
Abstract
It has been assumed for a long time that except for limited areas close to respiratory bronchioles or their satellite arteries, there is no evidence of lymphatic vessels deep in the pulmonary lobule. An immunohistochemical study using the D2-40 monoclonal antibody was performed on normal pulmonary samples obtained from surgical specimens, with particular attention to the intralobular distribution of lymphatic vessels. This study demonstrated the presence of lymphatics not only in the connective tissue surrounding the respiratory bronchioles but also associated with intralobular arterioles and/or small veins even less than 50 μm in diameter. A few interlobular lymphatic vessels with a diameter ranging from 10 μm to 20 μm were also observed further away, in interalveolar walls. In conclusion, this study, using the D2-40 monoclonal antibody, demonstrated the presence of small lymphatic channels within the normal human pulmonary lobules, emerging from interalveolar interstitium, and around small blood vessels constituting the paraalveolar lymphatics. This thin intralobular lymphatic network may play a key pathophysiological role in a wide variety of alveolar and interstitial lung diseases and requires further investigation.
L
To our knowledge, apart from one very short comment (Pusztaszeri et al. 2006), there are no published reports of immunohistochemical studies of the pulmonary lymphatic network under normal conditions in humans. For this article, an immunohistochemical study using the D2-40 monoclonal antibody on normal human pulmonary samples was carried out, with particular attention to the depth of the pulmonary lobule.
Materials and Methods
Normal Lung Samples
Normal lung tissues were obtained from surgical specimens of five patients who underwent pulmonary resection for curative intent [lobectomy (n:3) and pneumonectomy (n:1) for non-small-cell lung carcinoma, or large wedge excision for secondary bowel adenocarcinoma (n:1)]. Tissues were fixed in 10% formalin (i.e., 3.7% formaldehyde and 1% methanol) for 18–24 hr at room temperature. After lung dissection, normal lung parenchyma on gross examination was selectively sampled from at least two sites situated away from the tumor, then immersed in the same fixative for 6 hr and processed for paraffin embedding as usual. Five-μm sections were stained with HE for routine histology, and normal lung sections were selected for histological examination. This retrospective study was conducted according to French legislation for medical research. Informed consent was considered unnecessary for this type of study.
Immunohistochemistry
D2-40 and Anti-CD34 Monoclonal Antibodies
The D2-40 monoclonal mouse IgG1 antibody recognizes podoplanin, a 38-kDa transmembrane glyco-protein expressed on lymphatic vascular endothelium (Breiteneder-Geleff et al. 1997) with both high specificity and high sensitivity (Evangelou et al. 2005; Schacht et al. 2005).
The anti-CD34 monoclonal mouse IgG1 antibody recognizes a heavily glycosylated type I 110-kDa transmembrane molecule, considered to be a marker for blood vessel endothelium. It is also occasionally expressed in tumor-associated lymphatic endothelial cells (Fiedler et al. 2006). Normal mouse IgG1 of the same isotype as the D2-40 and anti-CD34 antibodies was used as a control.
Immunostaining
After deparaffinization and rehydration in graded alcohol series, sections were labeled using an automated immunostaining procedure (Ventana Medical Systems; Tucson, AZ). Briefly, antigen retrieval was obtained by boiling at 95–100C. The D2-40 monoclonal mouse anti-podoplanin antibody (Dako, clone 07 3611; 1:100 dilution, concentration 0.25 mg/l−1) was incubated for 32 min, and the mouse monoclonal anti-CD34 (Dako, clone 96 3820 B; 1:100 dilution, concentration 0.5 mg/l−1) was incubated for 60 min at 42C. Sections were then incubated with the universal anti-mouse immunoglobulin biotinylated secondary antibody (Ventana SA; France) for 16 min at 37C, followed by incubation with the avidin-biotin-horseradish peroxidase (HRP) complex using the Ventana Medical Systems Basic DAB detection kit for 8 min at 37C. The final revelation system (Dako DMKiVIEW DAB) used HRP-DAB as the final chromogen. As negative controls, sections from each paraffin block were incubated with a normal mouse IgG1 (Dako; X0931 culture supernatant) instead of the primary antibody. Slides were counter-stained with hematoxylin.
