
Abstract
Inhibins and activins are important regulators of the female reproductive system. A novel inhibin subunit, named βC, has been identified and demonstrated to be expressed in several human tissues. We demonstrate here that inhibin βC is expressed in human placenta. Expression of the inhibin βC subunit was demonstrated at the protein level by means of immunohistochemical evaluation and at the transcriptional level by an inhibin βC-specific RT-PCR analysis. Expression of inhibin βC was detected in the human chorionic carcinoma cell lines JEG and BeWo. Although the precise role of this novel inhibin subunit in human placenta development and homeostasis is unclear, analogies with other inhibin sub-units and the strong expression of βC in normal human trophoblast cells and chorionic carcinoma cells suggest that βC may be involved in autocrine/paracrine signaling pathways, angiogenesis, decidualization, and tissue remodeling under normal and malignant conditions. Additionally, JEG and BeWo express βC and, therefore, can be used as a cell culture model for further functional analysis of this subunit in the human placenta.
Keywords
I
Primarily isolated from the gonads, inhibins and activins are synthesized in several other gynecological organs, including normal and pathological human endometrium (Mylonas et al. 2004, 2009; Worbs et al. 2007) and normal and pathological placenta (Petraglia et al. 1991; Mylonas et al. 2006a,b,2007). During pregnancy, the well-characterized inhibin α, βA, and βB subunits are also expressed in the placental decidua, the syncytiotrophoblast, and the tropho-blast (Petraglia et al. 1991; McCluggage et al. 1998; Debieve et al. 2000), suggesting different roles for these subunits, such as paracrine modulators of reproductive function (Welt et al. 2002). Interestingly, higher in-hibin levels in human serum have been described in preeclampsia (Muttukrishna et al. 1997) and Down syndrome (Aitken et al. 1996), demonstrating a substantial function during pregnancy. Additionally, in-hibin α, βA, βB, and βC subunits, follistatin, β-glycan, and the activin receptor were detected in placental tissue from both uncomplicated term pregnancies and term pregnancies with preeclampsia (Casagrandi et al. 2003), consistent with elevated levels of inhibin A and activin A in maternal serum (Muttukrishna et al. 1997).
The novel inhibin βC subunit is predominantly expressed in rat and human livers (Esquela et al. 1997; Zhang et al. 1997; Vejda et al. 2003). A βC transcript was also demonstrated in human ovary, placenta, and testis (Loveland et al. 1996). The precise role of the βC subunit has not yet been elucidated. Mice deficient in the activin βC gene appeared normal and viable (Lau et al. 2000). In an immortalized mouse hepatocyte cell line (AML12), an increase in the rate of DNA synthesis by activin βC could be demonstrated (Wada et al. 2005). Additionally, a possible autocrine growth modulator function in liver regeneration has been suggested (Gold et al. 2005). Interestingly, ectopic expression of this subunit induced apoptosis in hepatoma cells (Chabicovsky et al. 2003; Vejda et al. 2003). Whether, and to what extent, the βC subunit can also act as a mitotic agent in human placental tissue is still unclear.
Only limited data on the histological expression of the inhibin βC subunit in normal human placenta exist. Moreover, the synthesis of this subunit in chorionic carcinoma cell lines BeWo and JEG is also quite unclear. These chorionic carcinoma cell lines are available now for over 30 years and are often used as in vitro model to evaluate normal and pathological placental endocrine function (Sullivan 2004). Therefore, the aim of this study was to evaluate the synthesis of the inhibin βC subunit in normal human placental tissue and in chorionic carcinoma cell lines BeWo and JEG.
Materials and Methods
Tissue Samples
Placental tissues were obtained from six placentas of women giving birth at the First Department of Obstetrics and Gynecology of the Ludwig–Maximilians-University Munich, Munich, Germany. Whether gestation can influence inhibin/activin production in the placenta is still controversial and, therefore, normal placental tissue samples were acquired. Tissue specimens of normal pregnancies were obtained in the course of an elective cesarean section for breech presentation, during the 38th week of gestation, to avoid any influencing factors due to the physiological “stress” of normal delivery (Mylonas et al. 2006a).
