
Abstract
Tight junctions (TJs) are an essential structure of fluid-secreting cells, such as those in salivary glands. Three major families of integral membrane proteins have been identified as components of the TJ: claudins, occludin, and junctional adhesion molecules (JAMs), plus the cytosolic protein zonula occludens (ZO). We have been working to develop an orally implantable artificial salivary gland that would be suitable for treating patients lacking salivary parenchymal tissue. To date, little is known about the distribution of TJ proteins in adult human salivary cells and thus what key molecular components might be desirable for the cellular component of an artificial salivary gland device. Therefore, the aim of this study was to determine the distribution of TJ proteins in human salivary glands. Salivary gland samples were obtained from 10 patients. Frozen and formalin-fixed paraffin-embedded sections were stained using IHC methods. Claudin-1 was expressed in ductal, endothelial, and ∼25% of serous cells. Claudins-2, −3, and −4 and JAM-A were expressed in both ductal and acinar cells, whereas claudin-5 was expressed only in endothelial cells. Occludin and ZO-1 were expressed in acinar, ductal, and endothelial cells. These results provide new information on TJ proteins in two major human salivary glands and should serve as a reference for future studies to assess the presence of appropriate TJ proteins in a tissue-engineered human salivary gland.
Keywords
I
We have been working to develop an orally implantable fluid secretory device (an artificial salivary gland) that would be suitable for treating patients lacking salivary parenchymal tissue (Baum et al. 1999; Aframian et al. 2000, 2001, 2002a, b). Our initial choice of an allogeneic human submandibular gland cell line (HSG) was unable to attain a polarized monolayer organization and did not express any tight junction (TJ) proteins or generate a transepithelial electrical resistance (TER) (Aframian et al. 2002b). We re-engineered HSG cells to express individual or a combination of key TJ proteins (claudin-1 or/and claudin-2) by establishing stable clones of transfected cells. Nonetheless, when we examined these transfectants for functional tight junctional activity (TER or fluid barrier), none was observed (Aframian et al. 2002b). The inability of the HSG cell line to form TJs led to the development of a method of culturing and expanding primary human salivary gland (huSG) cells. The cultured huSG cells have been shown to possess tight and intermediate junctions between cells, a transepithelial electrical resistance, and a low paracellular fluid movement, all of which are characteristics of polarized cells (Tran et al. 2005).
Epithelial cells with primary secretory functions, such as in salivary glands, must be polarized to achieve a unidirectional secretion. TJs are the primary structure that forms a barrier between the apical and basal region of the cell. In the apical face of the cell, the transport proteins produce an osmotic concentration gradient, in addition to providing a facilitated water permeability pathway. TJs also promote a “fence” function that maintains the differential composition of the basolateral and apical domains by preventing the free diffusion of lipids and proteins between these compartments (Baum 1993; Mostov et al. 2000). In salivary glands, this gradient separation becomes important for the effective movement of ions, such as sodium and chloride, required for the production of saliva. Water and solutes that will form the exocrine secretion must pass across three different barriers: the vascular endothelium, the glandular interstitium, and the secretory epithelium (Smaje and Henderson 1984). This movement in effect is regulated by tissue-specific TJs, with permissive and barrier mechanisms, to achieve the chemical specificity of saliva (Baron et al. 1999). TJs are built from almost 40 different proteins, including members from multigene families (Morita et al. 1999b). Among these proteins are three types of transmembrane proteins: claudins, occludin, and junctional adhesion molecules (JAMs), as well as cytoplasmic proteins fulfilling roles in scaffolding, cytoskeletal attachment, cell polarity, signaling, and vesicle trafficking. The most important cytoplasmic protein is the peripheral membrane protein, zonula occludens (ZO) (Tsukita et al. 2001). The composition of TJs is quite complex and diverse, apparently much more than the other epithelial junctions: gap, adherens junctions, and desmosomes. These proteins have been comprehensively reviewed by Gonzalez-Mariscal et al. (2003) and Schneeberger and Lynch (2004).
