
Abstract
This study characterized the differentiation of neural stem/precursor cells (NSPCs) isolated from different levels of the spinal cord (cervical vs lumbar cord) and different regions along the neuraxis (brain vs cervical spinal cord) of adult male Wistar enhanced green fluorescent protein rats. The differentiation of cervical spinal cord NSPCs was further examined after variation of time in culture, addition of growth factors, and changes in cell matrix and serum concentration. Brain NSPCs did not differ from cervical cord NSPCs in the percentages of neurons, astrocytes, or oligodendrocytes but produced 26.9% less radial glia. Lumbar cord NSPCs produced 30.8% fewer radial glia and 6.9% more neurons compared with cervical cord NSPCs. Spinal cord NSPC differentiation was amenable to manipulation by growth factors and changes in in vitro conditions. This is the first study to directly compare the effect of growth factors, culturing time, serum concentration, and cell matrix on rat spinal cord NSPCs isolated, propagated, and differentiated under identical conditions.
Keywords
NSPCs have been identified and isolated from the developing embryo and from fetal and adult mammalian brain and spinal cord (Adrian and Walker 1962; Weiss et al. 1996a; Kehl et al. 1997; McKay 1997; Johansson et al. 1999; Namiki and Tator 1999; Gage 2000; Horner et al. 2000; Rietze et al. 2000; Shihabuddin et al. 2000; van der Kooy and Weiss 2000; Tropepe et al. 2001; Yamamoto et al. 2001; Kulbatski et al. 2007). In particular, proliferative cells isolated from the adult rat spinal cord can be propagated in culture and can differentiate into neurons, astrocytes, and oligodendrocytes (Weiss et al. 1996a; Shihabuddin et al. 1997; Kulbatski et al. 2007). The adult mammalian central nervous system (CNS) contains both multipotent and lineage restricted NSPCs (Temple and Alvarez-Buylla 1999), which are regulated by trophic factors (Reynolds and Weiss 1992; Morshead et al. 1994; Craig et al. 1996; Weiss et al. 1996b; Kuhn et al. 1997; Tropepe et al. 1997). Neural restricted precursors (NRPs) and glial restricted precursors (GRPs) have been identified in the brain and spinal cord (Rao 1999; Han et al. 2004; Lepore et al. 2004, 2006; Lepore and Fischer 2005). These cells share the common feature of being more limited in their differentiation potential than multipotent stem cells.
The ideal source of NSPCs for CNS repair is highly debated, with some arguing that all NSPCs are alike and that all can be triggered to differentiate into the appropriate phenotype in response to cues from the local microenvironment. On the other hand, the impetus to transplant region-specific NSPCs is based on studies that suggest that NSPCs have an innate potential to differentiate into particular phenotypes, making them better suited to the environment from which they are derived. Transplantation studies showed that NSPCs react to cues within the host environment and differentiate accordingly (Suhonen et al. 1996; Shihabuddin et al. 2000; Cao et al. 2001). Moreover, cerebral (Parmar et al. 2003) and retinal transplantation (Yang et al. 2002) studies suggest that regional cells are lineage restricted and may respond better to region-specific cues (Gotz 2003).
To date, there has not been a detailed examination of the effect of the regional origin of NSPCs on the differentiation profile of exogenously expanded adult mammalian NSPCs. To understand the post-transplantation differentiation potential of adult mammalian spinal cord NSPCs, we must first understand how they behave in vitro, in response to exogenous expansion. Such an in vitro analysis is important, because NSPCs must first be expanded in vitro before transplantation to generate a sufficient number of cells, and therefore, the pretransplantation differentiation potential of NSPCs must be determined. We chose to focus our efforts on adult rat spinal cord NSPCs specifically, because the adult rat is currently the most commonly used model for studying experimental SCI. This study compares the differentiation profiles of NSPCs isolated from different levels of the spinal cord (cervical vs lumbar spinal cord) and different regions along the neuraxis (brain vs cervical spinal cord). In doing so, we gain an understanding of the region-specific differences of CNS NSPCs. We also assessed the effect on differentiation of platelet-derived growth factor (PDGF), ciliary neurotrophic factor (CNTF), time in culture, serum concentration, and matrix.
In our view, it is desirable to regulate NSPC differentiation to facilitate generation of the selected progeny at specific times after SCI. Growth factors may be necessary to direct NSPCs along the appropriate path, either in vitro before transplantation or as an infusate concomitant with the transplanted cells. We chose to study the in vitro potential of PDGF and CNTF to produce more homogenous populations of glial progeny, based on their known effects on the glial lineage. In particular, we sought to increase the concentration of oligodendrocytic progeny to preserve and/or promote long-term axonal survival and function after SCI.
Time in culture is an important factor in assessing the lineage potential of NSPCs in vitro, because cellular differentiation progresses in a temporal fashion. For example, if the differentiation profile of a population of NSPCs is analyzed prematurely, the results may provide an inaccurate snapshot of the overall percentages of differentiated cells, especially if the overall goal is to transplant NSPCs in long-term studies of spinal cord repair. Therefore, time is another variable that may affect the overall differentiation profile of NSPCs, either by selecting for the survival of a specific population of differentiated progeny or by showing an inherent change in their differentiation potential over time. Thus, in this study, we chose to assess the influence of time in culture on the adult rat spinal cord NSPC differentiation, both in terms of neurosphere age and the total time cells are exposed to the differentiating conditions.
Serum as a cell culture supplement provides a broad spectrum of macromolecules, low molecular weight nutrients, hormones and growth factors, carrier proteins, trace elements, and attachment and spreading factors (Gstraunthaler 2003). Although serum is excluded from the growth media during NSPC propagation, a low dose of serum is added during differentiation, to promote cell binding and subsequent differentiation, as well as to support survival during the period of epidermal growth factor (EGF) / fibroblast growth factor-2 (FGF2) withdrawal. Varying the serum concentration during NSPC differentiation alters the ratio of the differentiated progeny. For example, increased serum concentration increased astrocytic differentiation of human fetal NSPCs and decreased neuronal differentiation (Yin et al. 2004). Thus, serum is another important factor in the in vitro differentiation of NSPCs, and manipulation of serum concentration may provide another strategy for directed NSPC differentiation.
Whittemore et al. (1999) found that cell matrix plays an important role in determining the lineage restriction of adult rat subventricular zone (SVZ) NSPCs. Collagen is produced in abundance by cells that infiltrate the lesioned core of the injured spinal cord, such as meningeal fibroblasts (Berry et al. 1983), endothelial cells (Schwab et al. 2001), and astrocytes (Liesi and Kauppila 2002). This dense collagen matrix forms the backbone of basal lamina sheets during the first week after SCI (Hermanns et al. 2001) and is thought to act as a scaffold for blood vessel, fibroblast, and macrophage activity in the early lesion environment (Maxwell et al. 1984). The effects of collagen on neurons are diverse, depending on the developmental stage and environmental factors, such as the binding and accumulation of inhibitory molecules to the collagen meshwork. Thus, it is important to consider the effects of collagen on the differentiation of NSPCs, because collagen is an important component of the post-SCI environment.
In this study, we hypothesized that the differentiation potential of adult rat spinal cord NSPCs varies between regions along the rostrocaudal neuraxis of the spinal cord as well as between the spinal cord and the SVZ of the brain. We also hypothesized that the following measures change the differentiation potential of adult rat spinal cord NSPCs: exposure to PDGF and CNTF, increased NSPC age in culture, increased differentiation time, 5% FBS, and collagen matrix.
Materials and Methods
Harvesting and Culturing Spheres
We used transgenic hemizygous enhanced green fluorescent protein (EGFP) rats [Wistar-TgN(CAG-GFP)184ys] for all NSPC characterization studies because this is the cell source for in vivo transplantation studies in our laboratory and allows unequivocal identification and tracking of transplanted cells in host tissue (Mothe et al. 2005; Parr et al. 2007). This strain was originally generated by Kobayashi and colleagues (Hakamata et al. 2001) and was obtained from the YS Institute (Utsunomiya, Tochigi, Japan) by Dr. Armand Keating and colleagues (Princess Margaret Hospital/Ontario Cancer Institute; Toronto, ON, Canada) who kindly provided the EGFP Wistar rats for this study from their local breeding stock. All animal procedures were performed in accordance with the National Guide to the Care and Use of Experimental Animals (Canadian Council on Animal Care) and approved protocols from the Animal Care Committee of the Research Institute of the University Health Network, Toronto, ON, Canada.
In this study, 4- to 7-month-old rats were sacrificed, and a dorsal laminectomy was performed to expose the entire length of the spinal cord. Both the cervical (C1-C7) and lumbar (L1-L6) spinal cord was excised and washed in 4C Dulbecco's PBS (DPBS) supplemented with 30% glucose (Sigma-Aldrich; Oakville, ON, Canada). For harvesting of pericentral canal spinal cord NSPCs, the overlying meninges and blood vessels were removed, and the white and some of the gray matter surrounding the central canal were removed. The tissue immediately surrounding the central canal, including the closely apposed ependymal and subependymal regions, as well as surrounding gray matter, was harvested under sterile conditions (Figures 1A and 1B). For harvesting of brain-derived NSPCs from adult male EGFP Wistar rats, the brain was excised and washed in 4C DPBS supplemented with 30% glucose. The overlying meninges and blood vessels were removed. The brain was cut along the midline, and the lateral walls of the lateral ventricles were dissected and collected in DPBS, according to previously described protocols (Reynolds and Weiss 1992; Doetsch et al. 1999). The procedure used for both spinal cord and brain NSPC isolation, including enzymatic dissociation of tissue, media components, and incubation conditions, is described in detail in Kulbatski et al. (2007). Neurospheres were passaged every 5–7 days, according to the methods of Kulbatski et al. (2007). All characterization studies were performed on pericentral canal region cervical or lumbar spinal cord or brain-derived secondary neurospheres (Figures 1C and 1D), unless otherwise indicated. A total of three independent experiments were performed for each experimental group.