Immunohistochemistry
IHC was performed by a combination of pressure cooker heating and the standard streptavidin-biotin-peroxidase complex using the goat-IgG VECTASTAIN Elite ABC Kit (Vector Laboratories; Burlingame, CA) as previously described (Kimmich et al. in press). Briefly, paraffin-fixed tissue sections were dewaxed using xylol for 15 min and dehydrated twice in 100% ethanol. Endogenous peroxidase activity was quenched by immersion in 3% hydrogen peroxide (Merck; Darmstadt, Germany) in methanol for 20 min. After washing, the slides were subjected to antigen retrieval for 5 min in a pressure cooker using sodium citrate buffer (pH 6.0), containing 0.1 M citric acid and 0.1 M sodium citrate in distilled water. After cooling to room temperature, sections were washed twice in PBS. Nonspecific binding was blocked by incubating the sections with Ultra V Block (Lab Vision; Fremont, CA) for 45 min at room temperature. Sections were then incubated at 4C overnight with the inhibin βC polyclonal goat antibody (R&D Systems; Wiesbaden, Germany) at a dilution of 1:50 in Ultra V Block. After washing with PBS, sections were incubated with biotinylated anti-goat antibody (Vector Laboratories) for 30 min at room temperature. After incubation with the avidin–biotin–peroxidase complex (diluted in 10 ml PBS; Vector Laboratories) for 30 min and repeated washing with PBS, the sections were visualized using the ABC substrate buffer (VECTASTAIN Elite ABC Kit) and the chromagen 3,3'-diaminobenzidine (Dako; Glostrup, Denmark) at a concentration of 1 mg/ml for 2 min. Sections were counterstained with Mayer's acidic hematoxylin and dehydrated in an ascending series of ethanol (50-98%). After xylol treatment, sections were mounted. Negative controls were performed by replacing the primary antibody with normal goat IgG as an isotype control at the same dilution used for the primary antibody. Positive cells showed a brownish color, and negative controls, as well as unstained cells, showed a blue color. Sections were examined using a Leitz photo-microscope (Wetzlar, Germany). Digital images were obtained with a digital camera system (JVC; Yokohama, Japan) and were saved on computer (Diskus Software; Hilgers, Königswinter, Germany).
Cells and Cell Culture
The chorionic carcinoma cells BeWo and JEG were kindly provided by Dr. B. Ugele (First Department of Obstetrics and Gynecology, Ludwig–Maximilians-University Munich, Munich, Germany). Cells were cultured in Quantum 263 medium (PAA; Pasching, Austria) supplemented with antibiotics at 37C in a humidified atmosphere with 5% CO2 as previously described (Brüning et al. 2009; Kimmich et al. in press).
Immunofluorescence Analysis
Cells grown on glass cover slips were fixed with acetone for 10 min at room temperature and washed twice with PBS. Nonspecific binding was blocked by incubating the sections with Ultra V Block for 15 min at room temperature. Thereafter, the slides were incubated with the inhibin βC antibody (1:50 in dilution medium; Dako) overnight at 4C, followed by incubation with a Cy3-conjugated donkey–anti-goat antibody (1:500; Dianova, Hamburg, Germany) as previously described (Kimmich et al. in press). The slides were embedded in mounting buffer containing 4,6-diamino-2-phenylindole for staining the nuclei. Then, the slides were embedded with VECTASHIELD Mounting Medium (AXXORA; Lörrach, Germany) and examined with an Axiophot photomicroscope (Zeiss; Jena, Germany). Digital images were obtained with a digital camera system (AxioCam; Zeiss) and saved using the AxioVision software (Version 4.7; Zeiss).
