
Abstract
During mineralization of the avian eggshell, there is a sequential and orderly deposition of both matrix and mineral phases. Therefore, the eggshell is an excellent model for studying matrix–mineral relationships and the regulation of mineralization. Osteopontin, as an inhibitor of crystal growth, potently influences the formation of calcium phosphate and calcium carbonate biominerals. The purpose of this study was to characterize matrix–mineral relationships, specifically for osteopontin, in the avian eggshell using high-resolution transmission (TEM) and scanning (SEM) electron microscopy to gain insight into how calcite crystal growth is structured and compartmentalized during eggshell mineralization. Osteopontin was localized at the ultrastructural level by colloidal-gold immunocytochemistry. In EDTA-decalcified eggshell, an extensive matrix network was observed by TEM and SEM throughout all regions and included interconnected fibrous sheets, irregularly shaped aggregates, vesicular structures, protein films, and isolated protein fibers. Osteopontin was associated with protein sheets in the highly mineralized palisades region; some of these features defined boundaries that compartmentalized different eggshell structural units. In fractured and un-decalcified eggshell, osteopontin was immunolocalized on the [104] crystallographic faces of calcite—its natural cleavage plane. The specific occlusion of osteopontin into calcite during mineralization may influence eggshell structure to modify its fracture resistance.
M
The eggshell is ∼96% calcium carbonate mineral, whereas the remaining organic material is distributed throughout the shell as a proteinaceous matrix (3.5%, with the remainder as water), of which approximately one half can be readily solubilized by decalcification of the shell. The native and soluble precursors of the eggshell matrix are present in the uterine fluid, where the protein composition varies during the initial, calcification, and terminal phases of eggshell deposition (Gautron et al. 1997). Proteomic analysis has identified >500 eggshell matrix proteins (Mann et al. 2006), with the most abundant corresponding to those previously identified and categorized as follows. Eggshell-specific proteins, such as the ovocleidins and ovocalyxins, were originally identified by N-terminal amino acid sequencing and immunochemistry. One of these, ovocleidin-116, has been cloned and found to correspond to the protein core of a novel dermatan sulfate proteoglycan (Hincke et al. 1999). Another ovocleidin, ovocleidin-17, is a C-type lectin-like phosphoprotein related to pancreatic stone protein, which occurs in glycosylated and non-glycosylated forms in the shell matrix (Hincke et al. 1995; Mann 1999; Mann and Siedler 1999). Ovocalyxin-32 is a 32-kDa protein with similarity to the mammalian carboxypeptidase inhibitor latexin and the human skin protein RAR-TIG1 (retinoic acid receptor-responsive, tazarotene-induced gene 1) (Gautron et al. 2001a). The sequence of another ovocalyxin, ovocalyxin-36, has homology to proteins associated with the innate immune response, such as lipopolysaccharide-binding proteins, bactericidal permeability-increasing proteins, and palate, lung, and nasal epithelium clone (Plunc) family proteins (Gautron et al. 1997). The egg white proteins—ovalbumin, lysozyme, and ovotransferrin—are also present in the uterine fluid and are primarily localized in the shell membranes and in the mammillary cone layer of the eggshell (Hincke et al. 1995, 2000; Gautron et al. 2001b). Last, osteopontin, a mineralized tissue protein found in bone, teeth, and cartilage, is also a prominent phosphoprotein of the eggshell matrix (Pines et al. 1994; Fernandez et al. 2003; Mann et al. 2007). Sequential incorporation of matrix proteins into the mineralizing eggshell results in their differential localization between the inner (mammillary) and outer (palisade) layers of the mineralized shell (Hincke et al. 1992).