Microscopic Analysis
First, the visceral pleura, peribronchiolar and perivascular interstitial sheaths, and interlobular septa were screened to determine the presence of D2-40–immunolabeled open-lumen lymphatic vessels on all pulmonary sections observed.
Second, the intralobular distribution of lymphatic vessels was selectively investigated by the two observers (MK and JFB). Image acquisition of the D2-40–immunostained slides was performed using a Zeiss microscope equipped with a Sony CCD digitization video camera (Figures 1–3). A calibrated grid (as shown in Figure 3A) was superimposed on each computerized image of the sections observed using the X10 objective, and their sections were entirely digitized.
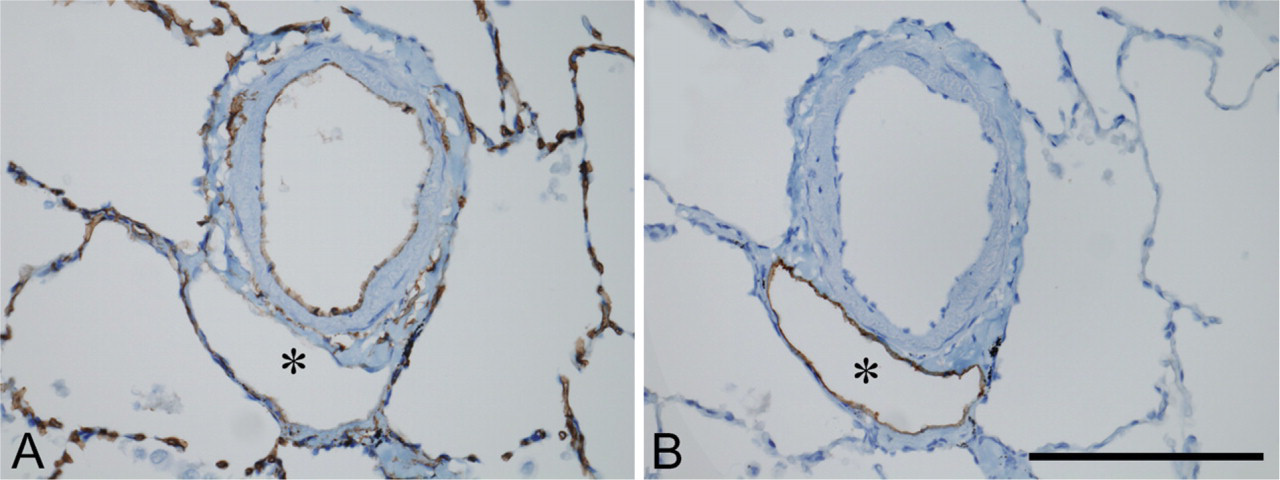
Immunohistochemical results on consecutive sections from a sample of normal human lung showing an intraacinar small pulmonary artery associated with a lymphatic vessel (asterisk). (
The lymphatic vessels were analyzed according to their location within the lobule: (1) associated with respiratory bronchioles; (2) associated with blood vessels less than 50 μm in diameter, between 50 μm and 100 μm, and more than 100 μm in diameter, respectively; and (3) not associated with either respiratory bronchioles or blood vessels, referred to as interalveolar lymphatic vessels. The intralobular blood vessels away from bronchioles less than 50 μm in diameter, between 50 μm and 100 μm, and more than 100 μm in diameter were also analyzed individually for the presence of a lymphatic vessel in their vicinity, which was reported.
Statistical Analysis
Data concerning the presence of lymphatic vessels according to the diameter of the adjacent blood vessels were compared using a Student's t-test.
Results
D2-40 and Anti-CD34 Immunostaining
Endothelial cells of lymphatic vessels observed in the interlobular septa and in the submesothelial space, characterized by the presence of valves, as those localized in the peribronchiolar connective interstitial tissue or perivascular interstitial tissue, were positively stained with D2-40 but did not react with the anti-CD34 antibody (Figure 1). Vessels of various diameters, with D2-40–positive endothelium, were also observed within the pulmonary lobule (Figures 2 and 3). Endothelial cells lining alveolar capillaries, intralobular arteries (>100 μm in diameter), or arterioles, as well as small veins (<100 μm in diameter) strongly expressed CD34 but were not stained with D2-40 (Figures 1A and 1B and Figures 2A and 2B). Epithelial basal cells of the bronchial and/or bronchiolar mucosa were immunostained with D2-40 (data not shown). No labeling was observed after incubation with the isotypic IgG1 controls (not shown).