RT-PCR Analysis
RNA was extracted from cells using the NucleoSpin RNA II kit (MACHEREY-NAGEL; Düren, Germany) as previously described (Kimmich et al. in press). Reverse transcription was performed with M-MLVreverse transcriptase and oligo(dT) (Promega; Mannheim, Germany) as recommended by the supplier. PCR was performed in an Eppendorf Mastercycler with GoTaq (Promega). Primer sequences were (5'–3') GCAGCCC-GGGTGAGAGTTGG (forward primer) and AC-TGCACCCACAGGCCTC (reverse primer), which amplify a 393-bp product of the inhibin βC cDNA, as previously described (Kimmich et al. in press). PCR cycling was performed after a 5-min initiation at 94C with 36 cycles of 1 min at 94C, 1 min at 57C, and 2 min at 72C, followed by a 5-min extension at 72C. For cDNA quality control, actin primers (Stratagene; Amsterdam, The Netherlands), which amplify a 661-bp product, were used. As a further control, cDNA was omitted (water control) to identify any PCR contamination. PCR products were separated on a 1.5% agarose gel with pBR328 ladder markers (Roth; Karlsruhe, Germany). Gels contained the SYBR Safe dye (1:10,000 dilution; Invitrogen, Karlsruhe, Germany), and after a complete run, they were transferred on a UV-permeable tray to a BioRad Image Analyzer (Munich, Germany). The image file was exported as a TIFF file and imported into PowerPoint to crop and label the figure.
Results
Immunolabeling of Inhibin βC
To test the specificity of the βC subunit antibody, evaluation of the immunohistochemical staining was performed using appropriate positive controls, including normal human liver specimens. The inhibin βC antibody (Figures 1A and 1B) stained hepatocytes, confirming previous results (Hötten et al. 1995; Schmitt et al. 1996; Zhang et al. 1997; Vejda et al. 2003). In normal placental tissue, inhibin βC was primarily expressed in the cytoplasm of extravillous trophoblast cells, whereas immunostaining in syncytiotrophoblast cells was moderate but weaker than trophoblast cells (Figures 2A–2D).