There are few reports on the distribution of TJ proteins in human salivary glands in the literature (Kriegs et al. 2007; Lourenço et al. 2007). Accordingly, in this study, we evaluated the presence and distribution of various TJ proteins in human submandibular and parotid glands to serve as a reference for future studies where it may be necessary to assess the presence of specific TJ proteins in a tissue-engineered human salivary gland.
Materials and Methods
Source of Human Salivary Glands
Portions of human submandibular (
IHC
Frozen tissue slides were fixed in methanol over dry ice for 10 min, followed by two washing steps in PBS for 5 min each. Endogenous peroxidase and biotin activities were blocked with hydrogen peroxide 3% and the Avidin D/Biotin kit (Cat. SP-2001; Vector Laboratories, Burlingame, CA), respectively. All primary antibodies were obtained from Zymed Labs (South San Francisco, CA). We used the following antibodies: rabbit anti-claudin-1 (Cat. 51–9000), claudin-2 (Cat. 51–6100), claudin-3 (Cat. 34–1700), occludin (Cat. 42–2400), JAM-A (Cat. 36–1700) and mouse anti-ZO-1 (Cat. 33–9100), claudin-4 (Cat. 32–9400), and claudin-5 (Cat. 35–2500). In addition, we used rabbit and mouse isotype control antibodies (Cat. 08–6599 and 08–6199). These antibodies are reactive against the respective proteins from human and other species. Secondary broad spectrum antibodies (Cat. 95–9743) were also obtained from Zymed Labs. All primary antibodies used were diluted 1:20 in PBS containing 5% goat serum (Cat. S-1000; Vector Laboratories). The slides were incubated with the primary antibodies for 60 min. A secondary broad spectrum antibody kit was used with adding either 3-amino-9-ethyl-carbazole (AEC; Cat. 00–1111, Zymed Labs) or diaminobenzidine (DAB; Cat. 00–2014, Zymed Labs) chromogen to observe the reaction color in either red or brown, respectively. The slides were counterstained with Mayer's hematoxylin (Cat. 245–653; Fisher Scientific, Kalamazoo, MI). For formalin-fixed paraffin-embedded (FFPE) tissue slides, we used the same steps as described above, in addition to a boiling step (15 min) using an EDTA solution (pH 8.0, Cat. 00–5500; Zymed Labs) for heat-induced epitope retrieval. Primary antibodies were incubated overnight with the FFPE slides at 4C.