We hypothesize that the incorporation of shell matrix proteins into specific eggshell compartments regulates calcite mineral growth and provides eggshell with its requisite structure and functional properties to afford protection to the embryo from potentially harmful environmental factors. The soluble proteins of calcitic matrices modify crystal growth and thus regulate the macroscopic structure and biomechanical properties of the resulting bioceramic. In mollusk shells, specific proteins control mineral phase switching between the calcite and aragonite forms (Belcher et al. 1996). Partially purified eggshell matrix proteins inhibit calcium carbonate precipitation and alter patterns of calcite crystal growth; however, the role of individual matrix proteins is unknown (reviewed in Nys et al. 2004).
Osteopontin is associated with mineralization in all mammalian hard tissues except tooth enamel (McKee and Nanci 1996). In the chicken, uterine expression of the osteopontin gene is temporally associated with eggshell mineralization through a coupling of physical distension of the uterus with osteopontin gene expression (Lavelin et al. 1998). Partially purified chicken eggshell osteopontin strongly inhibits calcium carbonate precipitation in a phosphorylation-dependent manner (Hincke and St. Maurice 2000), suggesting that it could potentially have a significant influence over eggshell mineralization in vivo. To learn more about how eggshell osteopontin affects mineralization in biological systems, we studied its ultrastructural localization pattern in the shell gland and within the eggshell matrix of the domestic chicken.
Materials and Methods
Gel Electrophoresis and Western Blotting
White Leghorn hens were individually caged and maintained under conditions approved by the animal care committee of the Animal Research Centre, Agriculture Canada, Ottawa, Canada, as previously described (Tsang et al. 1990). Unfertilized eggs laid by White Leghorn hens were obtained from commercial sources. Powdered tibial bone (from 18-day chicken embryos), eggshell, and uterine tissue from the mid-uterus of domestic White Leghorn chickens (
Light Microscopy, Transmission Electron Microscopy, and Immunocytochemistry
Eggshell (as above) and shell gland (uterus) taken from a laying hen with an egg in mid-calcification within the shell gland were fixed and processed in LR White acrylic resin (London Resin Company; Berkshire, UK) as previously described (McKee and Nanci 1995). Survey sections (0.5 μm) of embedded tissue cut with a diamond knife on an ultramicrotome (model Reichert Ultracut E; Leica, Wetzlar, Germany) were stained with toluidine blue and coverslipped. Light micrographs were obtained using a DXC-950 3-CCD camera (Sony; Tokyo, Japan) mounted on an optical microscope (model Leitz DMRBE; Leica). For transmission electron microscopy (TEM), selected regions were trimmed, and ultrathin sections (80 nm) were placed on polyvinyl formal- and carbon-coated nickel grids. Grid-mounted tissue sections were processed for colloidal-gold immunocytochemistry by incubation of the sections with primary antibody (1/10 dilution), after which immunolabeling patterns were visualized by incubation with protein A-colloidal gold complex (14 nm gold particles; Dr. G. Posthuma, University of Utrecht, Utrecht, The Netherlands), followed by conventional staining with uranyl acetate and lead citrate, as described previously (McKee and Nanci 1995). Incubated grids were examined in a JEM 2000FXII transmission electron microscope (JEOL; Tokyo, Japan) operated at 80 kV.
Scanning Electron Microscopy and Immunocytochemistry
For morphological imaging of undecalcified eggshell, freshly fractured eggshell fragments were dried in air and mounted with conductive carbon cement onto metallic scanning electron microscopy (SEM) stubs to provide a transverse cross-section view of eggshell/membrane. Fractured eggshell fragments were also decalcified in an 8% EDTA solution with 1% glutaraldehyde and sequentially dehydrated with ethyl alcohol and hexamethyldisilazane or dehydrated by critical point drying for imaging of eggshell matrix organization. Samples were sputter-coated with a 20- to 25-nm-thick Au-Pd thin film and imaged using a Hitachi field-emission gun scanning electron microscope (FE-SEM) operating at an accelerating voltage of 5 kV (model S-4700; Hitachi High Technologies America, Pleasanton, CA). To localize osteopontin in eggshell matrix and to investigate eggshell ultrastructure and matrix–mineral relationships, we used SEM coupled with immunogold labeling for osteopontin. Briefly, undecalcified and aldehyde-fixed, or EDTA-decalcified and fixed, eggshell fragments were incubated with primary antibody against osteopontin and protein A–colloidal gold complex as above. The samples were coated with a 40- to 60-nm-thick carbon layer and examined by FE-SEM using both secondary and back-scattered electron imaging modes while operating at an accelerating voltage of 9 kV.