Immunocytochemistry
Immunocytochemistry (ICC) was used to assess differentiation in the neurospheres and progeny. The five standard antibodies examined across all groups were glial fibrillary acidic protein (GFAP), radial glia antigen RC1 (RC1), phenobarbital-inducible P450 (RIP), β-III-tubulin (BIIIT), and adenomatous polyposis coli (APC)/CC1. To characterize differentiated progeny, neurospheres were transferred to Matrigel (BD Biosciences; Mississauga, ON, Canada)-coated multi-well culture plates in EGF/FGF2-free media containing 1% FBS. After 7 or 14 days in vitro, ICC was performed according to the methods described in Kulbatski et al. (2007). Specific information about the primary antibodies used and blocking conditions is listed in Table 1. Alexa 568 goat anti-mouse (1:500; Invitrogen, Burlington, ON, Canada) was used as the secondary antibody in all cases. Hoechst nuclear dye (1 μl/4 ml; Sigma-Aldrich) was included at the end of each ICC run to label nuclei. ICC was also performed on undifferentiated neurospheres to assess predifferentiation levels of the various cell markers examined, according to the methods of Kulbatski et al. (2007).
Plating of Neurospheres and Selection of Counting Fields
On average, three whole neurospheres were plated per well, for a total of nine neurospheres in three wells per antibody examined. The total number of cells plated, corresponding to nine spheres in three wells, was estimated based on calculations of neurosphere diameter and volume (Kulbatski et al. 2007). According to these calculations, each secondary neurosphere contained ∼387 cells. Therefore, a total of 3483 cells were plated in three wells for each antibody tested (387 cells per sphere × 9 spheres plated in total). For each antibody, a total of 10 fields of view were selected in the three wells. Also, each experimental manipulation to examine differentiation was repeated three times for a total of n = 30 fields of view.
Images of fields for counting cells were taken at X20 magnification (710 × 530-μm rectangular field) and were selected based on the following criteria: (a) fields containing non-overlapping cells and cells that had already migrated out from the sphere were selected, so that individual cells could be easily identified; (b) fields with either no cells or a small number of cells such as those close to the perimeter of the well were avoided; (c) a total of 10 fields of view were selected in a total of three wells per antibody; and (d) fields with cells that had migrated out from around the entire circumference of the neurospheres were sampled. Cell counts obtained based on these criteria were comparable to cell counts of neurospheres that were dissociated before differentiation. We plated whole spheres for the examination of differentiated progeny to maintain consistency with the in vivo studies (performed by other investigators in our laboratory), in which whole spheres were transplanted rather than dissociated cells because there was better survival in vivo with neurospheres.
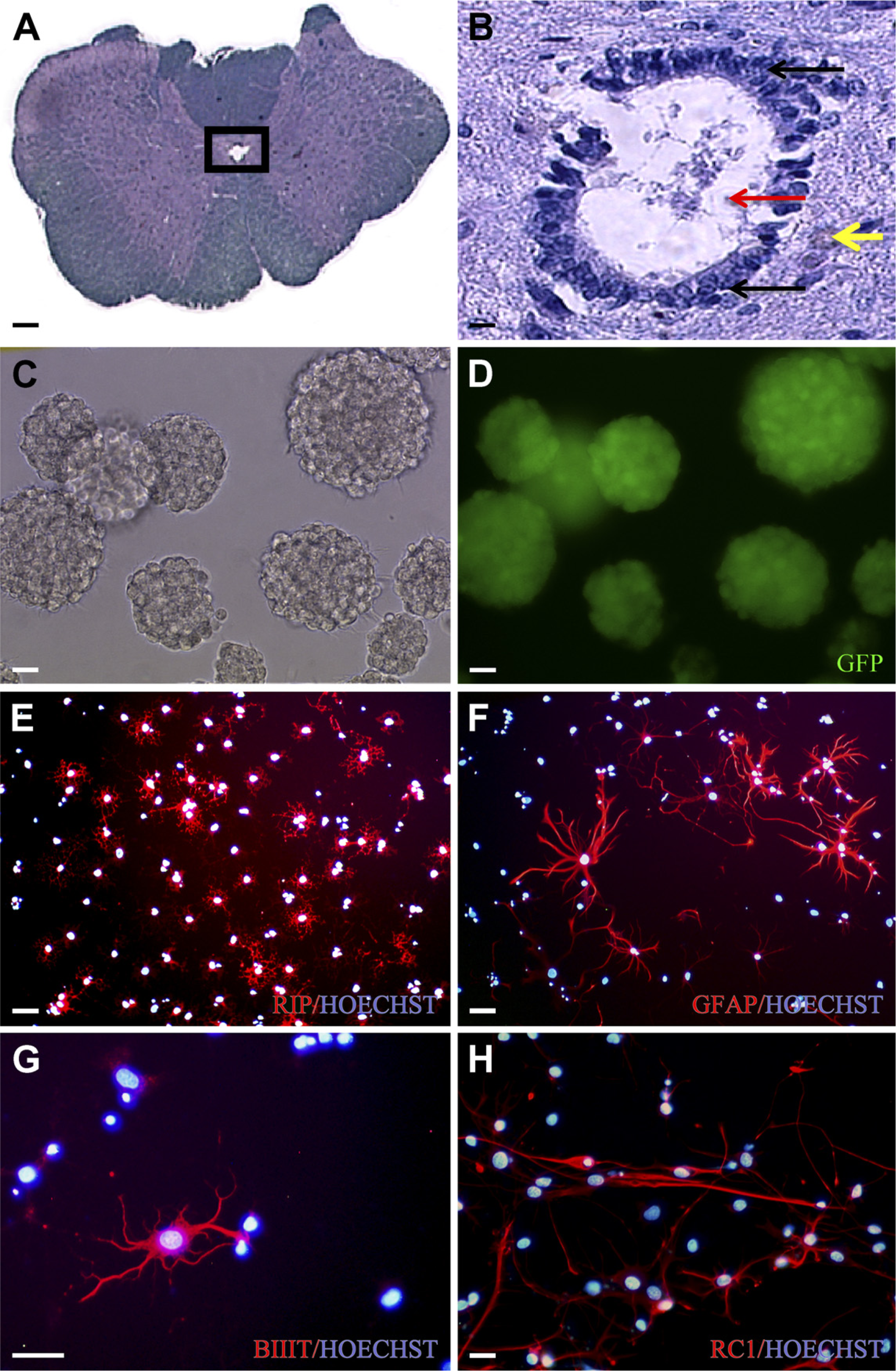
Isolation, propagation, and differentiation of cervical spinal cord neural stem/precursor cells (NSPCs). This figure summarizes, using IHC and immunocytochemical (ICC) images, the isolation, propagation, and differentiation of NSPCs from the pericentral canal cervical spinal cord region of adult enhanced green fluorescent protein (EGFP) rats. (
Detailed comparison of specificity of NSPCs and differentiated progeny ICC markers

This table provides detailed information for the antibodies used to identify NSPCs and their differentiated progeny in this study. The information provided includes antibody specificity, dilutions, and blocking conditions. Also indicated are the coexpression patterns between antibodies and a list of references for each marker. NSPC, neural stem/precursor cell; ICC, immunocytochemical; GFAP, glial fibrillary acidic protein; BIIIT, β-III-tubulin; APC, adenomatous polyposis coli; RIP, phenobarbital-inducible P450; RC1, radial glia antigen; NGS, normal goat serum; MBP, myelin basic protein.
Calculation of Baseline Percent Differentiated Progeny
We designated the pericentral cervical spinal cord neurospheres as the “baseline” control, as a reference for comparison with each experimental manipulation. The mean percent differentiated progeny for the baseline cervical spinal cord was generated from secondary spheres from the pericentral cervical spinal cord. Data from a total of nine independent replications were pooled to generate mean baseline values for each of the five antibodies tested. For each antibody, a total of 10 fields of view were selected in the three wells, for a total of n = 90 fields of view per antibody.
Calculation of Total Number of Hoechst-positive Cells and Percent Differentiated Progeny
The total number of Hoechst-positive cells was calculated for all experiments to determine cell survival and to be used in calculations of percent differentiated progeny. The mean number of Hoechst-positive cells was calculated for each antibody per experimental manipulation (three independent replications per experiment and 10 fields of view per antibody for a total of n = 30 fields of view per antibody). These average numbers of Hoechst-positive cells were used to determine the percentage of differentiated progeny for each antibody in an experiment. To determine the percentage of differentiated progeny, the number of immunopositive cells for each antibody was counted in 10 fields of differentiated cells and added to obtain the total number per antibody. These values were taken as a percentage of the mean total number of Hoechst-positive cells, which was calculated for each antibody in an experiment, as described above.
