
Abstract
Human aldo-keto reductase (AKR) 1C3 (type 2 3α-hydroxysteroid dehydrogenase/type 5 17β-hydroxysteroid dehydrogenase) catalyzes androgen, estrogen, and prostaglandin metabolism. AKR1C3 is therefore implicated in regulating ligand access to the androgen receptor, estrogen receptor, and peroxisome proliferator activating receptor γ in hormone target tissues. Recent reports on close relationships between ARK1C3 and various cancers including breast and prostate cancers implicate the involvement of AKR1C3 in cancer development or progression. We previously described the characterization of an isoform-specific monoclonal antibody against AKR1C3 that does not cross-react with related, >86% sequence identity, human AKR1C1, AKR1C2, or AKR1C4, human aldehyde reductase AKR1A1, or rat 3α-hydroxysteroid dehydrogenase (AKR1C9). In this study, a clone of murine monoclonal antibody raised against AKR1C3 was identified and characterized for its recognition of rat homolog. Tissue distribution of human AKR1C3 and its rat homolog in adult genitourinary systems including kidney, bladder, prostate, and testis was studied by IHC. A strong immunoreactivity was detected not only in classically hormone-associated tissues such as prostate and testis but also in non—hormone-associated tissues such as kidney and bladder in humans and rats. The distribution of these two enzymes was comparable but not identical between the two species. These features warrant future studies of AKR1C3 in both hormone- and non—hormone-associated tissues and identification of the rodent homolog for establishing animal models.
T
Originally cloned from human prostate (Lin et al. 1997) and placental cDNA libraries (Dufort et al. 1999), AKR1C3 catalyzes androgen, estrogen, PG, and xenobiotics metabolism. The relatively high 17β-HSD activity of this enzyme reduces δ4-androstene-3, 17-dione (a weak androgen) to yield testosterone (a potent androgen) and reduces estrone (a weak estrogen) to yield 17β-estradiol (a potent estrogen) (Penning et al. 2000). Using its 3α-HSD activity, AKR1C3 reduces 5α-dihydrotesterone (5α-DHT, a potent androgen) to 5α-androstane-3α,17β-diol (3α-diol, a weak androgen) (Lin et al. 1997). AKR1C3 also possesses PG 11-ketoreductase activity to reduce PGD2 to 9α,11β-PGF2α (Matsuura et al. 1998). As a result, AKR1C3 may deprive PGJ2, a ligand for the peroxisome proliferator activating receptor γ (PPARγ), and lead to suppressed cell differentiation (Desmond et al. 2003). AKR1C3 is therefore capable of governing ligand access to various nuclear receptors (Penning et al. 2006). Deregulated expression of AKR1C3 has been shown in multiple types of cancers, including myelodysplastic syndrome (MDS, refractory anemia) (Mahadevan et al. 2006), breast cancer (Lewis et al. 2004), endometrial cancer (Rizner et al. 2006), lung cancer (Lan et al. 2004), and localized (Nakamura et al. 2005; Fung et al. 2006) and advanced (Stanbrough et al. 2006; Wako et al. 2008) prostate cancer. With increasing implications of AKR1C3 in pathological development of human cancers, an animal model is urgently needed to study roles of AKR1C3 in disease development. In addition to human isoforms, enzyme activities and biological functions of AKR1C isozymes in rodent models are largely unclear. Rat AKR1C9 has been thoroughly studied for its hormone metabolism (Penning et al. 1984; Pawlowski et al. 1991). However, like their human counterparts, rats also express multiple members of the AKR1C family including AKR1C8, AKR1C15, AKR1C16, AKR1C17, AKR1C18, and AKR1C24 (www.med.upenn.eud/akr).
We described an AKR1C3 specific monoclonal antibody (NP6A6.G6) that recognizes AKR1C3 but not other related enzymes such as AKR1C1, AKR1C2, AKR1C4, or AKR1C9 (Lin et al. 2004). The objectives of this study are to understand the distribution of AKR1C3 in the human genitourinary system and to identify the murine homolog for an animal model. Using AKR1C3-specific monoclonal antibody and another clone of monoclonal antibody raised against AKR1C3 that recognizes rat homolog using IHC staining, our results showed comparable but not identical cellular distribution of AKR1C3 between human and rat genitourinary tissues. In addition to its expected distribution in hormone target tissues, we also showed that AKR1C3, and its rat homolog, are present in cells that are not classically defined as hormone-dependent tissues such as the kidney and the bladder.