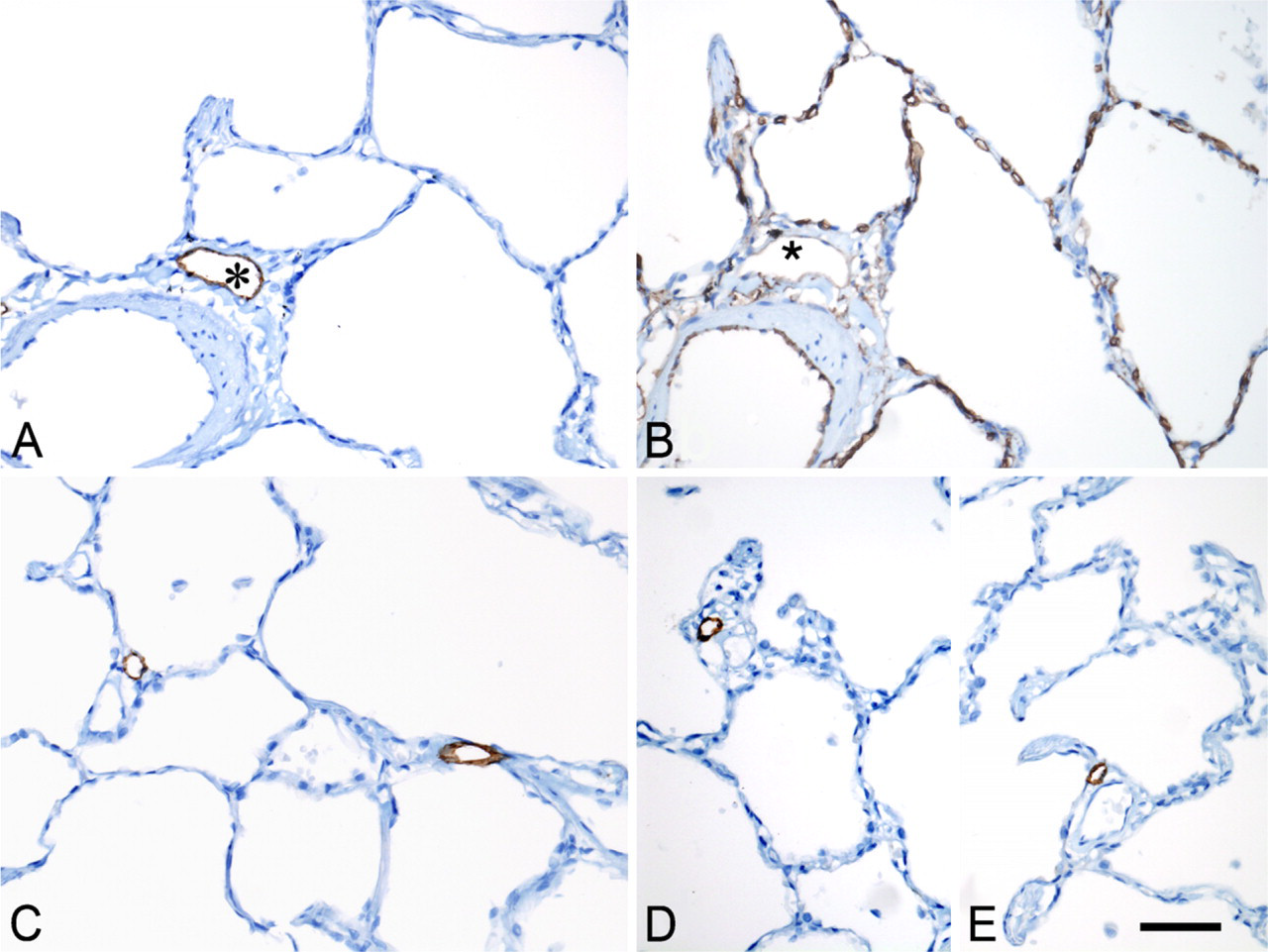
Immunohistochemical results on consecutive sections (
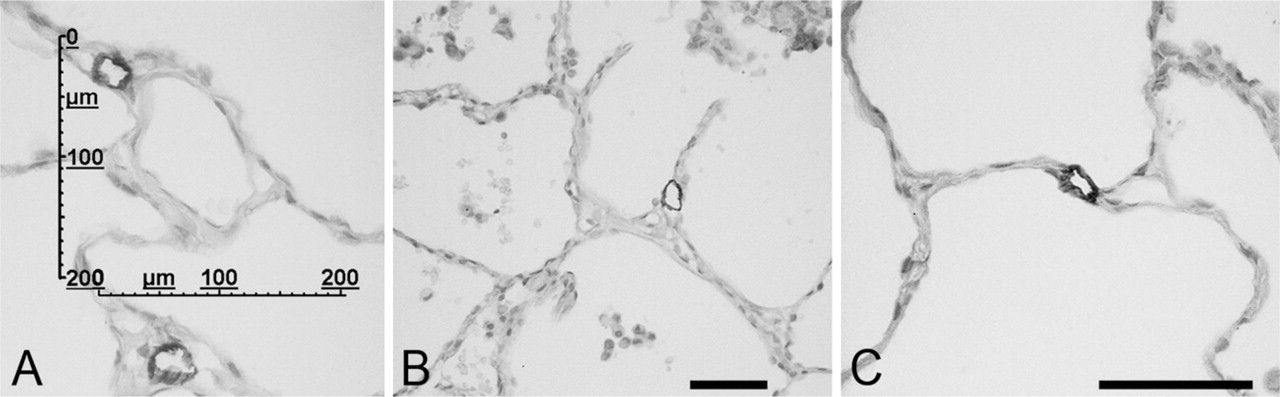
Immunolabeling after D2-40 incubation. (
Intralobular Distribution of Lymphatic Vessels
Lymphatic vessels identified by D2-40 immunostaining were constantly observed in the connective tissue sheath surrounding the respiratory bronchioles present on the sections (not shown). Lymphatic vessels were also observed in association with intralobular arterioles and/or small veins (Figures 2 and 3), in or very close to their juxtaadventitial interstitial sheath. A mean of 126 (range: 81–153) intralobular D2-40–stained lymphatic vessels were examined in each of the five samples. As shown in Figure 4, the percentage of blood vessels associated with a lymphatic vessel decreased significantly (p = 0.0004) according to the diameter of the blood vessel. Interalveolar septa were mostly devoid of lymphatic