Results
In these studies, we used a well-characterized antibody raised against chicken bone osteopontin (Gotoh et al. 1995). Powdered eggshell and chicken bone were sequentially extracted to obtain (a) extra-mineral proteins soluble in 4 M guanidine HCl, (b) mineral-bound proteins soluble in dilute acid, (c) mineral-bound proteins that are acid insoluble but soluble in neutral high salt conditions, and (d) mineral-bound proteins that are finally solubilized in 4 M guanidine HCl. Western blotting of these extracts showed that eggshell osteopontin is only present in the dilute acid mineral-bound extract; the majority of bone osteopontin was extractable under the same conditions. Osteopontin exists as two to three predominant forms in both eggshell and bone, with an apparent size ranging between 46 and 54 kDa (Figure 1). Slight differences in molecular mass and proportions of each form indicate that bone and eggshell osteopontin differ in their posttranslational modifications. We have previously shown, by microsequencing of partially purified eggshell osteopontin, that serine residues 12, 14, and 15 are phosphorylated (Hincke and St. Maurice 2000). Metabolic 32P labeling studies on osteopontin secreted by cultured chicken osteoblasts also identified the same phosphorylated residues (Salih et al. 1997). In addition to phosphorylation, mammalian osteopontins have been found to exhibit various degrees of glycosylation, glycanation, and/or sulfation (Prince et al. 1987; Nagata et al. 1989; Sodek et al. 1995; Masuda et al. 2000). These posttranslational modifications alter the apparent molecular mass of osteopontin as determined by SDS-PAGE (Kazanecki et al. 2007).
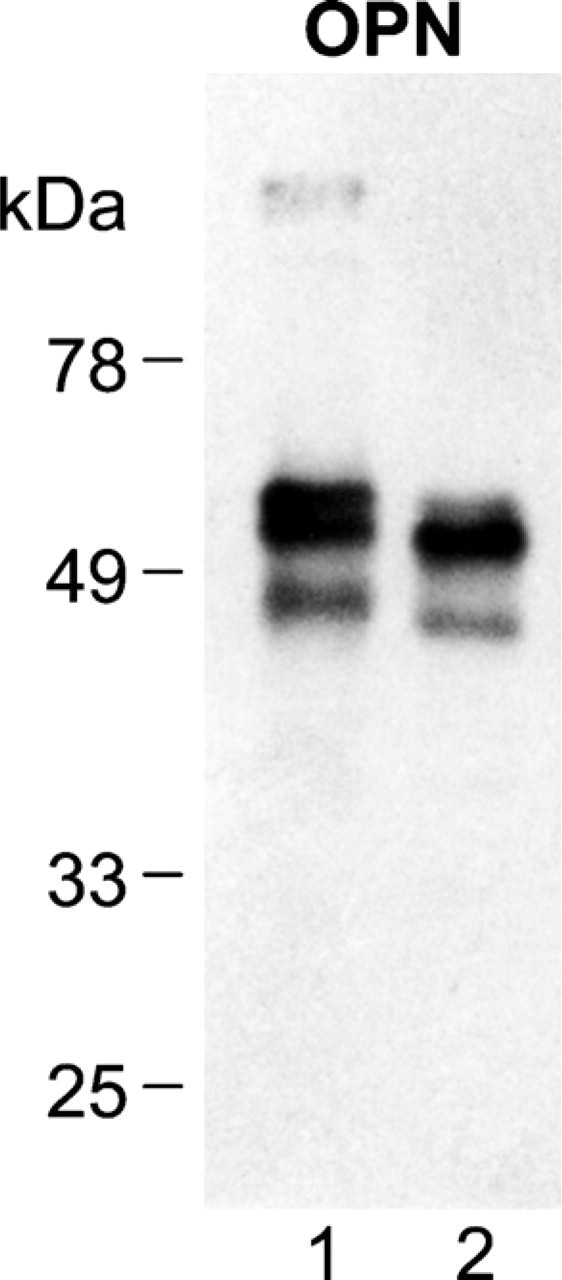
Western blotting of osteopontin extracted from eggshell and bone matrix. Acid extract of mineral-bound proteins from tibial bone (Lane 1) and eggshell (Lane 2), prepared as described in Materials and Methods. Bone extract, 25 μg; shell extract, 25 μg.