Materials and Methods
Materials
Kenneth's HY, diaminobenzidine tetrahydrochloride (DAB)-H2O2 substrate, and hematoxylin were purchased from Invitrogen (Carlsbad, CA). FBS was obtained from Hyclone (Logan, UT). Hybridoma cloning factor was obtained from Origen, IGN International (Gaithersburg, MD). Mouse IgG isotype control monoclonal was purchased from R&D Systems (Minneapolis, MN). Horseradish peroxidase (HRP)-conjugated goat anti-murine IgG and Permount Mounting Media were obtained from Sigma (St. Louis, MO). Biotinylated goat-anti mouse secondary antibody and HRP-conjugated streptavidin were obtained from Vector (Burlingame, CA). Biotinylated goat-anti-mouse, rat absorbed, secondary antibody was purchased from BioGenex (San Ramon, CA). Electrochemical Luminescence reagent (ECL) and a bicinchoninic acid (BCA) protein assay kit were acquired from Pierce (Rockford, IL). Proteinase inhibitor cocktail and PMSF were obtained from Roche (Indianapolis, IN). Tris-HCl protein gels and polyvinylidene difluoride (PVDF) membranes were obtained from Bio-Rad (Hercules, CA).
Human Tissues
Archival, formalin-fixed, paraffin-embedded human kidney, bladder, prostate, and testis tissues were procured in the Departments of Pathology and Urology, the University of Oklahoma Health Sciences Center. Human tissue specimens were obtained with Institutional Review Board (IRB) approval.
Rat Tissues
Male Sprague-Dawley rats at the age of 1–1.25 years were purchased from Charles River Laboratories (Wilmington, MA). Rat kidney, bladder, prostate, and testis tissues were harvested with a protocol approved by Institutional Animal Care and Use Committee (IACUC). All tissues were immediately either snap frozen in liquid nitrogen for protein extraction or placed in 10% formalin in neutral buffer for 24 hr followed by paraffin embedding for IHC.
Production of a Panel of Monoclonal Antibodies Against AKR1C3 and Rat Homolog
Hybridomas were produced in the Cell Center at the University of Pennsylvania as previously reported (Lin et al. 2004). After screening of culture media from >2000 hybridoma clones by ELISA on 96-well ELISA plates containing immobilized recombinant AKR1C1–AKR1C4 (Lin et al. 2004), a total of 45 subclones were ultimately selected. Four hybridoma clones were tested in this study: NP6G6.A6, NP4H10.D5, R5E9.C3, and R9F9.H1. Hybridoma clones were maintained in hybridoma growth media consisting of Kenneth's HY supplemented with 20% FBS, 2%
Immunoblot Analysis
Frozen rat tissues were powdered with mortar and pestle. Tissue proteins were extracted with RIPA lysis buffer supplemented with proteinase inhibitor cocktail and PMSF as reported previously (Fung et al. 2006). AKR1C9 was overexpressed in Escherichia coli; recombinant AKR1C9 protein was prepared based on previous description (Pawlowski and Penning 1994). Protein concentration was determined by a bicinchoninic acid (BCA) protein assay kit. Extracted tissue proteins (30 μg) along with AKR1C9 (100 ng to 1 mg) were electrophoresed on 10% Tris-HCl gels, and separated proteins were electro-transferred onto PVDF membranes. The protein membranes were incubated with TBS containing 0.1% Tween and 5% fat-free milk for 2 hr to block nonspecific binding. The membranes were incubated individually with the four clones of monoclonal antibody in the blocking buffer followed by HRP-conjugated goat anti-mouse IgG secondary antibody incubation. Immunoreactive protein was detected using the ECL reagent according to the manufacturer's recommendations.
IHC of Tissue Sections
IHC of human and rat tissue sections followed our previously reported procedures (Fung et al. 2006). Briefly, tissue sections cut ∼4–6 μm were mounted and baked at 60C for 1 hr. Sections were deparaffinized with xylene and re-hydrated in graded ethanol. Endogenous peroxidase activity was blocked by incubating the tissue sections with 1.6% H2O2 in methanol for 30 min. Antigen retrieval was performed with 10 mM sodium citric acid buffer (pH 6.0) at 95C for 1 hr. Nonspecific binding was blocked by incubating the tissue sections with 0.1 M Tris-HCl (pH 7.6) containing 10% goat serum for 2 hr. Human AKR1C3 was detected by NP6G6.A6 antibody at a 1:200 dilution, and rat homolog was detected by R9F9.H1 antibody at 1:50 dilution. Both antibodies were diluted with the above blocking buffer as diluent and incubation in a moist chamber at 4C overnight. After washing, the slides were treated with ready to use rat absorbed biotinylated goat-anti mouse secondary antibody and incubated at room temperature for 2 hr, followed by rinses with 0.1 M Tris-HCl. As negative controls, rat tissue sections were incubated with either mouse IgG isotype control monoclonal antibody (1:50 dilution or 10 μg/ml) followed by secondary antibody incubation or directly by secondary antibody alone. Antibody binding was detected by incubating the tissue sections with HRP-conjugated streptavidin at room temperature for another 30 min. DAB-H2O2 substrate was added to the slides and incubated at room temperature for an additional 4 min. Tissue sections were counterstained lightly with hematoxylin. Slides were dehydrated and sealed with Permount Mounting Media (Richard-Allan Scientific; Kalamazoo, MI) for visualization by light microscopy.