vessels, except for a very small number of septa, i.e., 3.6% to 19% of D2-40–immunostained intralobular lymphatic vessels were seen at a distance from either bronchioles, intralobular arterioles, or small veins within interalveolar walls (Figures 2 and 3B). The diameter of these intralobular lymphatic vessels ranged from 10 μm to 20 μm.
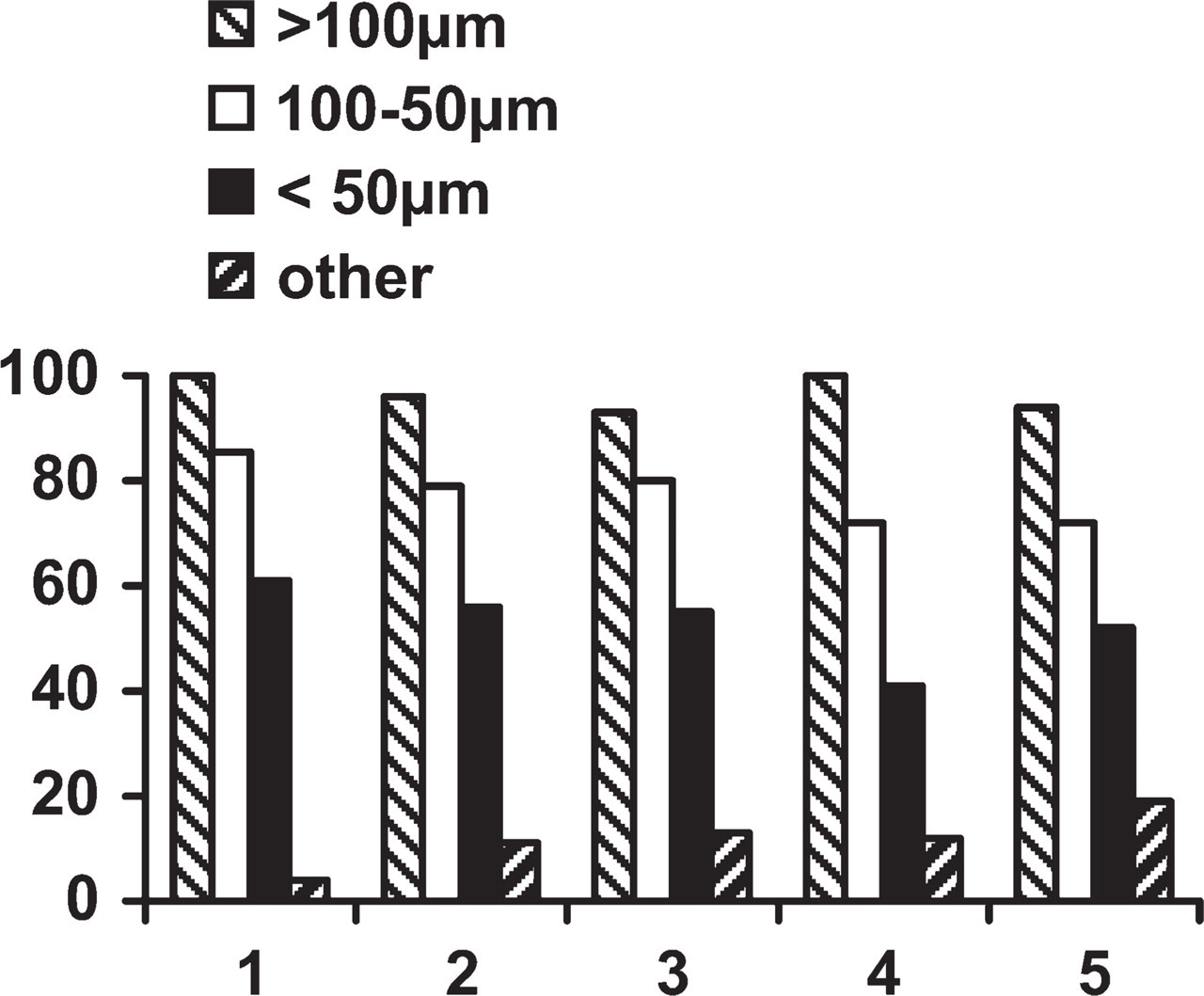
Distribution of the percentages of intralobular arteries (diameter >100 μm), arterioles, and/or small veins (diameter 100 μm to 50 μm and <50 μm) associated with a lymphatic vessel in samples from five normal lungs, indicated on the x-axis as 1 to 5. “Other” indicates the percentage of D2-40 intralobular-labeled lymphatics within the interalveolar septa. The differences observed between >100 μm, 100 μm-50 μm, and <50 μm were highly significant (p = 0.0004; Student's t-test).
Discussion
For a long time, reliable distinction of lymphatics from blood vessels on histological sections remained a major obstacle to precisely describing the lymphatic network. Over the last decade, the discovery of a spectrum of lymphatic markers, some of which are available on paraffin-embedded material, has made a major contribution to this field. One of these markers, the D2-40 antibody, has demonstrated very high specificity and sensitivity for identification of lymphatic endothelial cells in various organs (Van der Auwera et al. 2005; Pusztaszeri et al. 2006). It is also expressed by cells of various origins (Kono et al. 2007; Ishida et al. 2008), including the basal cell layer of the bronchial and/or bronchiolar mucosa in the present study. Pusztaszeri et al. (2006) confirmed that CD34 was expressed on pulmonary endothelial cells from arteries to capillaries and veins, but they found occasionally and irregularly weak staining with anti-CD34 antibody in lymphatic endothelium.