Expression Analysis of Inhibin βC in Human Chorionic Carcinoma Cell Lines by Immunofluorescence
Chorionic carcinoma cells are malignant cell lines derived from invasive trophoblast cells. We, therefore, tested the expression of inhibin βC in the human trophoblast cancer cell lines BeWo and JEG. Both cell lines expressed the novel βC subunit at the protein level, as demonstrated by positive immunohistochemical staining in the cytoplasm (Figures 3A and 3B).
RT-PCR Analysis
To analyze the inhibin βC subunit expression at the transcriptional level, RNA was extracted from samples of human placental tissue, transcribed into cDNA, and analyzed by PCR using specific primers. Figure 4 demonstrates that the mRNA of the inhibin βC subunit (393-bp PCR product) is expressed in normal placental cells. Further, expression of this subunit in the chorionic carcinoma cell lines BeWo and JEG was confirmed at the transcriptional level by RT-PCR analysis.
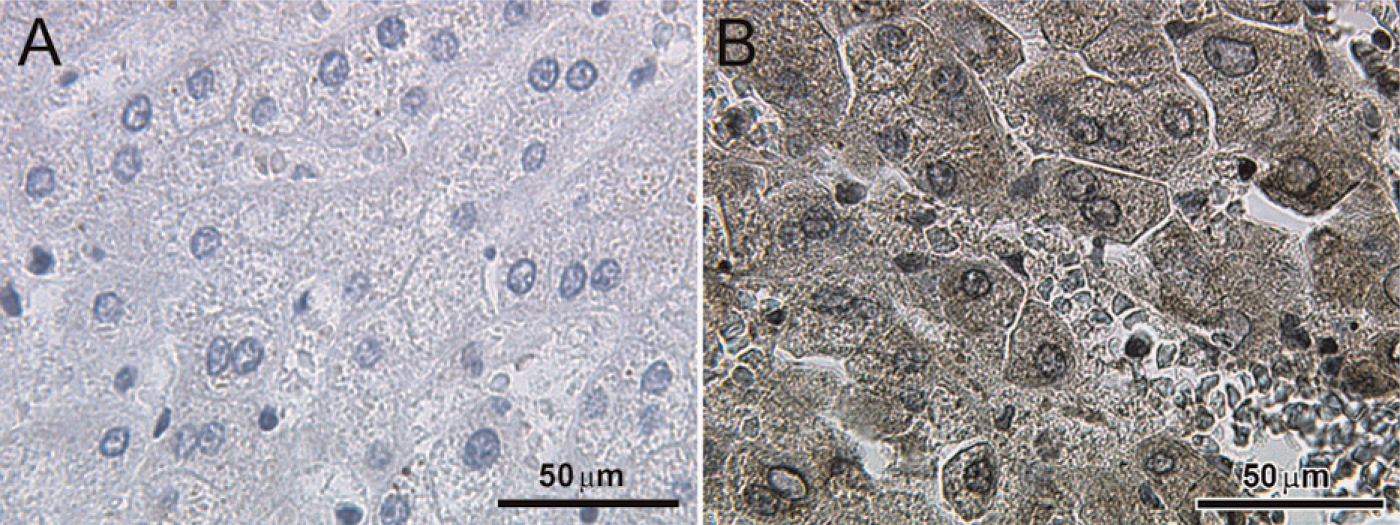
Immunohistochemical staining reaction of inhibin βC innormal human liver tissue. Normal human liver tissue was used as a positive control for inhibin βC immunolabeling. Negative controls were performed by replacing the primary antibody with normal goat IgG as an isotype control.
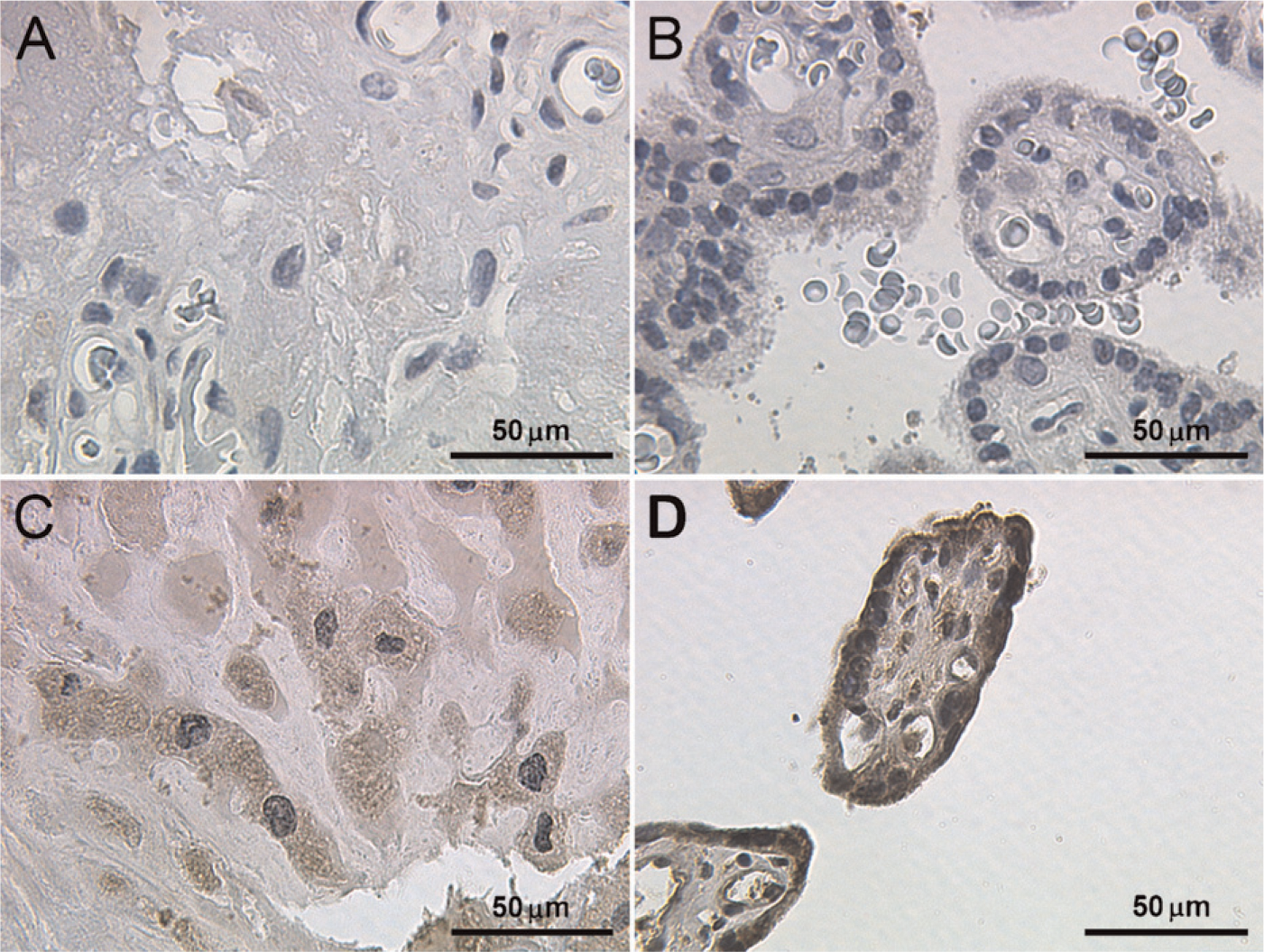
Immunohistochemical staining reaction of inhibin βC in normal human placenta tissue. No staining was detected in the negative controls for trophoblast cells
Discussion
Inhibins and activins were initially characterized as endocrine and paracrine hormonal regulators of the hypothalamic–pituitary–gonadal axis. In addition, the human placenta expresses inhibin/activin mRNA (Petraglia et al. 1991), and inhibin/activin subunits are thought to be the primary source of maternal circulating inhibin and activin (Florio et al. 2001). Although the precise physiological roles of placental inhibins and activins are still unclear, possible functions could include decidualization (Jones