To compare cell survival between different experimental manipulations, the mean pooled total number of Hoechst-positive cells was calculated by adding the total number of Hoechst-positive cells in 10 fields of view for each of the five antibodies studied and calculating the mean of all five antibodies to obtain a single value. Because each experiment was repeated three times, an average of the total number of Hoechst-positive cells for three independent replications was calculated using 15 Hoechst-positive total cell counts per experiment (n = 30 fields of view per antibody and n = 150 fields of view in total for each experiment). For the baseline group, the mean total number of Hoechst-positive cells was calculated using 45 values generated by nine independent replications for five antibodies (n = 450 fields of view in total).
Image Capture and Statistical Analyses
Fluorescent cells were examined using a Nikon Eclipse TE 300 microscope (Nikon Canada; Mississauga, ON, Canada), and images were captured with a CCD camera and Bioquant Imaging Software (R&M Biometrics; Nashville, TN). Data were collected and analyzed using SigmaStat version 3.1 software (San Jose, CA). Data are presented as mean ± SEM. Statistical differences between groups were evaluated with one-way ANOVA or Student's t-test and multiple comparison procedures with the Bonferroni test. In all analyses, p<0.05 was considered statistically significant.
Effect of PDGF and CNTF on Neural Stem/Precursor Cell Differentiation When Added at the Onset of Culturing
PDGF was added at a concentration of 20 ng/ml (8 × 10−10 M) to NSPCs at the time of harvesting and at each subsequent passage (Bogler et al. 1990; Wolswijk and Noble 1992; Fressinaud 2005). Secondary neurospheres were plated on Matrigel-coated plates for 1 week to differentiate in the absence of EGF/FGF2 and in the presence of 1% FBS. An additional 20 ng/ml PDGF was added at the time of plating. In a separate study, CNTF was added at a concentration of 10 ng/ml (4.4 × 10−10 M) to NSPCs at the time of harvesting and at each subsequent passage (Rajan and McKay 1998; Monville et al. 2001). An additional 10 ng/ml CNTF was added at the time of secondary sphere plating. Neither PDGF nor CNTF was added to the control group. ICC was performed, and the percentage of differentiated progeny and the total number of Hoechst-positive cells were quantitated as described above.
Effect of PDGF and CNTF on Neural Stem/Precursor Cell Differentiation When Added at the Onset of Differentiation
In contrast to the preferential selection studies with PDGF and CNTF described above, preferential differentiation with PDGF and CNTF was achieved by adding the growth factors only at the time of differentiation. Secondary neurospheres were differentiated in the presence of either 20 ng/ml PDGF (8 × 10−10 M) or 40 ng/ml PDGF (1.6 × 10−9 M). In a separate study, CNTF was added a concentration of either 10 (4.4 × 10−10) or 20 ng/ml (8.8 × 10−10 M). Neither PDGF nor CNTF was added to the control group. ICC was performed, and the percentage of differentiated progeny and the total number of Hoechst-positive cells were quantitated as described above.
Effect of Varying Serum Concentration and Cell Matrix on Neural Stem/Precursor Cell Differentiation
Spheres were differentiated in the presence of either 1% or 5% FBS in the absence of EGF/FGF2 on either Matrigel or 30% rat-tail collagen (type I collagen) substrates. A stock solution of rat tail collagen gel was produced by removing tendons from frozen rat tails and solubilizing in acetic acid for 48 hr (Hawrot and Patterson 1979). In total, four groups were studied for the affects of varying serum concentration and substrate on NSPC differentiation: Group A, Matrigel + 1% FBS; Group B, Matrigel + 5% FBS; Group C, collagen + 1% FBS; Group D: collagen + 5% FBS. Spheres were differentiated for 1 week, and ICC was performed to identify neurons, astrocytes, and oligodendrocytes, as described above. The percentage of differentiated progeny and the total number of Hoechst-positive cells were quantitated as described above and compared across each of the four groups.
Effect of Time in Culture on Neural Stem/Precursor Cell Differentiation
Spheres were differentiated in the presence of 1% FBS and in the absence of EGF/FGF2 on Matrigel. ICC was performed at day 7 in vitro on primary, secondary, and tertiary spheres. Neurospheres were first identified at 14 days after isolation (primary spheres) and passaged every 7 days thereafter to yield secondary and tertiary spheres. Table 2 summarizes the experimental outline for examining the influence of time in culture on the differentiation pattern, explaining the total days in culture for each group.
Results
Lumbar vs Cervical Spinal Cord Neural Stem/Precursors
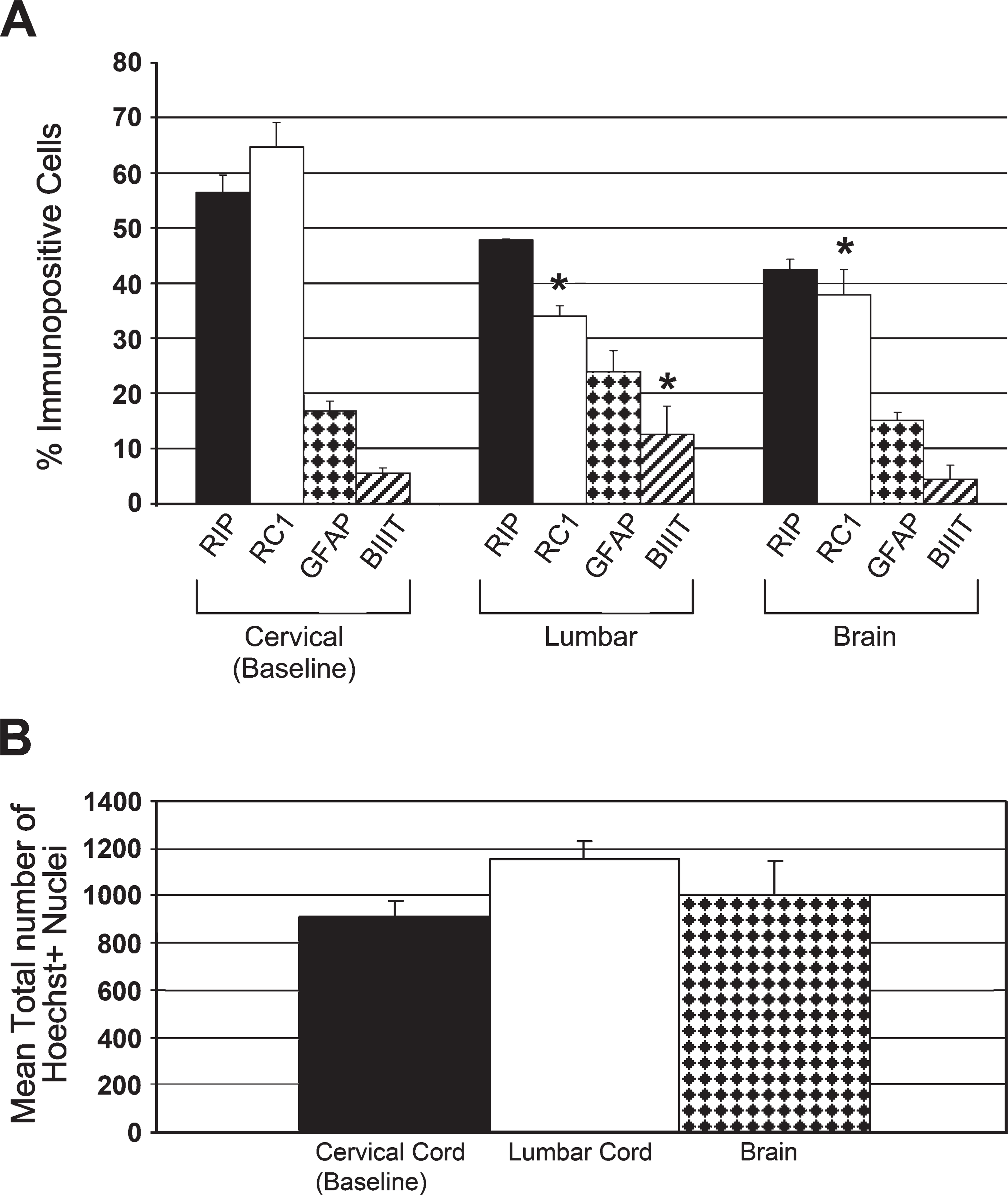
Lumbar and cervical spinal cord formed neurospheres with very similar proportions of progeny (Figure 2A). The majority of lumbar spinal cord NPSCs differentiated into oligodendrocytes (47.9%) and radial glia (RGs) (34.0%). There were relatively fewer astrocytes (23.9%) and neurons (12.5%) from lumbar spinal cord—derived NSPCs compared with the other cell types. Lumbar spinal cord—derived NSPCs differentiated into a significantly higher percentage of neurons than cervical spinal cord—derived NSPCs (p<0.001). As well, there was a significantly lower percentage of RGs from the lumbar spinal cord NSPCs compared with cervical spinal cord—derived NSPCs (p = 0.039). There was no significant difference in the total number of Hoechst-positive cells from differentiated lumbar spinal cord—derived spheres (p>0.05; Figure 2B).