Results
Specificity of Monoclonal Antibody for Rat AKR1C3 Homolog Using Immunoblot Analysis
ELISA screening led to the identification of a clone of antibody (NP6G6.A6) that can distinguish AKR1C3 from AKR1C1, AKR1C2, and AKR1C4 (Lin et al. 2004). NP4H10.D5 recognized both AKR1C1 and AKR1C3. R9F9.H1 recognized AKR1C1, AKR1C2, and AKR1C3. R5E9.C3 recognized all four human AKR1C isoforms (data not shown). Immunoblot analysis was conducted to determine whether these clones can recognize rat tissue proteins. NP6G6.A6, NP4H10. D5, and R5E9.C3 failed to detect any protein band for rat tissues or recombinant AKR1C9 (data not shown). In contrast, R9F9.H1 recognized a single protein band with the correct molecular mass, 37 kDa, in immunoblot analysis for all rat tissues analyzed except in the kidney (Figure 1). In the rat kidney, a second band of molecular mass 74–75 kDa was detected. This band was only seen within the kidney and corresponds to roughly double the mass of the other band. Because of the non-reducing conditions of the gel used for immunoblot analysis, it is possible that the rat kidney possesses a dimeric form of AKR1C. However, R9F9.H1 failed to recognize recombinant AKR1C9 protein (data not shown). Recombinant proteins from the other six identified rat AKR1C isoforms were unavailable.
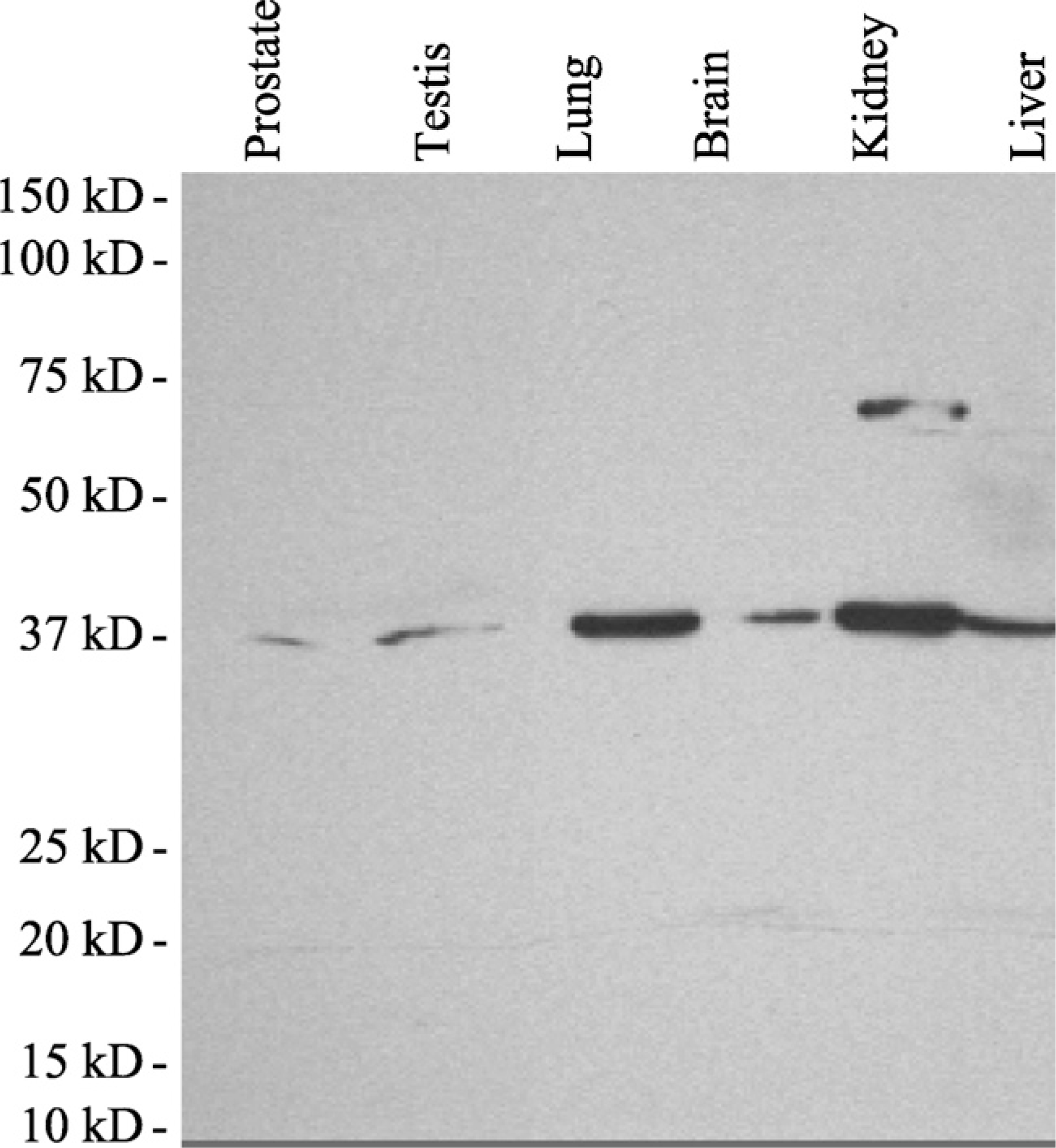
Specificity of a monoclonal antibody for a rat aldo-keto reductase (AKR)1C3 homolog. Total cellular proteins were extracted from rat tissues, electrophoresed on a Tris-HCl gel, and transferred to polyvinylidene difluoride (PVDF) membranes for immunoblot analysis. Detection of a rat homolog of human AKR1C3 in rat protein was performed by incubating the membranes with clone R9F9.H1. The immunoblot analysis shows that R9F9.H1 only reacts with one species of protein with ∼37 kDa prepared from rat tissue lysates in all samples except the kidney. In the kidney, a second band of ∼74–75 kDa is seen. This band corresponds to about double the molecular mass of the low molecular mass band.