Results
We examined slides of glands from both male (
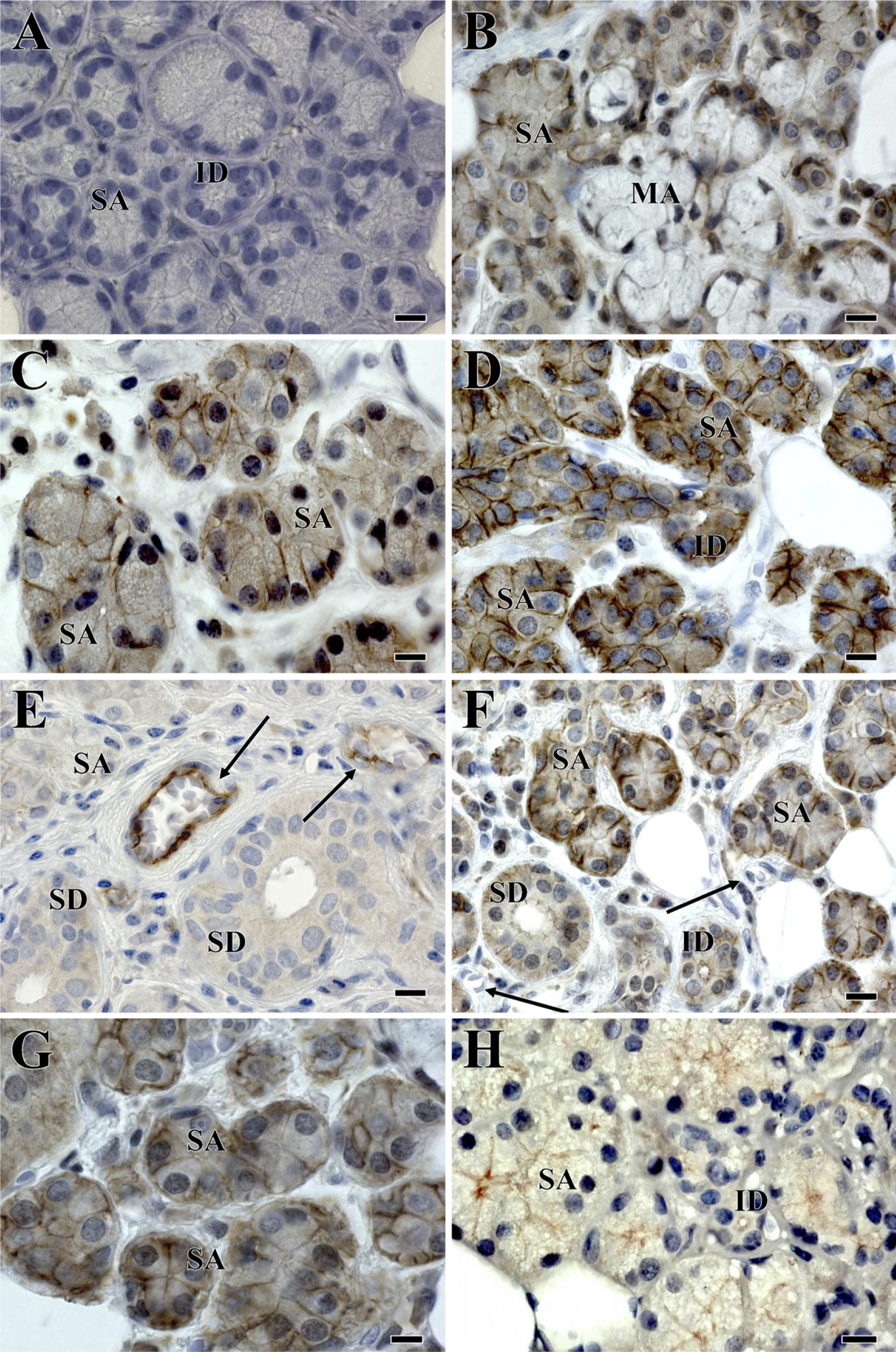
Light micrographs of IHC staining of human salivary gland sections.
Discussion
In this study, we evaluated the presence of multiple TJ proteins (claudins-1, −2, −3, −4, and −5, JAM-A, occludin, and ZO-1) in human parotid and submandibular salivary glands. The summary of our results, shown in Table 1, provides the most detailed examination of TJ proteins in human major salivary glands reported to date (Kriegs et al. 2007; Lourenço et al. 2007) and should be a useful guide for evaluating TJ proteins present in the cellular component of any artificial salivary gland device. However, we did not examine all possible TJ proteins in these tissues for practical reasons, but rather, selected the most common TJ proteins for which antibodies were readily available.
TJs, together with adherens junctions and desmosomes, form the apical junctional complex in epithelial and endothelial cellular sheets. Adherens junctions and desmosomes are responsible for the mechanical adhesion between adjacent cells, whereas TJs are essential for the tight sealing of the cellular sheets, thus controlling paracellular ion flux and therefore maintaining tissue homeostasis (Tsukita et al. 2001). TJs also play a crucial role in the maintenance of cell polarity by forming a fence that prevents lateral diffusion of membrane proteins and lipids, thereby maintaining the differential composition of the apical and basolateral domains. Finally, because of the ability of TJ proteins to recruit signaling proteins (Mitic and Anderson 1998), TJs have also been hypothesized to be involved in the regulation of proliferation, differentiation, and other cellular functions. Given these cellular biological roles, the presence and function of TJ proteins in the cellular component of an artificial salivary gland is critically important.