Influence of time in culture on differentiation pattern of adult rat cervical pericentral canal spinal cord NSPCs: experimental outline
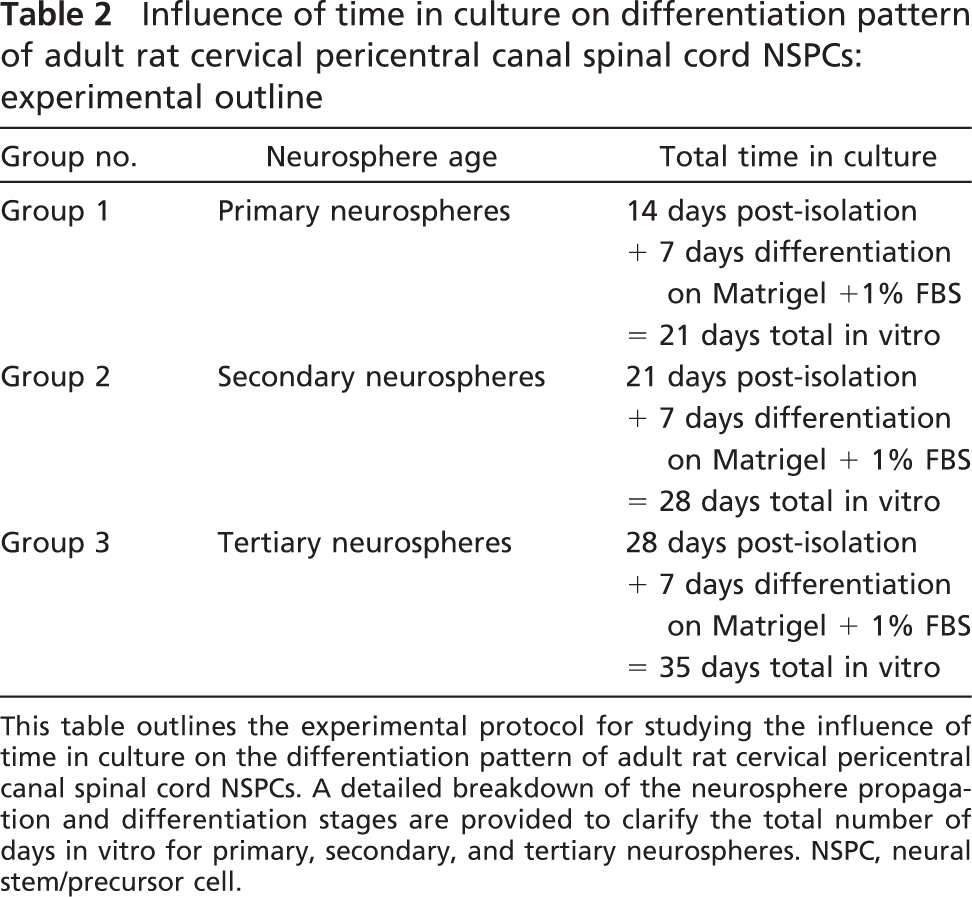
This table outlines the experimental protocol for studying the influence of time in culture on the differentiation pattern of adult rat cervical pericentral canal spinal cord NSPCs. A detailed breakdown of the neurosphere propagation and differentiation stages are provided to clarify the total number of days in vitro for primary, secondary, and tertiary neurospheres. NSPC, neural stem/precursor cell.
Brain vs Cervical Spinal Cord Neural Stem/Precursors
The SVZ region of the brain formed neurospheres that differentiated into the same proportions of progeny as neurospheres from the cervical spinal cord (Figure 2A). The majority of brain-derived NSPCs differentiated into oligodendrocytes (42.5%) and RGs (37.9%), and there were relatively fewer astrocytes (15.0%) and neurons (4.4%). There were no significant differences (p>0.05) between brain- and cervical spinal cord—derived NSPCs in percentages of astrocytes, oligodendrocytes, or neurons. In contrast, brain-derived NSPCs produced significantly fewer RGs compared with the cervical spinal cord NSPCs (p = 0.008). There was no significant difference in the total number of Hoechst-positive cells from differentiated brain- vs cervical cord—derived spheres (p>0.05; Figure 2B).
Effect of Total Differentiation Time on Neural Stem/Precursor Cell Differentiation
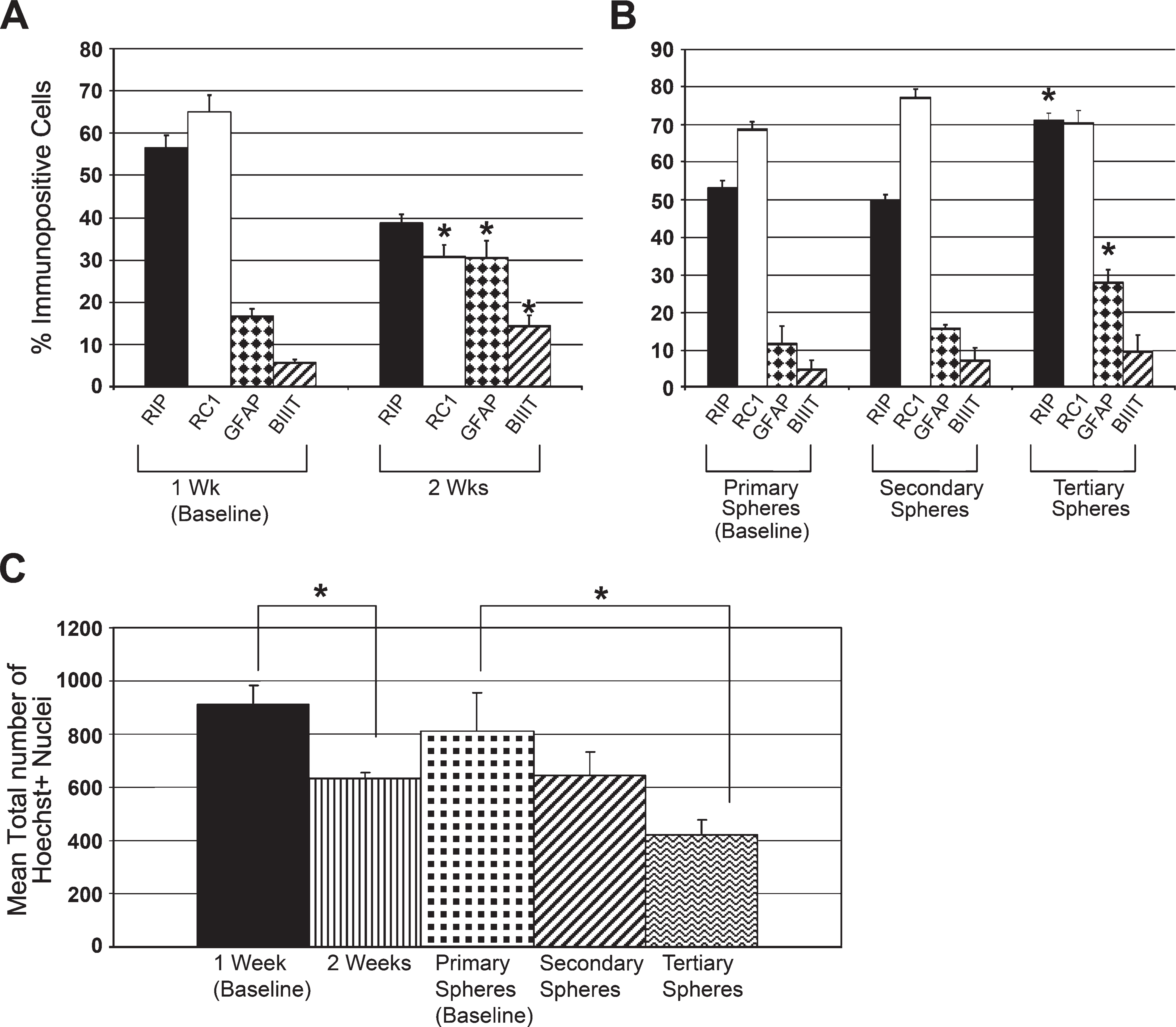
The cervical cord is considered the baseline or reference group. ICC images of the differentiated progeny for cervical spinal cord NSPCs are shown in Figure 1. The differentiation capacity of these NSPCs was assessed at 1 and 2 weeks after plating on Matrigel. It is noteworthy that the percentages of differentiated progeny do not add up to 100%, most likely because of coexpression between antibodies, as summarized in Table 1 and discussed further in the Discussion section. By 2 weeks postdifferentiation, there was a significant increase in GFAP and BIIIT positivity (p = 0.026 and p<0.001, respectively) and a significant decrease in the percentage of RC1-positive cells (p = 0.002). However, there was no significant difference in the number of RIP-positive oligodendrocytes between 1 and 2 weeks (Figure 3A). The differentiated astrocyte progeny consisted of both type 1 and type 2 astrocytes, based on their fibrous or protoplasmic morphology, respectively. There was a significant decrease in the total number of Hoechst-positive cells at 2 weeks (p = 0.018; Figure 3C).
Effect of Time in Culture on Neural Stem/Precursor Cell Differentiation
There was a significant increase in the percentage of GFAP-positive cells between primary and tertiary differentiated neurospheres (p<0.001) and between secondary and tertiary differentiated neurospheres (p = 0.005). There was also a significant increase in the percentage of RIP-positive cells between primary and tertiary differentiated neurospheres (p<0.001) and between secondary and tertiary differentiated neurospheres (p<0.001). The percentages of GFAP- and RIP-positive cells did not change significantly between primary and secondary spheres. There was no interpassage variability in the differentiation patterns of BIIIT-, nestin-, and RC1-positive cells. Overall, there was a doubling of GFAP-positive cells and a 20% increase in the percentage of oligodendrocytes. These data are summarized in Figure 3B. There was a significant decrease in the total number of Hoechst-positive cells between primary and tertiary spheres (p = 0.013; Figure 3C).