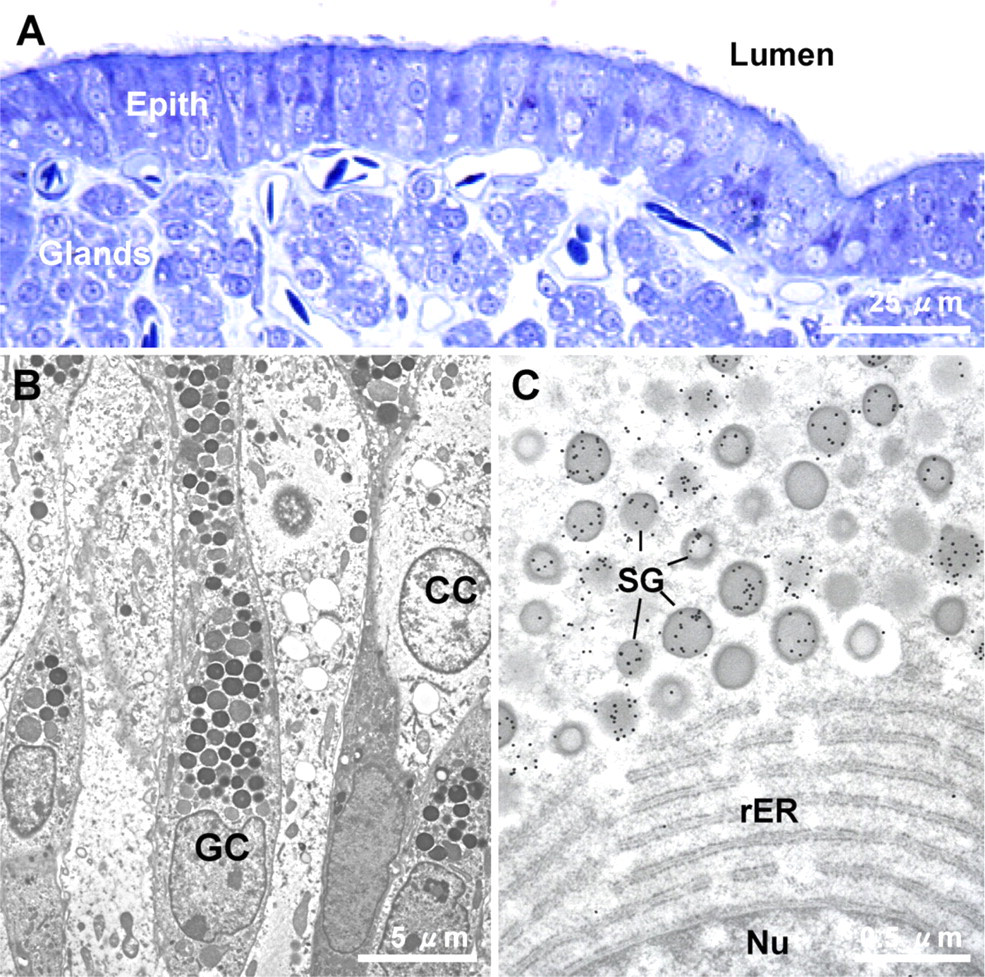
Shell gland mucosal histology by light and electron microscopy. (
The cellular source of eggshell osteopontin was studied within the uterine mucous membrane by immunogold labeling and TEM. Ciliated (clear) and granular (non-ciliated) cells constitute the surface epithelium that lines the lumen, as shown in Figure 2A by light microscopy. Colloidal-gold immunocytochemistry showed that secretion granules in the granular cells were intensely immunopositive (Figures 2B and 2C). Ciliated cells and cells of the subjacent tubular glands of the shell gland mucosa were not immunolabeled (data not shown). These results indicate that osteopontin is synthesized and secreted by the granular epithelial cells of the shell gland, a finding in agreement with conclusions previously drawn from in situ hybridization and immunohistochemical studies performed at the light microscopic level (Pines et al. 1994; Fernandez et al. 2003). The granular cells also synthesize and secrete the novel dermatan sulfate proteoglycan of the eggshell matrix, ovocleidin-116 (Hincke et al. 1999), suggesting that this epithelial cell type is a key player in the secretion of the eggshell matrix.