et al. 2006), trophoblast differentiation (Caniggia et al. 1997), immunomodulatory functions (Keelan et al. 2000), steroidogenesis (Ni et al. 2000; Ferreira et al. 2008), and apoptosis (Chen et al. 2002). Moreover, these proteins are intriguing as possible prognostic serological markers, because inhibin subunits and the entire inhibin or activin proteins have been associated with placenta-associated diseases such as preeclampsia and HELLP syndrome (Muttukrishna et al. 1997; Craig et al. 2000; Seufert et al. 2004; Mylonas et al. 2006a; Grill et al. 2009).
The inhibin/activin βC subunit is predominantly expressed in hepatocytes (Hötten et al. 1995; Schmitt et al. 1996; Zhang et al. 1997; Vejda et al. 2003). However, this subunit has been detected in cells from several other organs, including normal or malignant prostate and ovarian, testicular, endometrial, and pituitary tissues (Loveland et al. 1996; Mellor et al. 2000; Gold et al. 2004,2009; Kimmich et al. in press). The inhibin βC gene was detected in placental tissue from uncomplicated term pregnancies and pregnancies with preeclampsia (Casagrandi et al. 2003). We have demonstrated the presence of the inhibin βC subunit at the transcriptional level and at the protein level in the normal human placenta. Additionally, chorionic carcinoma cell lines also express this novel subunit. These cell lines have become valuable in vitro tools for studying placental physiology, invasion, and differentiation (King et al. 2000; Al-Nasiry et al. 2007; Serrano et al. 2007). Moreover, knowledge of the expression patterns of the β subunits has become tremendously important, because activin signaling may be a promising molecular therapeutic target for a variety of diseases, including metabolic diseases and malignant tumors (Tsuchida et al. 2009). Because the precise function of this novel subunit in human reproduction is still unclear, these cell lines can be used for further analysis of normal and pathological placental functions.