Effect of PDGF and CNTF on Neural Stem/Precursor Cell Differentiation When Added at the Onset of Culturing
When 20 ng/ml PDGF was added at the onset of culturing (at the time of tissue isolation and continuously throughout the proliferation and differentiation stages), there was a significant increase in the percentage of oligodendrocytes (p = 0.036; Figure 4A) but a significant decrease in the total number of Hoechst-positive cells compared with the baseline group (p = 0.023; Figure 4C). In contrast, when 10 ng/ml CNTF was added at the onset of culturing, there was a significant increase in the percentage of oligodendrocytes (p = 0.013; Figure 4B), and the total number of Hoechst-positive cells did not change significantly (p>0.05; Figure 4D).
Effect of PDGF and CNTF on Neural Stem/Precursor Cell Differentiation When Added at the Onset of Differentiation
When 20 ng/ml PDGF was added at the onset of differentiation, there was a significant increase in the percentage of RG (p<0.001), but there was no change in the percentage of neurons, oligodendrocytes, or astrocytes. At 40 ng/ml, PDGF at the onset of differentiation significantly decreased the percentage of neurons (p<0.001) and increased the percentage of nestin-positive cells (p<0.001; Figure 4A). There was a significant increase in the total number of Hoechst-positive cells when PDGF was added at the onset of differentiation at a concentration of 20 (p<0.001) or 40 ng/ml (p = 0.023; Figure 4C).
Effect of central nervous system (CNS) region of NSPC isolation on percentage of differentiated progeny. (
When 10 ng/ml CNTF was added at the onset of differentiation, there was a significant increase in the percentage of astrocytes (p = 0.014) and neurons (p<0.001) compared with the untreated baseline group. CNTF (20 ng/ml) administered at the onset of differentiation caused a significant increase in the percentage of oligodendrocytes (p<0.001) and neurons (p<0.001) compared with the baseline. However, compared with the increase in the percentage of neurons seen at 10 ng/ml CNTF, 20 ng/ml CNTF decreased the percentage of differentiated neurons (p<0.001; Figure 4B). Neither concentration of CNTF significantly altered the total number of Hoechst-positive cells (p>0.05; Figure 4D).
Effect of Serum Concentration and Cell Matrix on Neural Stem/Precursor Cell Differentiation
Increasing serum concentration significantly increased astrocytic differentiation on Matrigel (p = 0.004) and collagen (p = 0.007) and significantly decreased oligodendrocytic differentiation on Matrigel (p<0.001) and collagen (p<0.001). In contrast, there was no significant change in neuronal differentiation with increased serum concentration on either Matrigel (p>0.05) or collagen (p>0.05). Collagen significantly decreased astrocytic differentiation at 1% (p = 0.012) and 5% (p = 0.007) serum concentrations and significantly decreased oligodendrocytic differentiation at 1% (p<0.001) and 5% (p<0.001) serum concentration. In contrast, collagen significantly increased neuronal differentiation at 1% (p = 0.008) and 5% serum concentration (p = 0.001; Figures 5A and 5B). Matrigel with 5% FBS significantly increased the number of Hoechst-positive cells compared with Matrigel with 1% FBS (p = 0.001), and collagen with 5% FBS significantly increased the number of Hoechst-positive cells compared with collagen with 1% FBS (p = 0.001). Collagen with 1% FBS significantly decreased the number of Hoechst-positive cells compared with Matrigel with 1% FBS (p = 0.01), and collagen with 5% FBS significantly decreased the number of Hoechst-positive cells compared with Matrigel with 5% FBS (p = 0.018; Figure 5C).
Effect of total differentiation time or total time in culture on percent differentiated progeny. (
Discussion
Understanding the differentiation potential of adult rat spinal cord NSPCs after in vitro expansion is crucial to the design and implementation of future transplantation studies for SCI treatment. We previously showed that adult rat spinal cord—derived NSPCs have the capacity to differentiate into oligodendrocytes, astrocytes, neurons, and RGs, with the highest percentage of progeny being oligodendrocytes and RGs (Kulbatski et al. 2007). This study examined the effect of the region of the CNS from which the NSPCs were derived and the effect of exogenous manipulations on NSPC differentiation. We found that the percent differentiated neurons, astrocytes, and oligodendrocytes from adult rat pericentral canal spinal cord spheres does not differ significantly from that of rat SVZ brain—derived spheres. We also showed that adult rat spinal cord NSPC differentiation is amenable to manipulation by exogenous factors such as growth factors and by changes in vitro environmental conditions and that neurosphere age influences NSPC survival and differentiation.
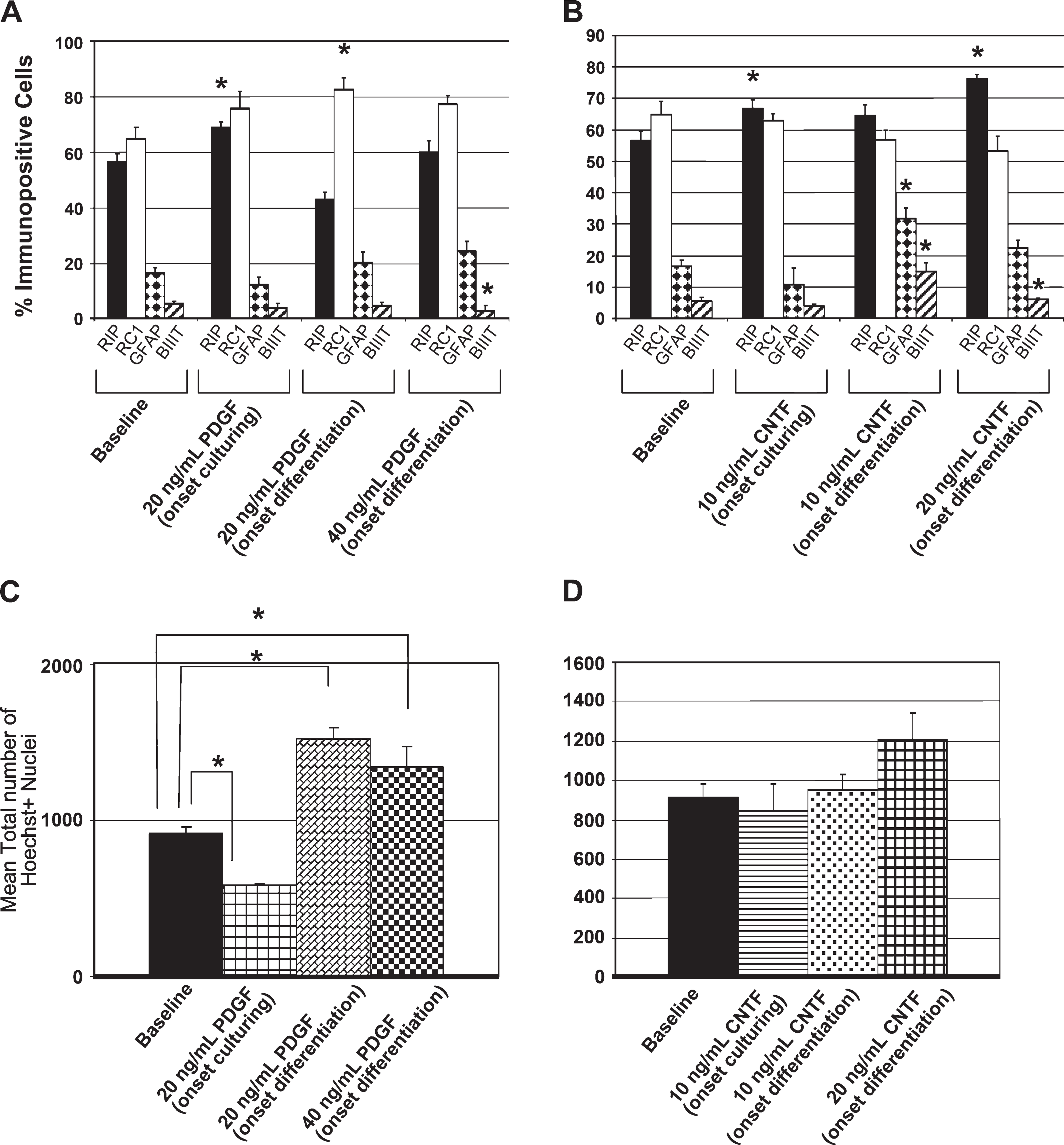
Effect of platelet-derived growth factor (PDGF) or ciliary neurotrophic factor (CNTF) administration on the percentage of differentiated progeny. (
Shihabuddin et al. (1997) were the first to study the differentiation potential of NSPCs harvested from various regions of the adult rat spinal cord and showed that NSPCs from the cervical, thoracic, lumbar, and sacral spinal cord of adult rats varied with respect to the differentiated progeny (Shihabuddin et al. 1997). However, there are major differences between the culture conditions used by Shihabuddin et al. (1997) and the culture conditions we used in this study. They pooled tissue from both the pericentral canal and peripheral white matter and grew the cells as monolayer cultures on poly-
Brain vs Cervical vs Lumbar Spinal Cord—derived NSPCs
Remarkably, we found that adult rat brain—derived NSPCs did not differ from adult rat cervical spinal cord NSPCs in the percentages of differentiated neurons, astrocytes, or oligodendrocytes, although brain NSPCs produced significantly fewer RG cells. This latter difference is not attributable to selective cell death, because the total number of Hoechst-positive cells did not differ between the brain and spinal cord (Figure 2B). In contrast, the lumbar NSPCs produced a significantly higher percentage of neurons than the cervical spinal cord spheres and a significantly lower percentage of RGs. The lack of difference in the total number of Hoechst-positive cells between cervical and lumbar NSPCs (Figure 2B) confirms that these differences are not caused by selective cell survival and death, respectively. The increase in the percentage of differentiated neurons in the lumbar cord may reflect the differences in anatomy and function between the cervical and lumbar cord, specifically the presence in the lumbar cord of networks of interneurons such as the central pattern generator (CPG). The CPG is a network of spinal neurons that produce the rhythmic motor patterns needed for coordinated movements such as walking (Grillner 1975; Grillner and Dubuc 1988; Duysens and Van de Crommert 1998; Van de Crommert et al. 1998). The presence of this specialized network of spinal neurons may influence the local NSPC microenvironment. To our knowledge, there are no studies investigating whether endogenous or transplanted NSPCs from the adult lumbar spinal cord can replace damaged neurons in the CPG.