Pulmonary lymphatic vessels are classically described along the peribronchial vascular sheaths, inter-lobular septa, and pleural connective tissue, although their presence within the lobule, the deeper part of the lung parenchyma, is still open to debate, particularly in regard to the presence or absence of true alveolar lymphatics (Lauweryns 1971; Nagaishi 1972; Okada et al. 1979; Leak 1980). The pulmonary lobule, 0.5 cm to 1 cm wide, demarcated by connective interlobular septa, contains respiratory bronchioles, branches of arteries and veins (Von Hayek 1960), and interalveolar septa, comprising a rich pulmonary capillary network. By extrapolation with the lymphatic distribution observed in various mammalian species, it has been assumed that except for limited areas close to respiratory bronchioles or pulmonary arteries, there was no evidence of lymphatic vessels deep in pulmonary lobules (Schraufnagel 1992; Peão et al. 1993; Marchetti et al. 1994). Consequently, lymphatic drainage is considered to be non-functional in the intralobular pulmonary parenchyma, compared with other tissues. However, the presence of lymphatics deep in the lobules has been suspected by some authors: Leak (1980) described juxta-alveolar lymphatics, and Von Hayek (1960) described small lymphatic vessels in all parts of the lung containing loose interstitial connective tissue. Pusztaszeri et al. (2006) recently reported, in a short comment in a general study, that D2-40–labeled pulmonary lymphatics were seen “around veins, arterioles and bronchioles but they were very rare or absent in the alveolar wall.” In the present study, we identified lymphatic vessels on the basis of their open lumen and D2-40–immunolabeled endothelium, accompanying the majority of intralobular arteries, arterioles, veins, and venules. An average of 53% of blood vessels less than 50 μm in diameter, arterioles, and veins were associated with an adjacent lymphatic capillary and can be considered to be paraalveolar lymphatics (Weiss 1988). Moreover, an average of 10% of intralobular sections of lymphatic capillaries, not taking into account those located in the peribronchiolar interstitial spaces, were situated away from arterioles or venules, at the origin of interalveolar septa. However, it must be stressed that because the pulmonary lobule is a complex three-dimensional structure, it cannot be formally excluded that a certain proportion of the lymphatics observed in interalveolar septa may actually correspond to paraalveolar lymphatics situated near a small blood vessel not seen on the histological section.
The lymphatic system acts primarily as a transport system designed to maintain homeostasis of the interstitial environment by draining excess fluids and proteins from the interstitium for its return to the blood (Leak 1980; Swartz 2001). According to some authors, the pulmonary lymphatic system plays a role of removal of fluids from the abundant peribronchovascular or pleural connective tissue. It is also believed that fluid normally leaks out of capillaries located in the alveolar walls, moves out from very small septal interstitial spaces, and enters lymphatic capillaries in the vicinity of interstitial spaces and thereafter reaches larger lymphatics (Taylor and Parker 1985). The role of the deep distribution of intralobular lymphatics of the deep distribution of intratitial fluid and proteins very close to or within the interalveolar septa. Inter-alveolar interstitium would therefore play the role of prelymphatics, which are non-endothelialized tissue channels leading to the initial lymphatics as described elsewhere in the body (Castley-Smith 1976).
The intralobular lymphatic distribution observed in the present study could also explain the features observed in certain diseases, such as acute lung injury (Hedenstierna and Lattuada 2008; Mandal et al. 2008), or granulomatous diseases, such as tuberculoid granulomas. In an experimental model of Mycobacterium tuberculosis infection (Basaraba et al. 2006), it was clearly shown that inflammatory cells rapidly infiltrate the lymphatic walls and fill the lymphatic vessel lumens. These authors emphasized that pulmonary lymphatics not only constitute the initial site of M. tuberculosis dissemination but also develop progressive inflammation of their wall.
In conclusion, the present study using D2-40 immunostaining demonstrates that small lymphatic channels are observed within the lobules of normal human lung in the interstitial space around small blood vessels and occasionally emerging from the interalveolar interstitium. They ensure the drainage of excess proteins and fluid in this part of the lung. Studies using the D2-40 antibody may also provide an opportunity to more accurately investigate a potential pathophysiological role of intra-lobular lymphatics in a variety of lung diseases.
Footnotes
Acknowledgements
We are grateful to Aurélie Chenal and Lydie Germain for their technical help, Prof. P. Callard for his generous and helpful assistance, and Prof. Antoine Martin and Amis du Centre des Tumeurs de Tenon for their constant support.