients with ovarian pathologies, 47 with epithelial carcinomas—active disease, 12 with epithelial carcinomas—under remission, and 23 with ovarian benign cysts. In all the serum samples, concentrations of CA-125 and CRP were also measured (Figure 4). High levels of SAA were found in patients with carcinoma—active disease, which were accompanied by high levels of CA-125 and CRP. Strong correlation was found between levels of the three proteins (Pearson's correlation coefficient: 0.6, 0.73, and 0.76 for SAA vs CA-125, SAA vs CRP, and CRP vs CA-125, respectively). Low levels of SAA were found in patients with carcinoma—under remission, which were accompanied by low levels of CA-125 and CRP. Various correlations were found between levels of the three proteins (0.42, 0.59, and 0.86 for SAA vs CA-125, SAA vs CRP, and CRP vs CA-125, respectively). The SAA levels in these patients, although low, were significantly higher compared with levels in patients with benign cysts. In contrast, the CA-125 and CRP levels were within their normal values, with no significant difference between the two patient groups.
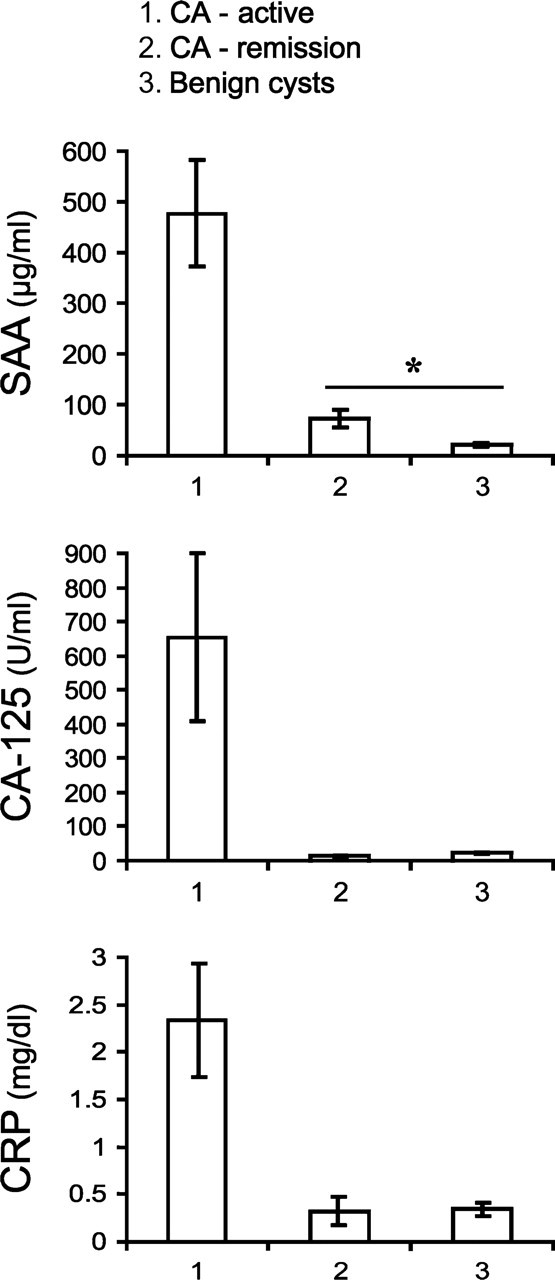
SAA serum levels in patients with ovarian carcinoma. Serum levels of SAA, CA-125, and CRP were measured in patients with ovarian pathologies classified as follows: (1) carcinoma—active disease (
Discussion
We describe here the local expression of SAA in human ovarian epithelial tumors, expression that is increased gradually as epithelial cells progress through benign and borderline adenomas to primary and metastatic adenocarcinomas. In addition, high serum levels of SAA were found in patients with ovarian carcinoma, levels that correlated strongly with levels of the ovarian tumor marker, CA-125, and the acute phase protein, CRP. The data suggest the involvement of SAA in ovarian tumorigenesis and may have prognostic and therapeutic applications.
SAA expression was studied by non-radioactive ISH and IHC. In normal ovaries, staining for SAA in the surface epithelium was mostly negative. In benign tumors, staining of the epithelium was mostly weak and focal. In borderline tumors, staining of the epithelium was mostly moderate to strong and focal. In malignant epithelium of carcinomas and metastases, staining was strong and diffuse. These results indicate positive correlation between SAA expression and the malignant state of ovarian epithelial cells. Positive correlation between epithelial SAA expression and the progressive stages of colonic neoplasia was also reported (Gutfeld et al. 2006). Furthermore, enhanced SAA expression was recently reported in other carcinomas, i.e., uterine serous papillary, renal cell, and endometrial carcinomas, and the authors suggest SAA as a novel biomarker for these cancers (Cocco et al. 2009, 2010; Paret et al. 2009). Whether SAA could be a tissue biomarker in ovarian cancer requires additional studies, determining SAA expression levels in ovarian tumors from a larger patient cohort in correlation with patient clinicopathological parameters.