We expected to see a significant difference in the percentage of astrocytic progeny between the brain and spinal cord NSPCs, based on previous studies that showed that NSPCs from the adult forebrain (Reynolds and Weiss 1992; Gritti et al. 1996) and fetal brain (Lobo et al. 2003) differentiated predominantly into astrocytes. However, aside from the significant decrease in the percentage of RGs, there were no significant changes for astrocytes, oligodendrocytes, or neurons.
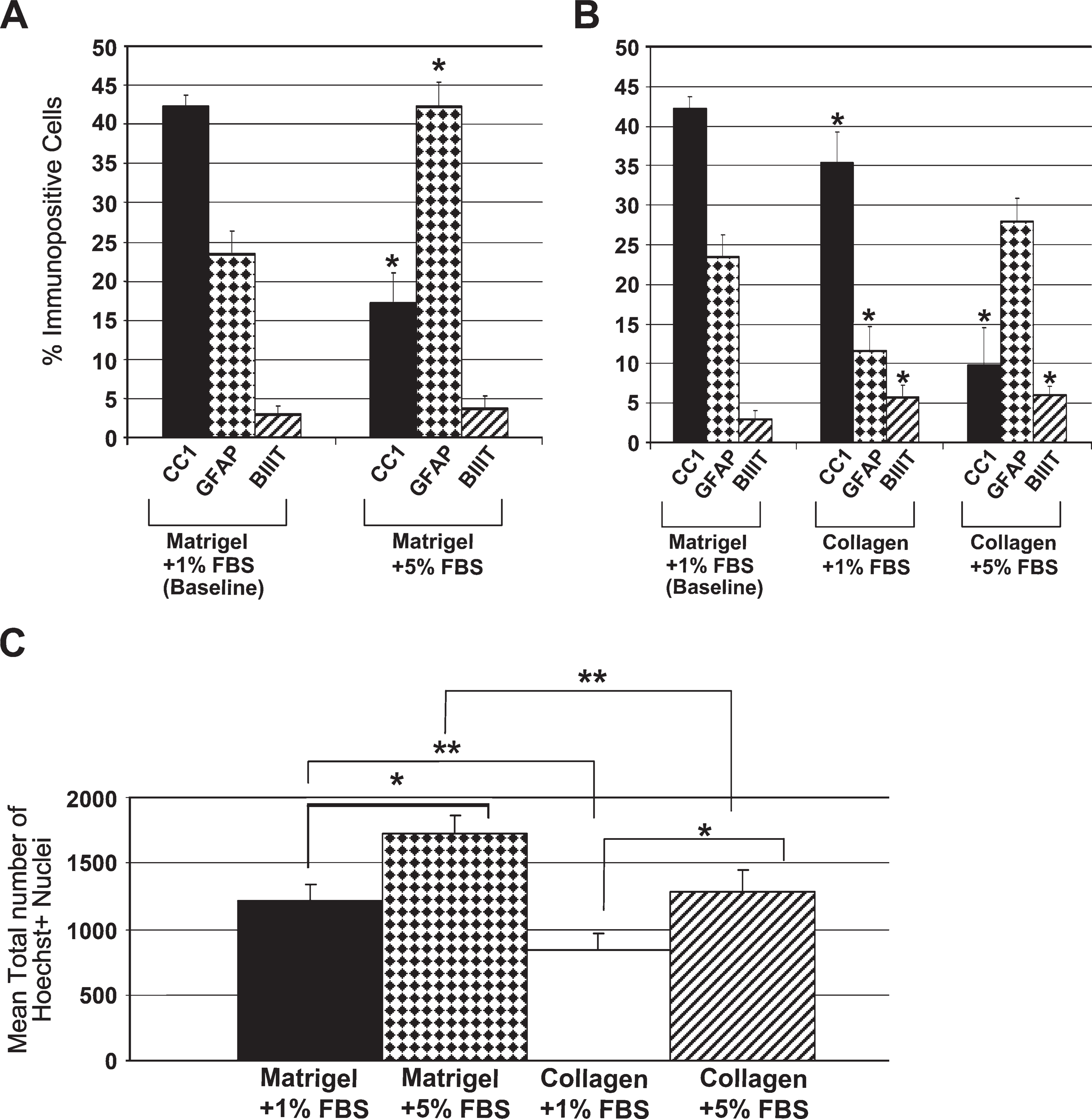
Effect of variations in serum concentration and cell matrix on percentage of differentiated progeny. (
It is known that culture conditions such as cell density can regulate NSPC differentiation (Tsai and McKay 2000). Our high-density culture conditions may have homogenized the differences in fate potential of NSPCs between the brain and spinal cord. For example, neurospheres grown under high-density conditions produce stem cell factor (SCF), a survival and proliferative factor (Erlandsson et al. 2004) involved in the modulation of oligodendrocyte precursors (Ida et al. 1993). Although we did not test for the presence of SCF in our cultures, our in vitro expansion of NSPCs was under high-density conditions and may have involved promotion of oligodendrocyte differentiation by SCF. Future studies should compare the differentiation of brain and spinal cord NSPCs under high- and low-density conditions.
All cell cultures in this study were at high density, because we wished to maintain consistency of the conditions needed to expand NSPCs for subsequent transplantation. We chose to study non—clonally derived populations because of the impracticality of transplanting clonally derived spheres. Large numbers of cells are needed for transplantation, and because the proportion of true stem cells within the culture environment is so low, it is inefficient and impractical to establish a high yield of clonally derived neurospheres for transplantation purposes. Moreover, although the limiting dilution assay and/or clonal assay have been traditionally used to show that a neurosphere originates from a single proliferative NSPC, it has recently been shown that, even under limiting dilution conditions, cell aggregation occurs, producing chimeric neurospheres that cannot be considered clones under the strictest sense of the term (Singec et al. 2006). Furthermore, neurospheres can be formed from a variety of neural stem/precursor “neurosphere-forming” cells. For example, Laywell et al. (1999) showed that a population of multipotent astrocyte stem cells were capable of producing neurosphere clones.
Generation of Enhanced Populations of Differentiated Progeny
PDGF Enhancement of Oligodendrocyte Progeny. PDGF was chosen in this study because it encourages oligodendrocyte progenitor cells to proliferate and differentiate into oligodendrocytes (Bogler et al. 1990; Wolswijk and Noble 1992; Allamargot et al. 2001; Wilson et al. 2003) and increases myelination in spinal cord explants (Park et al. 2001). The methods used for the in vitro administration of PDGF varied considerably in previous studies, and concentrations of PDGF ranged from 1 μg/ml (Wolswijk and Noble 1992) to 20 ng/ml (Chandran et al. 2004). Positive results occurred when PDGF was added to cultures at the onset of culturing and remained present in the medium through multiple passages (Bogler et al. 1990; Wolswijk and Noble 1992; Fressinaud 2005). The timing of growth factor administration is an important consideration, because the same growth factor may vary in the effect it has on NSPC differentiation, depending on when NSPCs are exposed to the growth factor. For example, growth factors added at the time of NSPC isolation and throughout multiple passages vs exposure only during the differentiation phase may select for a particular population of cells that is preferentially responsive to the growth factor. In this study, we compared growth factor administration under both conditions.
PDGF added at the onset of culturing at a concentration of 20 ng/ml significantly increased the percentage of differentiated oligodendrocytes compared with the baseline. In contrast, when added at the onset of differentiation at the same concentration, PDGF did not significantly alter the percentage of differentiated oligodendrocytes but rather significantly increased the percentage of RGs. Even at a higher dose of 40 ng/ml, addition of PDGF at the onset of differentiation did not significantly increase oligodendrocytic differentiation but significantly increased the percentage of nestin-positive cells. Interestingly, 20 ng/ml PDGF added at the onset of culturing significantly decreased cell survival, whereas either 20 or 40 ng/ml PDGF administered at the onset of differentiation significantly increased overall cell survival. Therefore, the increase in the percentage of differentiated oligodendrocytes seen with 20 ng/ml PDGF added at the onset of culturing is likely the result of selective survival of oligodendrocytes in comparison with other cell types. It is unlikely that, under these circumstances, PDGF is promoting oligodendrocyte differentiation or selectively increasing the pool of GRPs or oligodendrocyte precursor cells (OPCs) with each subsequent passage leading up to the differentiation phase. This method of increasing oligodendrocyte differentiation may be a suitable strategy for producing enhanced populations of differentiated oligodendrocytes for transplantation. Additional studies are needed to determine the ideal dose and timing of administration of PDGF. Several other promising strategies for enhancing oligodendrocyte differentiation of NSPCs are currently being studied in other laboratories. For example, the over-expression of Olig2 in NSPCs has been shown to induce oligodendrocyte differentiation in vitro (Copray et al. 2006), and the morphogen sonic hedgehog plays a role in oligodendrocyte specification of NSPCs (Rogister et al. 1999; Murray et al. 2002). Although Keirstead and colleagues recently reported the induction of 98% oligodendrocyte progeny from embryonic stem cells, details were lacking, probably because of patent protection (Keirstead et al. 2005; Nistor et al. 2005)
The enhancement of overall cell survival with PDGF administration at the onset of differentiation may be caused by a trophic effect on differentiating NSPCs or an enhancement in the proliferative capacity of the GRP subpopulation. The increase in nestin-positive cells seen with 40 ng/ml PDGF administered at the onset of differentiation supports this hypothesis of increased cell proliferation. It would be of interest in future studies to investigate this further by testing with proliferation markers such as Ki67 or BrdU to determine the mechanisms of cell survival and/or fate selection. Overall, additional data on the proliferation of NSPCs under the various conditions examined in this study are important for understanding the mechanism of selective lineage enhancement in vitro. Moreover, if any of the in vitro manipulations tested alter the mitogenic capacity of the NSPCs, proliferation assays must also be conducted in vivo after NSPC engraftment to evaluate the potential of the cell to form tumors.