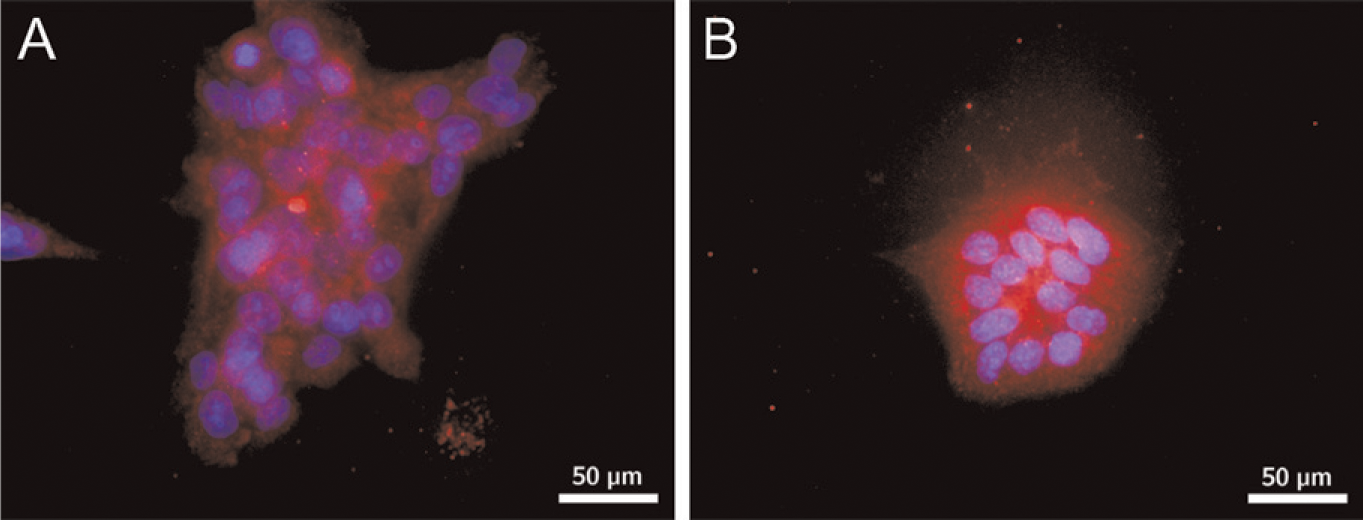
Localization of inhibin βC in BeWo and JEG. The carcinoma cell lines were analyzed by immunofluorescence for expression of inhibin βC. Positive cytoplasmic staining was detected in BeWo
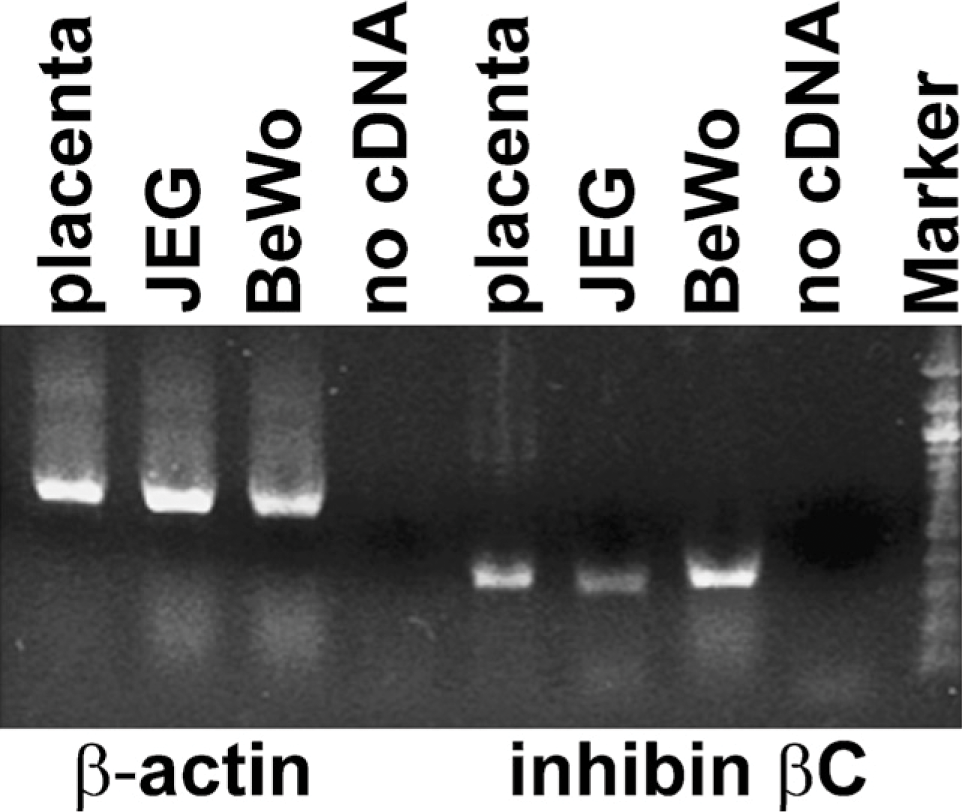
Inhibin βC expression in normal placental tissue and chorionic carcinoma cell lines. Normal placental tissue and BeWo and JEG cells were analyzed by RT-PCR for the expression of the inhibin βC subunit. PCR products from inhibin βC mRNA were detected in each sample. Interestingly, the inhibin βC mRNA level was less intense in JEG cells compared with BeWo cells and placental tissue. Furthermore, the water control to demonstrate any PCR contamination was negative.