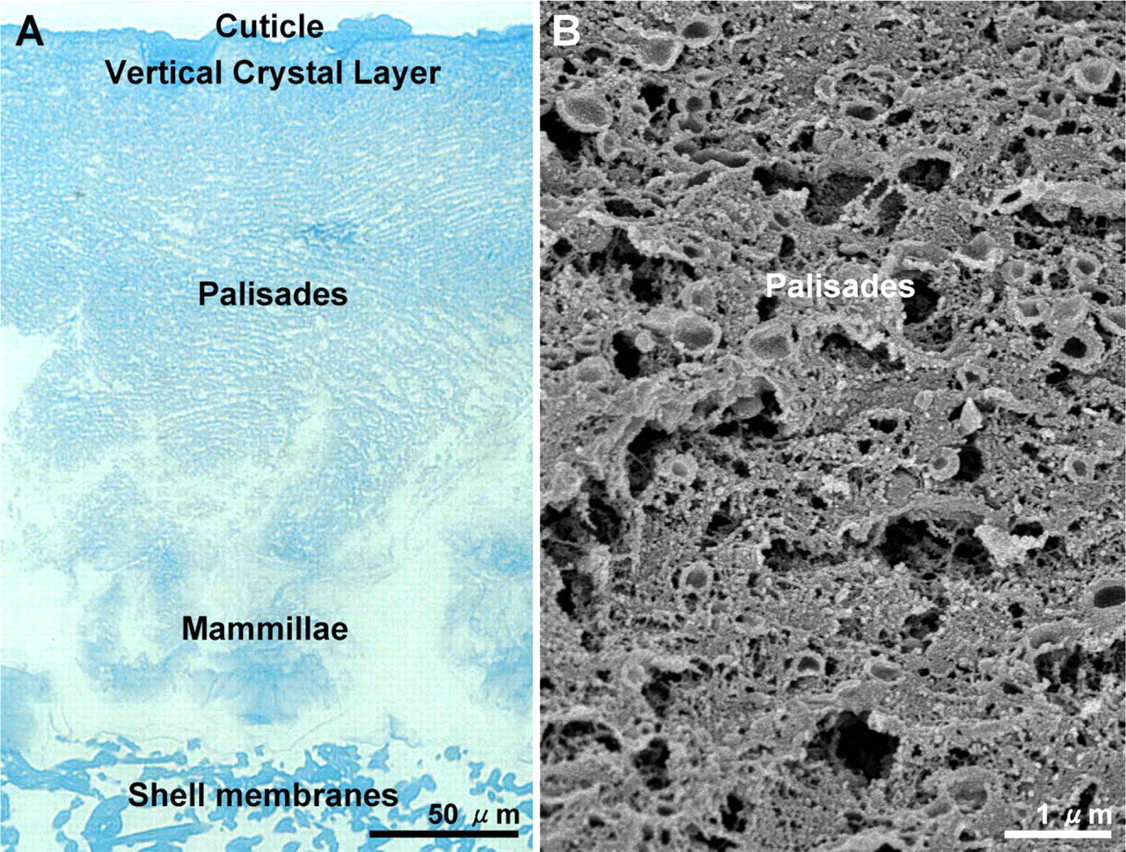
Examination of cross-fractured eggshell by SEM showed the overall structure of the calcified eggshell that forms on the shell membranes (Figure 3A). The mammillary layer is composed of a regular array of cones or knobs; at higher magnification, the individual fibers of the outer eggshell membrane are seen to penetrate the tips of these structures (see also Hincke et al. 2000). The palisade layer consists of groups of aligned calcitic columns (or prisms) that are perpendicular to the eggshell surface and extend outward from the mammillary cones. This layer ends at the vertical crystal layer having a mineral texture differing from that of the palisade region. At higher magnification, the palisades show extensive planes of cleaved calcite and abundant spherical voids (Figures 3B and 3C). The overall architecture of the strikingly abundant matrix that coexists with the mineral phase in all regions of the eggshell was shown in decalcified shell prepared for histology (Figure 4A). At higher magnification of the palisades region, SEM showed details of the matrix presenting as an extensive network of protein sheets and fibers and also as small spherical granules/vesicles dispersed within this matrix (Figure 4B).

Calcified eggshell structure. (
Matrix architecture in eggshell. Micrographs of a chemically fixed, fully decalcified eggshell preparation sectioned for histology (