Detection of AKR1C3 and Its Rat Homolog in the Genitourinary System
Prostate
As described before (Fung et al. 2006), the immunoreactivity of human AKR1C3 in normal human prostate was limited mainly to the stromal cells and a small number of basal cells (Figure 2A and arrow in Figure 2C) and a small number of basal cells (Figure 2C, arrowhead). There was also strong immunoreactivity in endothelial cells similar to what we reported before (data not shown). Rat tissue sections from the prostate, kidney, bladder, and testis incubated with the IgG isotype control monoclonal antibody or secondary antibody alone failed to show any immunoreactivity (data not shown). In rat prostate tissue, there was strong immunoreactivity of the prostatic epithelial cells (Figure 2B and arrow in Figure 2D) but not the stromal cells (Figure 2D, curved arrow) or endothelial cells (Figure 2D, arrowhead).
Kidney and Urinary Bladder
In the human renal cortex, endothelial cells of the glomeruli lacked immunoreactivity for AKR1C3 (Figure 3A), but the Bowman's capsule (Figure 3A, arrow) and mesangial cells (Figure 3A, arrowhead) were strongly reactive. Some of the tubules, most consistent with proximal convoluted tubules, were immunoreactive (Figure 3A, curved arrow), and there was also positive nuclear staining, whereas others were either non-reactive or weakly reactive (Figure 3A). In the medulla, tubules with larger epithelial cells showed strong immunoreactivity (Figure 3C, arrow) similar to those in the renal cortex. Tubules with thin endothelial cells were only weakly immunoreactive or negative. These features suggested that the collecting tubes were not immunoreactive. The urothelial epithelium lining the renal pelvis was strongly immunoreactive (Figure 3E, arrow and inset), but stromal cells in the underlying supporting connective tissue were negative. Note that vascular endothelial cells (Figures 3C and 3E, arrowheads) showed strong immunoreactivity in both cytoplasm and nuclei, similar to what we described in the prostate (Fung et al. 2006).