CNTF Enhancement of Astrocyte, Oligodendrocyte, and Neuronal Progeny. CNTF was chosen in this study because of its ability to enhance astrocytic differentiation (Hughes et al. 1988; Lillien et al. 1988; Johe et al. 1996; Bonni et al. 1997; Koblar et al. 1998; Rajan and McKay 1998; Whittemore et al. 1999). Astrocytes regulate the immune response and blood—brain barrier, promote oligodendrocyte remyelination, influence the localized metabolism of glutamate (thereby reducing glutamate toxicity and secondary injury), and provide trophic support for surviving neurons (reviewed in Sofroniew 2000). Astrocytes aid remyelination by releasing leukemia inhibitory factor, which stimulates myelination by oligodendrocytes (Ishibashi et al. 2006). Whittemore and colleagues showed that in the absence of astrocytes, endogenous OPCs failed to remyelinate the injured spinal cord (Talbott et al. 2005). Thus, NSPCs that differentiate into astrocytes may be useful for enhancing recovery after SCI.
We found a significant increase in the percentage of differentiated oligodendrocytes when 10 ng/ml CNTF was added at the onset of culturing. The total number of Hoechst-positive cells remained unchanged between the baseline group and the 10 ng/ml CNTF group. In contrast, 10 ng/ml CNTF added at the onset of differentiation caused a significant increase in the percentage of astrocytes and neurons, whereas 20 ng/ml CNTF at the onset of differentiation significantly increased the percentage of oligodendrocytes and neurons. Neither concentration of CNTF altered the total number of Hoechst-positive cells. Despite the ability of CNTF to promote the differentiation of astrocytes, the relative percentage of GFAP positive cells remained <35%, and in our view, this is not a practical strategy for obtaining enhanced percentages of astrocytic progeny. These results may depend on the particularly high inherent potential of adult rat spinal cord NSPCs to differentiate into oligodendrocytes. Moreover, these cultures exhibit high levels of RC1 expression under normal differentiating conditions. It is known that, depending on the developmental stage, radial glial markers such as RC1 and 3CB2 can label astrocytes as well. It is believed that RGs and astrocytes share a common lineage pathway, and therefore, the minimal response to CNTF seen in these cultures may be attributed to the already high baseline percentages of cells that are positive for RC1. An interesting avenue for future study may be to investigate, using Western blot analysis, whether CNTF can promote maturation of astrocytes from an immature pool of RC1-expressing cells to a pool of cells that express the mature astrocyte marker, GFAP.
It is also important to note that there are two astrocyte lineages, and some studies have reported limited responsiveness of type 2 astrocytes to CNTF in comparison to type 1 astrocytes (Hughes et al. 1988; Levison et al. 1998). The differentiated astrocyte progeny in our adult rat spinal cord NSPC cultures consists of both type 1 and type 2 astrocytes, based on their fibrous or protoplasmic morphology, respectively. Although we did not quantitate the relative numbers of type 1 vs type 2 astrocytes, it is possible that progenitors for type 2 astrocytes outnumber type 1 astrocytes in this culture system, thereby limiting the influence of CNTF.
Downregulation of the CNTF receptor during maturation of astrocytes has been observed, which suggests reduced effects of exogenously administered CNTF on cells kept in culture for longer periods of time (Monville et al. 2001). This hypothesis may explain the reduced effects of CNTF after several days in culture (Hughes et al. 1988; Saadat et al. 1989; Levison et al. 1998) and may thus indicate a need for higher dosages. In addition, several studies reporting optimal doses of CNTF used purified progenitor cultures (Hughes et al. 1988; Levison et al. 1998), which may not be applicable to the dose that should be used in cultures of NSPCs. Additional studies of dose and timing of administration of CNTF may generate higher percentages of astrocytes from adult rat NSPC cultures. Promising strategies currently investigated by other laboratories for enhancing astrocytic progeny include the use of microglia activated interleukin-6 and leukemia inhibitory factor (Nakanishi et al. 2007).
We found that CNTF added at the onset of culturing promoted oligodendrocyte differentiation and when added at the onset of differentiation CNTF promoted oligodendrocyte and neuronal differentiation. Recently, Whittemore and colleagues showed that CNTF promoted cell survival and differentiation into oligodendrocytes in adult rat spinal cord OPCs in vitro (Talbott et al. 2007). CNTF has been reported in vitro to promote differentiation, proliferation, and survival of oligodendrocytes from OPCs (Mayer et al. 1994; Barres et al. 1996). However, these results may be confounded by other factors such as the use of purified O-2A progenitor cultures rather than stem cell cultures or the presence of PDGF in the culture, which as described above stimulates oligodendrocyte differentiation.
CNTF induces differentiation of GABAergic and cholinergic neurons from human and rat tissue in vitro (Saadat et al. 1989; Lawrance et al. 1995; Caldwell et al. 2001), although in some cases, cells in culture were already committed to a neuronal lineage or neurons were present in culture before the addition of CNTF. CNTF also showed a protective effect on neurons in vitro after excitotoxic damage (Semkova et al. 1999). In vivo, there have been controversial results because overexpression of CNTF is correlated with both increased neuronal survival (Finn et al. 1998) and increased motor neuron destruction (Winter et al. 1996). In the latter, however, CNTF in itself was most likely not the underlying cause of neuronal degeneration.
Serum Concentration and Matrix. The use of serum, particularly FBS, is beneficial because it provides a broad spectrum of macromolecules, low molecular weight nutrients, hormones and growth factors, carrier proteins, trace elements, and attachment and spreading factors (Gstraunthaler 2003). Although we excluded FBS from the growth media used to propagate NSPCs, a low dose of serum was added to the differentiating media to promote cell binding and subsequent differentiation, as well as to support survival during the period of EGF/FGF2 withdrawal. We found that increased FBS significantly increased astrocytic differentiation and significantly decreased oligodendrocytic differentiation on either Matrigel or collagen substrates. This supports previous study by Yin et al. (2004), which showed that, as the concentration of serum increased, the differentiation of human fetal NSPCs shifted toward astrocytic progeny. FBS has also been reported to promote differentiation of O-2A progenitors into astrocytes (Raff et al. 1983; Barres et al. 1993). Conversely, studies using culture media without FBS report less astrocytic and more oligodendrocytic differentiation (Raff et al. 1983; Mayer et al. 1994). However, it is important to note that serum is added to the differentiation media during mitogen withdrawal to enhance survival. We found that NSPCs differentiated on either Matrigel plus 5% FBS or collagen plus 5% FBS survived better, based on an increase in the total number of Hoechst-positive cells compared with either matrix in the presence of 1% FBS (Figure 5C). Therefore, it is possible that a higher concentration of FBS caused selective survival of astrocytes. Collagen decreased astrocytic, oligodendrocytic, and neuronal differentiation at 1% serum concentration, which is suggestive of reduced cellular survival overall. Indeed, the total number of surviving cells was lower on collagen compared with Matrigel in the presence of either 1% or 5% FBS (Figure 5C). Given that collagen is an important component of the post-SCI environment, it is important to consider that the presence of collagen in vivo after SCI may decrease survival of transplanted NSPCs in future SCI transplantation studies. Collagen is produced in abundance by cells that infiltrate the injured spinal cord. In fact, the majority of the cells that comprise the post-SCI scar, including infiltrating meningeal fibroblasts (Berry et al. 1983), endothelial cells (Schwab et al. 2001), and astrocytes (Liesi and Kauppila 2002), express collagen. This dense collagen matrix forms the backbone of basal lamina sheets during the first week after SCI (Hermanns et al. 2001) and is thought to act as a scaffold for blood vessel, fibroblast, and macrophage activity in the early lesion environment (Maxwell et al. 1984).
The implantation of synthetic three-dimensional biodegradable tubes into CNS lesions has been studied as a bridging strategy to promote axonal regrowth into the lesion site (Woerly et al. 1998; Plant and Harvey 2000; Novikova et al. 2003; Patist et al. 2004; Nomura et al. 2006a, b, 2008a, b; Zahir et al. 2008). Such techniques can also be used to transplant NSPCs into damaged neural tissue after CNS injury. One of the benefits of using a tube seeded with NSPCs for repairing CNS lesions is that the tube biomaterial, or the cell matrix used to coat the inner lumen of the tube, can provide a platform for the NSPCs, which influences their differentiation. The use of NSPC-seeded tubes can also provide an opportunity to influence the selective differentiation of seeded NSPCs by providing an environment for combination therapy with growth factors (Levenberg et al. 2003). Thus, studying the effect of cell matrix on the differentiation of NSPCs is also highly relevant to future bioengineering strategies involving NSPCs. Our results suggested that Matrigel is a more appropriate matrix for adult rat NSPC differentiation than collagen alone. The major extracellular matrix protein component of Matrigel is laminin, followed by collagen IV, heparin sulfate proteoglycans, and entactin 1, the combination of which may mimic the more complex environment found in tissues.