Thus far, 24 members of the claudin family have been identified in mouse and human (Morita et al. 1999b; Tsukita et al. 1999). Recent findings indicate the importance of claudin-1 in essential physiological functions and in the formation of barriers. For example, claudin-1 knockout mice survive only for 1 day after birth and present with altered epidermal barrier properties (Furuse et al. 2002). We observed claudin-1 in salivary ductal, endothelial, and in ∼25% of serous acinar cells. Why claudin-1 was apparently not detected in the remaining serous acinar or in mucous acinar cells is not clear. It may reflect differences in the functional state of the cells at the time of surgery or represent an example of different acinar cell subpopulations. Lourenço et al. (2007) reported that claudin-1 was expressed only in ductal cells of adult human minor salivary glands. The presence of claudin-1 protein may suggest that it functions as a barrier to prevent the loss of salivary tonicity through the walls of the salivary system.
We found a ubiquitous distribution of claudin-2 in salivary gland sections. The IHC staining was stronger in the acinar cells (both serous and mucous) than ductal cells. Claudin-2 has been reported to weaken the tightness of TJs compared with the proposed barrier function of the claudin-1 protein (Stevenson et al. 1988). For example, Madin-Darby Canine Kidney (MDCK) type I and II cells have similar numbers of TJs expressed; however, MDCK I cells have a tighter barrier, reflected by a higher TER, compared with MDCK II cells (Stevenson et al. 1988). MDCK I cells mainly express claudin-1, whereas MDCK II cells express claudin-2 in addition. Because epithelial cells of salivary glands are considered to be relatively leaky exocrine tissues (Augustus et al. 1978), our observation seems to be in agreement with this organ's function. Results from human minor salivary glands indicate the absence of claudin-2 in both ductal and acinar cells (Lourenço et al. 2007).
In our study, claudin-3 and claudin-4 were present in both acinar and ductal cells. Similar results were reported from studies in rats (Peppi and Ghabriel 2004) and in human minor salivary glands (Lourenço et al. 2007). Claudin-3 is not involved directly in barrier properties of epithelial cells (Furuse et al. 2001; Amasheh et al. 2002), although the presence of claudin-4 in TJs is known to decrease sodium permeability, causing a decreased paracellular conductance (Van Itallie et al. 2001). When the claudin-4 in MDCK I cells was removed, there was a decrease in the number of TJ strands along with an increase in cellular permeability (Sonoda et al. 1999). Claudin-5 seems to have an important role in the vascular permeability of endothelial cells (Morita et al. 1999a). In this study, the detection of claudin-5 only in endothelial cells of human salivary glands is consistent with the reported specificity of this molecule. However, Lourenço et al. (2007) reported the expression of claudin-5 in both ductal and acinar cells. This difference might be a reflection of technical differences from our study. Herein, we used a claudin-5 monoclonal antibody, whereas in the study of Lourenço et al. (2007), a claudin-5 polyclonal antibody was used.
Summary of tight junction proteins present in adult human submandibular and parotid salivary glands
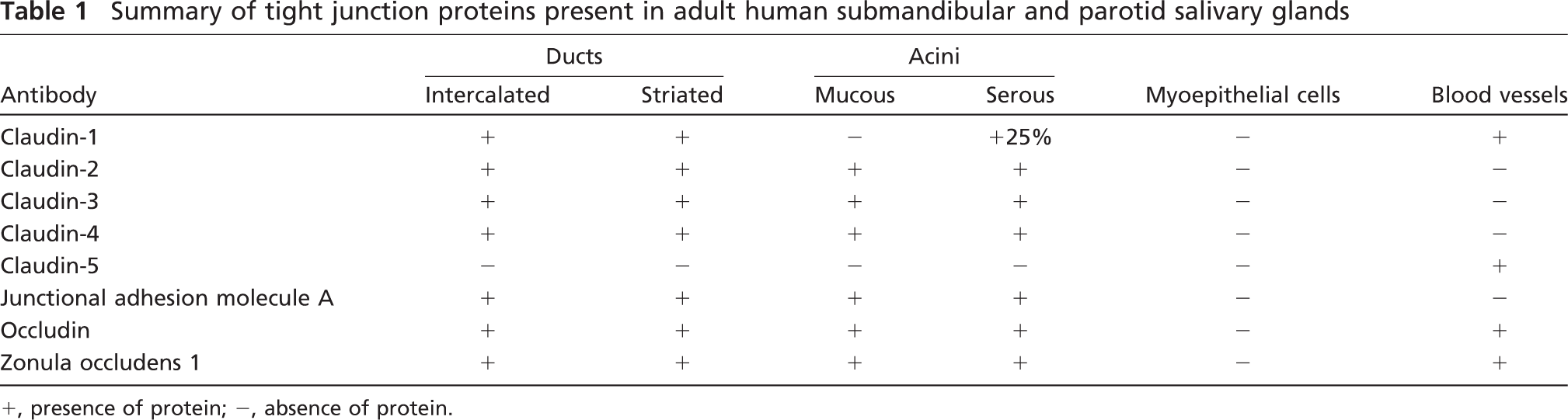
+, presence of protein; −, absence of protein.