Osteopontin immunogold labeling and TEM of thin sections of decalcified eggshell were performed to determine its localization within the organic matrix. This approach showed a prominent concentration of osteopontin in the palisades region (Figure 5), where osteopontin immunolabeling was associated with flocculent and diffuse sheets of organic material (Figures 5A–5C). Gold particles were predominantly associated with the planar sheets of matrix that aligned generally parallel or slightly angled to the eggshell surface (Figures 5A–5C). Unlabeled spherical vesicles ranging in size from 200 to 300 nm, and having electron-lucent centers, were dispersed between and along the sheets of matrix. These structural features of the matrix architecture were also readily apparent by SEM (Figure 4B). SEM performed in the secondary (Figure 5D) or backscattered (Figure 5E) electron imaging mode, for morphology and enhanced gold particle detection, respectively, confirmed osteopontin immunogold labeling along the sheets of matrix. Osteopontin imaging in the palisades of cross-fractured eggshell that was not decalcified was obtained by SEM analysis in both secondary and back-scattered electron imaging modes (Figures 6A and 6B) and showed specific gold particle labeling for osteopontin at the surface of the cleaved calcite along the [104] crystallographic faces.

Osteopontin as a constituent of the eggshell matrix in the palisades region in a decalcified eggshell sample. (
Discussion
In this study, we characterized matrix–mineral relationships in avian eggshell using ultrastructural approaches that included TEM, SEM, and high-resolution immunogold labeling that associated osteopontin with specific structures of the eggshell matrix and with specific crystallographic faces of the calcitic mineral. We also identified the granular epithelial cells of the shell gland mucosa as the source of this osteopontin.