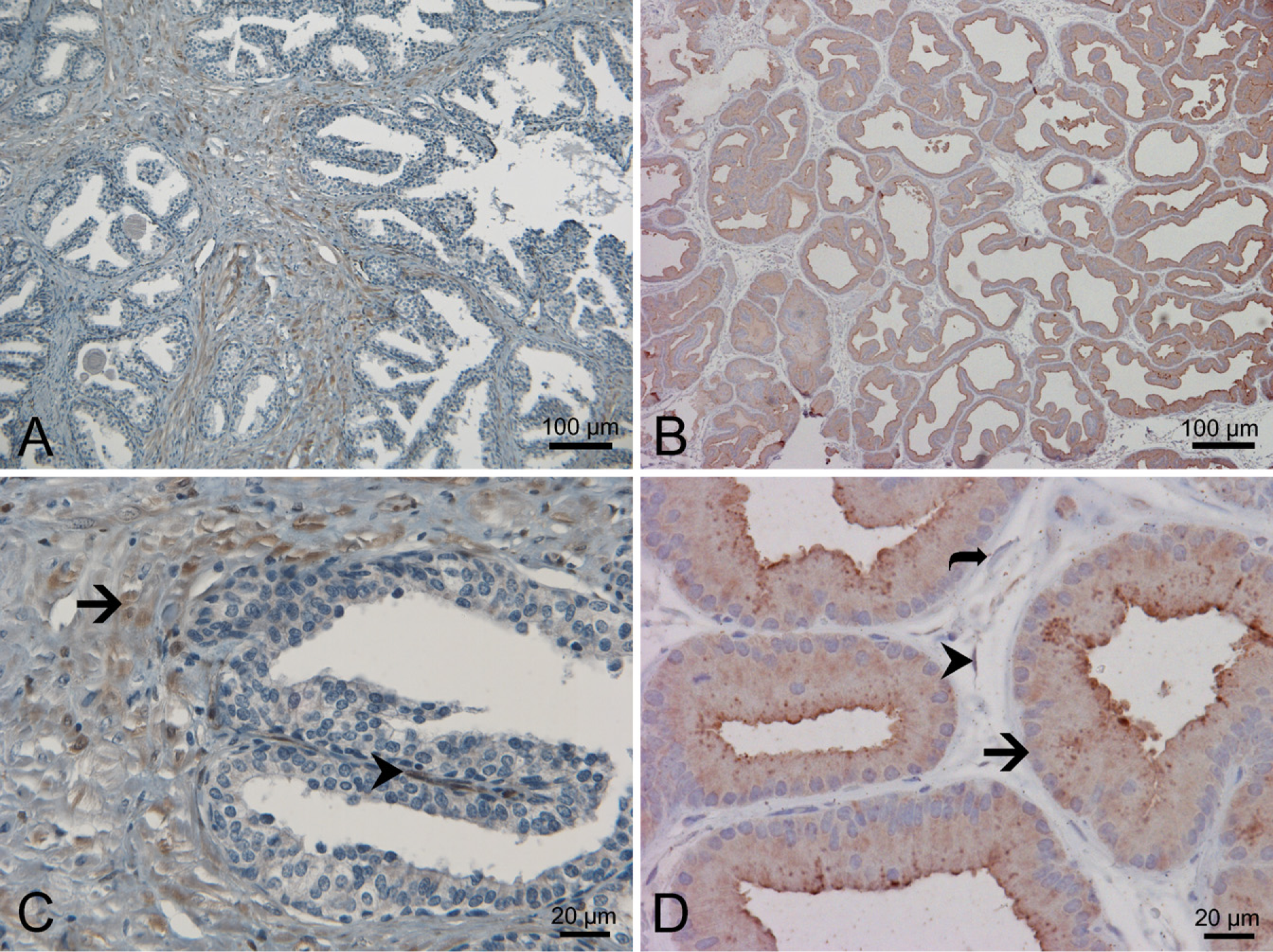
Distribution of AKR1C3 and rat homolog in the prostate. IHC with antibody NP6G6.A6 against human AKR1C3 on human prostate (
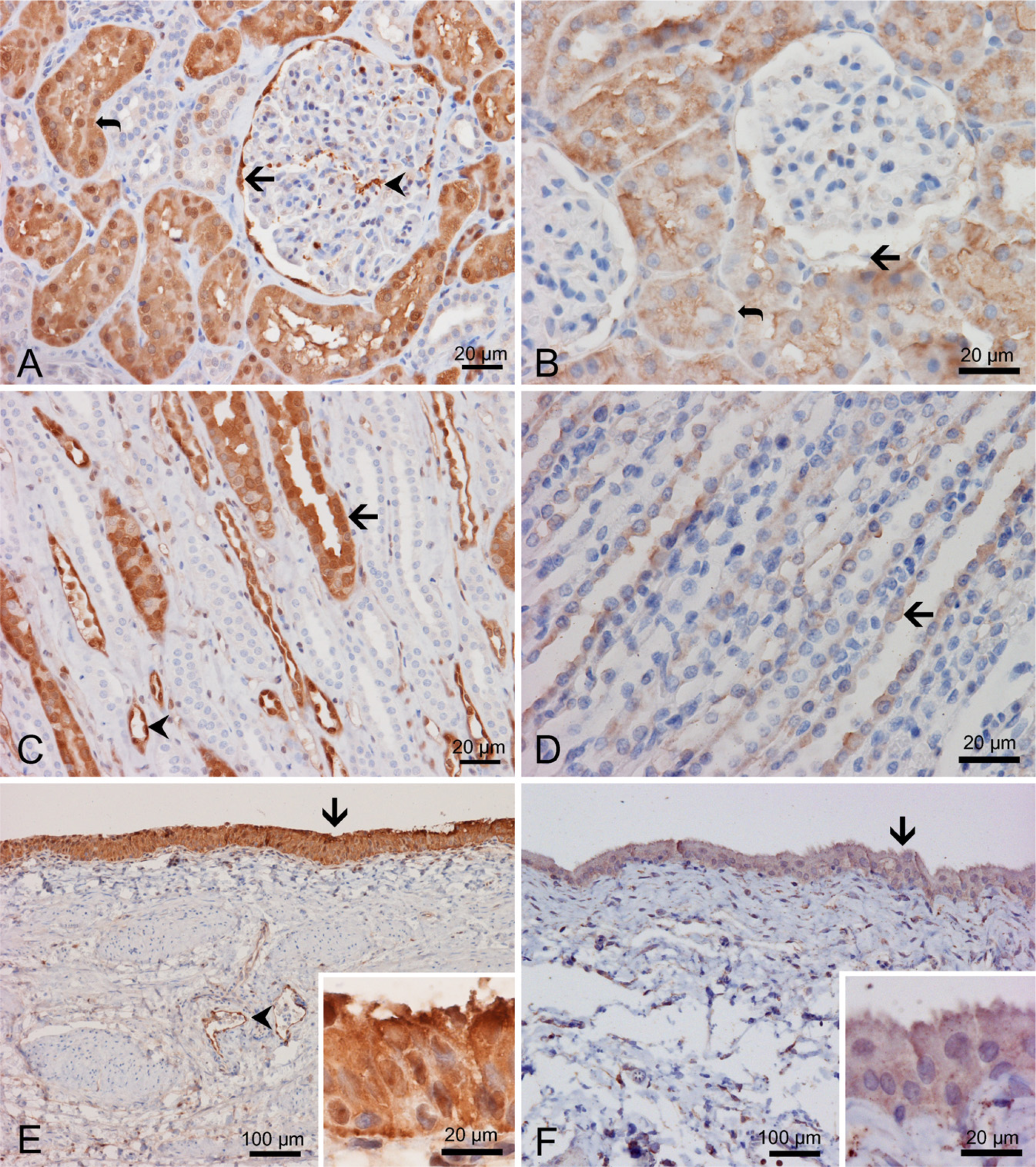
Distribution of AKR1C3 and rat homolog in the kidney and bladder. IHC with antibody NP6G6.A6 against human AKR1C3 on human kidney (
In the rat kidney, a similar but not identical pattern of expression for a rat AKR1C3 homolog was shown. Similar to the human renal cortex, the endothelial cells of glomeruli lacked immunoreactivity. In contrast to the human kidney, the Bowman's capsule (Figure 3B, arrow), mesangial cells, and endothelial cells of regular blood vessels were non-reactive (Figure 3B). The proximal convoluted tubules were immunoreactive (Figure 3B, curved arrow). In the renal medulla, some tubules were immunoreactive (Figure 3D, arrow), but many were not reactive. The pattern of immunoreactivity was similar to that seen in humans. Similar to human tissue, the urothelium of the urinary bladder was strongly immunoreactive (Figure 3F).
Testis and Epididymis
Strong nuclear and cytoplasmic AKR1C3 immunoreactivity was detected in the Leydig cells of human testis (Figure 4A, thick arrow) in the intertubular region and fibromyocytes (myoid cells) around seminiferous tubules (Figure 4A, curved arrow) in the peritubular region. However, the germ cells (Figure 4A, arrowhead) and Sertoli cells were non-reactive or, at the most, minimally reactive. Similar to the aforementioned observations in the prostate and kidney, the endothelial cells of blood vessels were strongly immunoreactive. In rat testis (Figure 4B), the pattern of immunoreactivity seemed to be reversed in comparison to the human testis. The Leydig cells were only weakly positive both in the cytoplasm and the nuclei (Figure 4B, arrow). There was strong immunoreactivity in the germ cells and Sertoli cells (Figure 4B, arrowhead). No endothelial cell reactivity was detected in rat testicular tissue.
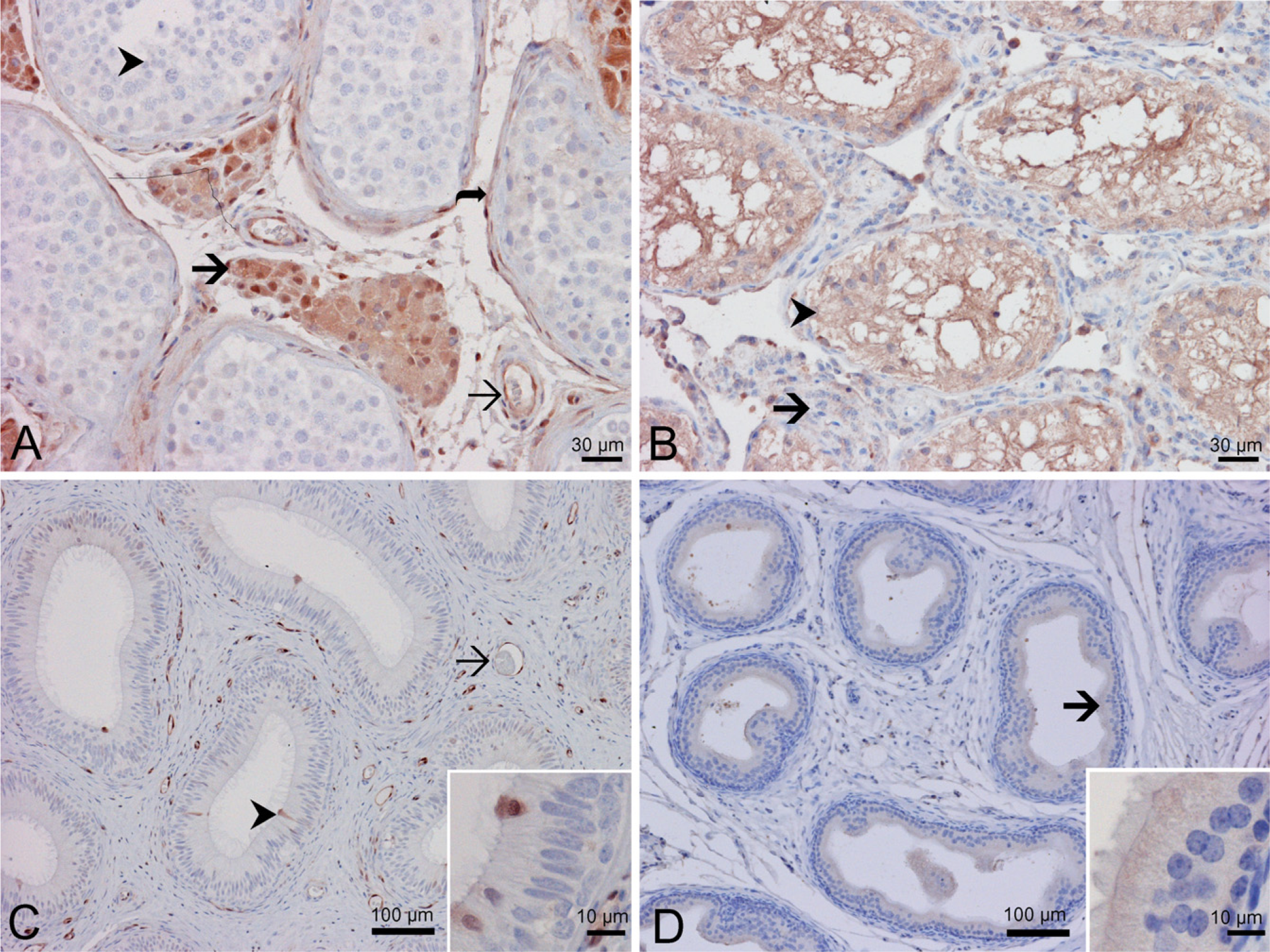