The JAMs are four glycosylated transmembrane proteins (Shin et al. 2006), but in this study, we examined the expression of JAM-A (also known as JAM-1) protein only. JAMs were recently shown to bind directly to ZO-1 (Bazzoni et al. 2000; Ebnet et al. 2000), although the full role of JAM-related proteins still needs to be clarified. Herein, JAM-A protein was detected in ductal and acinar epithelial cells.
Occludin, a phosphoprotein, was the first transmembrane protein to be identified in TJs (Furuse et al. 1993). We localized occludin in ductal, acinar, and endothelial cells of human salivary glands. These results were consistent with the results reported by Kriegs et al. (2007) with regard to acinar and ductal cells but not for endothelial cells. The presence of occludin may provide the TJs with the ability to form seals between cells, preventing the leakage of saliva. Similarly, the expression of occludin has been reported in ductal and acinar cells of rat salivary glands (Peppi and Ghabriel 2004). Occludin interacts directly with ZO proteins (ZO-1, −2, and −3) (Gonzalez-Mariscal et al. 2000). Also, occludin interacts with the actin cytoskele-ton and JAMs proteins indirectly through ZO proteins (Mandell and Parkos 2005). Tsukita et al. (2001) indicated that occludin might contribute to the electrical barrier function of TJs and possibly to the formation of aqueous pores within TJ strands; however, TJs can be present independent of occludin.
There are three isoforms of ZO proteins: ZO-1, −2, and −3. Because ZO proteins bind to actin, they act as scaffolds to link other TJ proteins and cytoskeleton (Shin and Margolis 2006). We only evaluated the distribution of ZO-1 in human salivary gland tissue. Unlike other TJ proteins, ZO-1 is not a transmembrane protein; rather, it is a large cytosolic phosphoprotein (Stevenson et al. 1986), which is critically important for claudin polymerization (Umeda et al. 2006). ZO-1 was found in endothelial as well as ductal and acinar cells of human salivary glands. Gresz et al. (2004) and Kriegs et al. (2007) reported that ZO-1 seemed to be restricted to the apical lateral region of acinar cells, presenting spider-like distribution, similar to that of aquaporin-5, in rat salivary glands, and our findings are generally comparable to their results.
In summary, we presented new data on the localization of several TJ proteins (claudin-1 to −5 and JAM-A) in normal human submandibular and parotid glands, as well as confirmed the presence of occludin and ZO-1 in these tissues (Kriegs et al. 2007). In showing the presence and localization of these proteins, we expect that our findings will allow a more critical assessment of TJ proteins in cultured primary salivary gland cells for use in an envisioned tissue-engineered salivary gland. Further studies are needed to find the exact roles and interactions of TJ proteins essential for physiological salivary gland functions.
Footnotes
Acknowledgements
This study was supported by Canada Research Chair and Canadian Institutes of Health Research, Egyptian Cultural and Educational Department (to OMM), and the Division of Intramural Research, National Institute of Dental and Craniofacial Research.
The authors thank Dr. Christopher Moskaluk for the histopathological analysis of salivary glands obtained from Cooperative Human Tissue Network and Dr. Younan Liu for technical assistance.