After decalcification and processing of the eggshell for TEM and SEM, the organic matrix of the palisades region of the avian eggshell exhibited two structural features: vesicular structures with electron-lucent cores dispersed along and between flocculent sheets of organic material that aligned generally parallel, or slightly angled, to the eggshell surface. Previous observations on this matrix have led to the proposal that its properties and organization might inhibit crack propagation and add to the overall strength of the shell (Simons and Wiertz 1963; Silyn-Roberts and Sharp 1986; Nys et al. 2004). We observed that osteopontin immunoreactivity was concentrated in the palisades region, where it was almost exclusively associated with the planar sheets of matrix, while being absent from the dispersed spherical vesicles. Notably, in other mineralized tissues, there is likewise a striking accumulation of osteopontin at planar matrix–matrix/mineral interfaces in bones and teeth where osteopontin accumulates at cement lines (reversal and resting lines, actually planes in three dimensions) and is thought to serve in matrix adhesion at these sites or in limiting microcrack propagation (McKee and Nanci 1996). Alternatively, layered osteopontin-containing matrix sheets may reflect a self-assembly mechanism by which calcite growth rates are periodically limited (regulated) in this rapidly mineralizing system—one of the fastest known in biology. Cyclical variations in osteopontin secretion into the uterine fluid, or cyclical phases of accelerated calcite growth, might each contribute to this layered matrix structure. In contrast to the localization of osteopontin in these planar matrix accumulations, previously we have shown that the eggshell-specific matrix protein ovocleidin-116 immunolocalizes to the vesicular structures in the palisades region (Hincke et al. 1999).

Osteopontin as a constituent of the eggshell matrix in the palisades region in an undecalcified and fractured eggshell sample. Scanning electron microscopy performed in the (
The relationship of osteopontin to the calcitic mineral phase was determined by immunolabeling of undecalcified shell followed by visualization of colloidal-gold detection sites using the secondary and backscattered electron imaging modes of SEM. These imaging modalities revealed that occluded osteopontin is present on the surface of cleaved calcite along the [104] crystallographic faces. The elongated calcite crystals in the palisades region tend to be preferentially orientated with their (001) planes parallel (c-axis perpendicular) to the shell surface, which would orient the [104] plane at 44° tangential to the surface (Silyn-Roberts and Sharp 1986; Rodriguez-Navarro et al. 2002). The [104] calcite face is the natural cleavage plane, and specific osteopontin binding to this growing crystal face during mineralization could modify the resistance of the shell to fracture along this plane. In support of this hypothesis, we note that, in sea urchin calcitic adult exoskeletons (test plates, spines) and larval endoskeletal spicules, occluded acidic glycoproteins are specifically adsorbed on the [110] planes and improve the fracture properties of calcite crystals by interfering with the [104] natural cleavage planes (Emlet 1982; Berman et al. 1988; Seto et al. 2004). Whereas the primary function of osteopontin may be to regulate crystal growth patterns and speed by binding to mineral in the eggshell, its incorporation into calcite as an occluded crystal protein might serve the secondary function of also providing some resistance to [104] cleavage.
The relationship of occluded osteopontin observed in undecalcified preparations to that found in the matrix sheets of fully decalcified samples has yet to be determined, although experiments are currently underway using gentle mineral- and protein-etching procedures to decipher their respective contributions. Although it is generally difficult to simultaneously observe both matrix and mineral, we have made some progress in this regard as reported in this study. Further work using etching approaches should provide insight into the coexistence of these phases in eggshell. Likewise, it will be important to study the interaction of osteopontin and other purified eggshell matrix proteins with growing calcite crystals and to study binding partners of osteopontin and self-assembly into a matrix to fully understand these interactions. Finally, we are currently examining whether osteopontin incorporation into the eggshell may also serve some role in the dissolution of the shell to provide calcium to the skeleton of the growing embryo.
Footnotes
Acknowledgements
Funding for this research was provided by the National Sciences and Engineering Research Council of Canada (to MTH and MDM), the Poultry Industry Council (to MTH), the Canadian Institutes of Health Research (to MDM), and the Fonds Quebecois de la Recherche sur la Nature et les Technologies (to MDM).
M.D.M. thanks Isabelle Turgeon, Lydia Malynowsky, and Line Mongeon for expert technical assistance with this project. M.D.M. is a scholar of the Fonds de la Recherche en Sante du Quebec and a member of McGill's Centre for Bone and Periodontal Research (Jamson T.N. Wong Laboratories), Centre for Biorecognition and Biosensors, and Institute for Advanced Materials.