Distribution of AKR1C3 and rat homolog in the testis. Immunohistochemistry with antibody NP6G6.A6 against human AKR1C3 on human testis (
In human epididymis (Figure 4C), only a small number of epithelial cells were immunoreactive in the epididymal epithelium with both nuclear and cytoplasmic reactivity, and the nuclei of most of these were located close to the luminal surface (Figure 4C, arrowhead and insert). There was strong immunoreactivity in the endothelial cells (Figure 4C, arrow) and some stromal cells. In rat epididymis, there was no reactivity in the stromal cells or endothelial cells. A diffuse but weak immunoreactivity was shown in the epididymal epithelial cells (Figure 4D, arrow and inset).
Discussion
Based on increasing knowledge about AKR1C3 in hormone metabolism and in interaction with various environmental factors, this enzyme may play a critical role in cancer development. Recent reports on deregulated expression of AKR1C3 in multiple types of cancers, including classical hormone-related cancers (Lewis et al. 2004; Fung et al. 2006; Rizner et al. 2006) and non—hormone-related cancers (Lan et al. 2004; Mahadevan et al. 2006), may reflect the above hypothesis. We have suggested that AKR1C3 is an enzyme that is poised to govern steroid hormone action at the prereceptor level by governing ligand access to the appropriate nuclear receptor, including the androgen receptor (AR), the estrogen receptor (ER), and the PPARγ, in hormone target tissues (Penning 2003; Penning et al. 2006). Tissue distribution of AKR1C3 would be important to understand and predict potential biological functions of this enzyme in multiple tissues. In this report, the distribution of human AKR1C3 and a rat homolog protein expression in the genitourinary organs by IHC was described.