The SAA gene family comprises four discrete loci containing two highly homologous genes,
We also found strong expression of the
Overexpression of SAA in ovarian carcinomas may contribute to high levels of circulating SAA. Indeed, we found high levels of SAA in sera of patients with ovarian carcinoma—active disease, which correlated strongly with high levels of CA-125 and CRP. In contrast, low levels of all three proteins were found in patients with ovarian carcinoma—under remission. The SAA levels in patients in remission, although low, were significantly higher when compared with levels in patients with benign cysts. The CA-125 and CRP levels, however, were within their normal values, with no significant difference between the two patient groups. This finding suggests a higher specificity for SAA compared with CA-125 and CRP in differentiating these two patient groups. Several studies have reported elevated serum concentrations of CRP in association with ovarian cancer, but only limited data are available for SAA. Two recent proteomic studies identified SAA as a major protein in the serum of patients with ovarian carcinoma (Moshkovskii et al. 2005; Helleman et al. 2008). Furthermore, SAA together with other serum biomarkers, including CRP in combination with CA-125, shows a significant increase in sensitivity for detection and monitoring progression of ovarian cancer when compared with CA-125 alone (Helleman et al. 2008; Edgell et al. 2010). Our data are consistent with these previous observations and add evidence that the high SAA serum levels may be derived from the ovarian carcinoma lesions and are not only the product of hepatocytes. Further investigation is required to elucidate the potential of SAA as a serum biomarker in ovarian cancer, alone or in combination with other biomarkers.
The role of the locally expressed SAA in human ovarian epithelial tumors is not known. However, in line with the concept that inflammation plays a causative role in ovarian tumorigenesis (Cannistra 2004; Fleming et al. 2006), SAA may enhance the inflammatory micro-environment of the ovarian tumors by inducing inflammatory cell recruitment (Badolato et al. 1994; Xu et al. 1995), cytokine production (Furlaneto and Campa 2000; Koga et al. 2008), and activity of inflammation-associated nuclear factors and signaling pathways (Jijon et al. 2005; Mullan et al. 2006). SAA may induce the adhesion and migration of ovarian tumor cells as it does for human leukocytes (Badolato et al. 1994; Xu et al. 1995) and enhance tumor cell proliferation, survival, and angiogenesis as has been shown in relevance to rheumatoid arthritis (Lee et al. 2006; Mullan et al. 2006). SAA may also enhance ovarian tumor cell invasion and metastatic spread by enhancing the activity of matrix-degrading enzymes, MMPs (Migita et al. 1998; Lee et al. 2005). Finally, expression of components of the plasminogen activation system, i.e., urokinase PA (uPA), its receptor (uPAR), and its inhibitor (PAI-1), is increased going from normal ovarian tissues, via benign and borderline adenomas, to primary and metastatic adenocarcinomas (Borgfeldt et al. 2001), similar to the expression pattern that we found for SAA. This concomitant expression raises the possibility of a functional relationship between the two systems. In line with this assumption, we had recently reported that exogenous SAA enhances plasminogen activation activity of HT-29 colon cancer cell line, implicating its role in colonic tumorigenesis (Michaeli et al. 2008). Whether any of these possibilities have relevance in the development of human ovarian cancer requires further investigation.
In conclusion, our data indicate the local expression of SAA in human ovarian epithelial tumors and the direct correlation between SAA intracellular expression and the malignant state of ovarian epithelial cells. In addition, high concentrations of SAA were found in the serum of ovarian carcinoma patients, which strongly correlated with levels of CA-125 and CRP. Although our data need further validation in future studies, they support the hypothesis that SAA is involved in ovarian tumorigenesis and is a candidate tissue and serum biomarker and perhaps a therapeutic target in ovarian cancer.
Footnotes
Acknowledgements
This work was supported by grants from the Israel Science Foundation (no. 686/00-1; to SU-S), the Israel Cancer Association, and the Sylvia and David Salzberg Cancer Research Fund (to SU-S and IB-S).
We thank R.L. Meek (The Heart Institute of Spokane) for the p125 probe.