The precise role of the βC subunit has not yet been elucidated. Mice deficient in the activin βC gene, the activin βE gene, or both appeared normal and viable (Lau et al. 2000), leading to the hypothesis that this subunit has a minor role in human reproduction (Gold et al. 2009). Therefore, it is not surprising that the research focus has concentrated on liver function. In an immortalized mouse hepatocyte cell line (AML12) and in primary rat hepatocytes, activin βC increases the rate of DNA synthesis (Wada et al. 2005). Moreover, the βC subunit was identified as an autocrine growth modulator in liver regeneration, leading to mitosis in a subset of hepatocytes (Gold et al. 2005). However, ectopic expression of this subunit induced apoptosis in hepatoma cells (Chabicovsky et al. 2003; Vejda et al. 2003). Whether, and to what extent, the βC subunit can also act as a mitotic agent in human placental tissue is still unclear.
One major issue remains unresolved, the question of putative inhibin or activin synthesis and the physiological function of these proteins in vivo. The formation of homodimeric activin C (βC–βC), heterodimeric activins AC (βA–βC), BC (βB–βC), and CE (βC–βE), and inhibin C (α–β) has been demonstrated by ectopic expression of the respective subunits in different cell models (Mellor et al. 2000; Ushiro et al. 2006). Interestingly, activin C (βC–C) does not activate activin A (bA–bA)-responsive promoters, suggesting that the βC subunit regulates the levels of bioactive activin A (βA–βA) through the formation of signaling-incompetent activin AC heterodimers (Mellor et al. 2003; Butler et al. 2005; Gold et al. 2009). Therefore, the βC subunit might function as an antagonist of activin function and thereby regulate activin signaling (Mellor et al. 2000, 2003). However, data that confirm secretion of the intact inhibins or activins are still lacking because of the lack of appropriate immunoassays.
In conclusion, inhibin βC mRNA and protein were detected in normal placental tissue by immunohisto-chemical and molecular assays. Additionally, chorionic carcinoma cell lines, which are often used to evaluate normal placental function, also express this novel sub-unit. However, the functional role of inhibin in the normal and pathological human placenta is still quite unclear, and therefore, inhibin subunits may have pleiotropic but distinct roles during pregnancy and gestational diseases.
Footnotes
Acknowledgements
This study was partially supported by the FöFoLe Program of the Ludwig–Maximilians-University Munich (297/03), the Friedrich–Baur-Institute of the Ludwig–Maximilians-University Munich, and the Weigland Stipendium Program of the Ludwig–Maximilians-University Munich (to IM). A.B. and I.M. are supported by the German Research Foundation (Deutsche Forschungsgemeinschaft, DFG BR 3641/3-1).
We thank Mrs. C. Kuhn, Mrs. S. Kunze, Mrs. S. Schulze, and Mrs. I. Wiest for their excellent work with the placental samples. Additionally, we thank Prof. Jeschke and Prof. Friese for their help in conducting this study, and Mr. P. Fuchs for his help with the images.