The four human AKR1C isoforms have been well studied for their hormone metabolism. Because AKR1C1–AKR1C4 share high amino acid sequence identity, these isozymes actually may share some common domains that can be recognized by one monoclonal antibody. Although multiple rat isoforms including AKR1C8, AKR1C9, AKR1C15, AKR1C16, AKR1C17, AKR1C18, and AKR1C24 have been cloned and identified, only AKR1C9 has been well characterized for its hormone metabolism (Penning and Sharp 1987; Pawlowski et al. 1991). AKR1C3 and AKR1C9 do not catalyze the identical reactions in androgen metabolism. For example, AKR1C3 only reduces 5α-DHT to form 3α-diol, whereas AKR1C9 interconverts 5α-DHT and 3α-diol (Lin et al. 1997). A monoclonal antibody raised against AKR1C3 may, therefore, not recognize AKR1C9. Because of the lack of recombinant proteins for other rat AKR1C isozymes or complete enzyme kinetics, we could not specifically pinpoint which rat isoform may share the same epitope with AKR1C3 and is recognized by R9F9.H1.
IHC staining showed the expression of AKR1C3 in both stromal and endothelial cells but not in epithelial cells in normal prostate tissue. In contrast, we described elevated AKR1C3 expression in prostatic adenocarcinoma cells (Fung et al. 2006). Current hypotheses are that the basal and luminal epithelial cells contain 5α-reductase type 2 and the AR, respectively (Barbier et al. 2000). When these cells become transformed, AKR1C3 may be poised to covert androstenedione to testosterone in these cells. AKR1C3 may also be playing a role in gland maintenance within the prostate and in sexual response with stromal contraction during ejaculation. Although general functions of the prostate remain the same for rats as they are in humans, the differences in immunoreactivity we observed are most likely the result of variance between the two species, particularly copulatory differences. The lack of immunoreactivity of the AKR1C3 rat homolog in the stromal cells of rat prostate may be related to the intrinsic lack of smooth muscle. The positive immunoreactivity in epithelial cells suggests either this antibody recognizes another AKR1C isoform with different function or that, in contrast to human, the AKR1C3 rat homolog is necessary in maintaining the function of rat prostatic epithelial cells.