In this study, the combination of collagen and 5% serum produced the highest number of neuronal progeny. The decrease in oligodendrocytic progeny and increase in neuronal progeny on collagen in the presence of 5% FBS is attributed to selective neuronal survival, given the decrease in overall cell survival seen with collagen. However, the percentage of differentiated neurons was not high enough to constitute an enhanced population. Thus, with respect to serum concentration and matrix, these studies showed differentiation profiles that were not high enough to consider such approaches suitable for producing enhanced populations of cells. Additional strategies may be needed, as outlined in Table 3.
Influence of Time in Culture. We assessed the influence of time in culture in terms of neurosphere age and the total exposure time to the differentiating conditions and found that both time factors influenced differentiation. Increased time in culture during the expansion and propagation phase caused a significant increase in the percentage of GFAP-positive astrocytes and RIP-positive oligodendrocytes. The most marked increase was seen with tertiary neurospheres at day 7 in vitro (35 days total in vitro). The significant decrease in the total number of Hoechst-positive cells in differentiated tertiary spheres suggests that the increase in GFAP- and RIP-positive cells may be a result of selective cell survival. The differentiation profile after 2 weeks differed from that at 1 week. At 2 weeks, there was a significant increase in the percentage of GFAP- and BIIIT-positive cells and a significant decrease in the number of RC1-positive cells (Figure 3A). Interestingly, the sharp decline in RC1 positivity between weeks 1 and 2 coincides with the increase in neuronal and astroglial differentiation. Because RC1 expression decreased over time and because the expression of mature antigens progresses in a temporal fashion, it is reasonable to hypothesize that there is a progression toward increased cell maturation and/or differentiation between weeks 1 and 2 and that the temporal shift toward more restricted phenotypes may occur through the RG cell pool. However, we cannot rule out a selective survival effect in this case, because there was a significant decrease in the total number of Hoechst-positive cells between 1 and 2 weeks (Figure 3C).
Combination of factors to produce enhanced populations of differentiated progeny
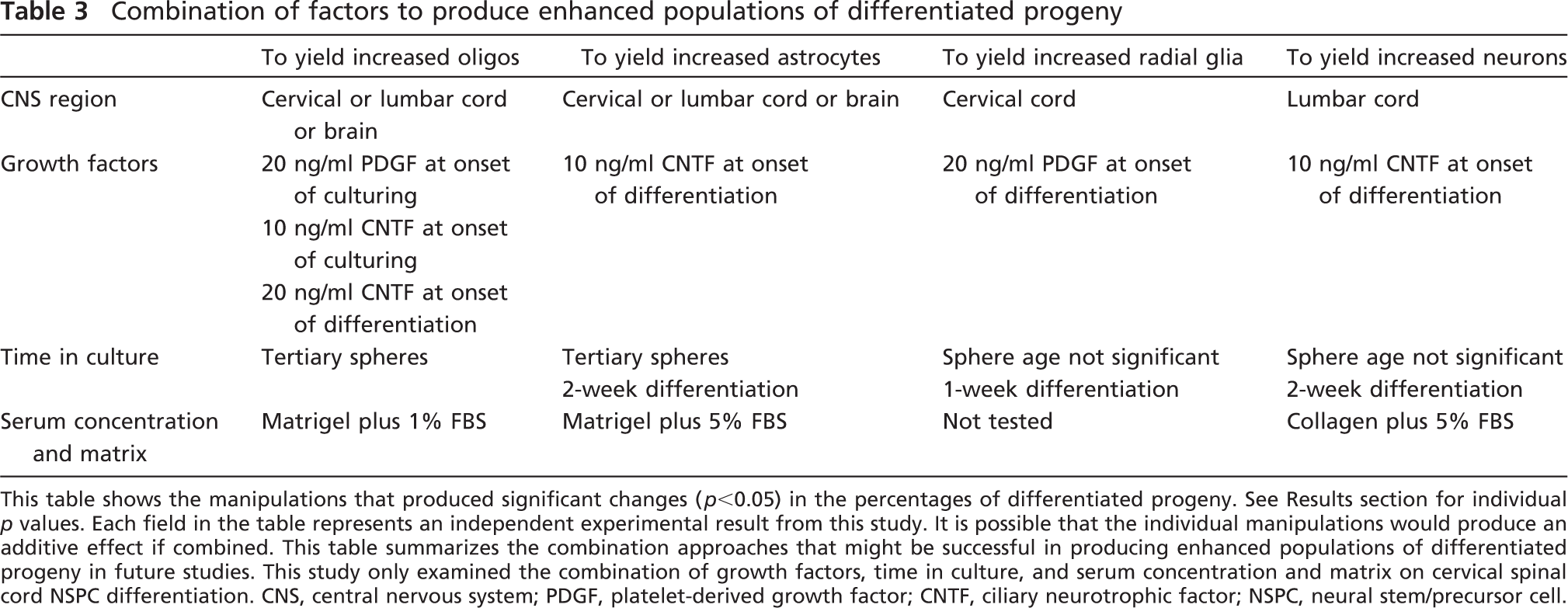
This table shows the manipulations that produced significant changes (p<0.05) in the percentages of differentiated progeny. See Results section for individual p values. Each field in the table represents an independent experimental result from this study. It is possible that the individual manipulations would produce an additive effect if combined. This table summarizes the combination approaches that might be successful in producing enhanced populations of differentiated progeny in future studies. This study only examined the combination of growth factors, time in culture, and serum concentration and matrix on cervical spinal cord NSPC differentiation. CNS, central nervous system; PDGF, platelet-derived growth factor; CNTF, ciliary neurotrophic factor; NSPC, neural stem/precursor cell.
In conclusion, we showed that adult rat spinal cord NSPC differentiation is amenable to manipulation by exogenous factors such as growth factors and by changes in vitro environmental conditions. The differentiation pattern of adult rat pericentral canal cervical spinal cord spheres does not differ significantly from that of rat SVZ brain—derived spheres. Our study emphasizes that the time in culture of neurospheres at the time of transplantation is crucial for optimizing NSPC survival and influencing differentiation. Our results contribute to the basic understanding of adult mammalian spinal cord—derived NSPCs and are useful for the design of future studies in animal models of NSPC transplantation after SCI. Moreover, it shows that CNS NSPCs grown under different culture conditions display differences in their characteristics, and therefore, caution must be taken in making direct comparisons.
From a clinical perspective, adult human spinal cord NSPCs can theoretically be obtained from the spinal cord of organ transplant donors and used in transplantation therapies for SCI patients, avoiding ethical and oncogenic concerns expressed about embryonic stem cell transplantation. Another advantage of adult spinal cord—derived NSPCs is that they more closely resemble the cells they are meant to replace. Our study has generated many different avenues for future study to address the identified limitations. This study was designed with the overall purpose of facilitating transplantation therapies for human SCI, and future in vitro studies are needed to achieve this goal. For example, we identified some combinations of strategies to produce enhanced populations of differentiated phenotypes from NSPCs, as described in Table 3. Moreover, it would be useful to use some of our in vitro manipulations as an adjuvant to in vivo NSPC transplants, in particular the administration of growth factors to guide differentiation. It is noteworthy that our laboratory recently showed that, after transplantation into the injured adult rat spinal cord, the NSPCs derived from the adult rat pericentral cervical spinal cord differentiate into similar percentages of oligodendrocytes, astrocytes, and neurons as found in vitro (Parr et al. 2007). Based on this information, we can presume that these NSPCs retain a similar differentiation potential in vivo. As such, strategies to enhance specific phenotypes in vitro, such as oligodendrocytes using 20 ng/ml CNTF administered at the onset of differentiation, can be applied to guide differentiation in vivo as well. Some of the factors investigated in this study also showed the ability to improve overall cell survival (i.e., 20 ng/ml PDGF administered at the onset of differentiation). Because survival of NSPCs after transplantation is currently a challenge (Parr et al. 2007), the findings presented in this study may be highly applicable to future in vivo studies.
Footnotes
Acknowledgements
Funding for this study was provided by grants from the Ontario Neurotrauma Foundation, Canadian Paraplegic Association (Ontario Branch), Physician's Services Incorporated, International Foundation for Research in Paraplegia, and the Christopher Reeve Paralysis Foundation (to CHT). Personal support was provided by an Ontario Student Opportunity Trust Fund/Vision Sciences Scholarship and Sandra and David Smith Graduate Student Award (to IK). We extend our appreciation to all these organizations for their support.
We thank Xinghua Wang, Huijie Jiang, Dr. Armand Keating, and Dr. Eiji Kobayashi for maintaining a steady supply of Wistar-TgN(CAG-GFP)184ys rats for the studies, Dr. Andrea Mothe and Dr. Cindi Morshead for their constructive advice, Sheer Ramjohn for his assistance in electron microscopy, Alanna Weisman for her help with the data analysis, and Richard Logan, Vikas Kaushal, and Vishanthan Sivagnanam for kindly providing cortical neurons for the co-culture experiments.