The cells of the human renal cortex expressing AKR1C3 indicate that this enzyme may be playing a role in regulating glomerular filtration (Campean et al. 2003; Stichtenoth et al. 2005; Glodny and Pauli 2006; Matzdorf et al. 2007). AKR1C3 might also be affecting the kidney's water and solute reabsorption rate, particularly in the proximal convoluted tubules, because fewer tubules of the renal medulla showed positive immunoreactivity (Hebert et al. 1991; Vitzthum et al. 2002; Qi et al. 2006). The rat AKR1C3 homolog's similar expression pattern in the renal cortex could mean it is playing a similar role in solute reabsorption in the proximal tubule. The slight differences in expression in the glomeruli also suggest that it might not be playing a role in initial filtration in rat kidney.
AKR1C3 and its rat homolog are strongly expressed by the urothelium. To date, little is known about the role of AKR1C3 in the urothelium. The urothelium has been shown to be the main source of PGs within the bladder (Krasnopolsky et al. 1995), and it has been suggested that these PGs could be related to a stretch sensing role during bladder filling (Azadzoi et al. 2004; Schroder et al. 2004; de Jongh et al. 2007). Alternate roles for AKR1C3, and its rat homolog, in the bladder could be for the creation of a watertight barrier for confining urine and as a possible regulator controlling the inflammatory effects of urine (Oleksiewicz et al. 2005). AKR1C3, and its rat homolog, may as also be serving to facilitate the turn over of old epithelial cells with new ones in a fashion similar to that seen within the intestines (Rakoff-Nahoum and Medzhitov 2007).
In the testis of the human, the Leydig cells' positive immunoreactivity for AKR1C3 is expected because these cells are responsible for the production of androgens, including testosterone. AKR1C3 may be playing a key role in providing the necessary trophic components and metabolites for the testis to function properly (Leihy et al. 2004). Because the immunoreactivity pattern for the rat AKR1C3 homolog is reversed from AKR1C3 in the human testis, it could mean that one of the seven other isoforms of the AKR1C superfamily found in rats is governing Leydig cell androgen conversions.
The general observation of this compare and contrast study is that the expression of AKR1C3 and its rat homolog have an almost identical pattern in the urologic tissues: the kidney and urinary bladder. In contrast, the staining pattern is the reverse of humans compared with the rat tissues in the genital organs, the testis, and prostate. These generalized observations indicate that the two monoclonal antibodies may recognize the same molecule, which explains the almost identical staining pattern in kidney and bladder, albeit the same molecule seems to have different, and perhaps opposite roles, in the genital organs such as the testis and prostate.
One significant note about the expression of AKR1C3 in the various human tissues is that the endothelial lining of non-specialized blood vessels showed strong positive immunoreactivity. The exact role that AKR1C3 is playing in blood vessels is unknown. With the suggested involvement of AKR1C3 in PG metabolism, the role of AKR1C3 in endothelial cells may be closely related to and/or regulated by the PG metabolism (Matsuura et al. 1998; Desmond et al. 2003). Recent studies have attempted to illuminate the possible roles that PGs are playing in regulation of blood vessels, especially with regards to angiogenesis. The rat tissues' lack of positive immunoreactivity for the AKR1C3 homolog in blood vessels leaves no definitive conclusions as to its role in rodent blood vessels. In contrast, the endothelial cells of the renal glomeruli lack immunoreactivity for AKR1C3 in human and rat kidney.
Although we have previously shown that AKR1C3 and its related mRNA species are extensively expressed in human tissues (Lin et al. 1997), IHC identifies specific cell types within each tissues and raises questions about previously undefined roles of AKR1C3 in the genitourinary system. Of particular note, tissues that are not classically classified as being hormonally dependent also express AKR1C3. Another interesting point is that all of the human tissues studied in this report were normal, non-cancerous, tissues. Most work to date has focused on AKR1C3's expression in cancer, particularly cancer in endocrine tissues such as the prostate and breast. These data suggested that AKR1C3 may play significant roles in regulating physiological function and pathological development of diseases that require further study. It was also hoped that this particular rat antibody, raised against AKR1C3, could help to serve in future studies and help establish an animal model. However, the variance in immunoreactivity patterns and failure of the rat AKR1C3 homolog antibody (R9F9.H1) to recognize recombinant AKR1C9 leave its exact specificity in doubt.
Footnotes
Acknowledgements
This work was supported by the U.S. Department of Veterans Affairs Merit Award to H.-K.L.