
Abstract
Lactate and ATP formation by aerobic glycolysis, the Warburg effect, is considered a hallmark of cancer. During angiogenesis in non-cancerous tissue, proliferating stalk endothelial cells (ECs) also produce lactate and ATP by aerobic glycolysis. In fact, all proliferating cells, both non-cancer and cancer cells, need lactate for the biosynthesis of building blocks for cell growth and tissue expansion. Moreover, both non-proliferating cancer stem cells in tumors and leader tip ECs during angiogenesis rely on glycolysis for pyruvate production, which is used for ATP synthesis in mitochondria through oxidative phosphorylation (OXPHOS). Therefore, aerobic glycolysis is not a specific hallmark of cancer but rather a hallmark of proliferating cells and limits its utility in cancer therapy. However, local treatment of angiogenic eye conditions with inhibitors of glycolysis may be a safe therapeutic option that warrants experimental investigation. Most types of cells in the eye such as photoreceptors and pericytes use OXPHOS for ATP production, whereas proliferating angiogenic stalk ECs rely on glycolysis for lactate and ATP production.
Keywords
Introduction
Cellular energy metabolism, and in particular synthesis of ATP, is considered to be an attractive therapeutic target in cancer1–7 and ocular diseases.8–10 However, anticancer therapeutic approaches, including clinical trials, to block ATP synthesis in cancer cells either via inhibition of glycolysis or via oxidative phosphorylation (OXPHOS) have not been successful thus far.11–19 In ocular diseases, sparse research has been focused on the inhibition of metabolic pathways, such as glycolysis or OXPHOS, in endothelial cells (ECs) of sprouting capillaries during angiogenesis,8–10,20–23 and thus far, studies in patients have not been reported.24,25
Recently, we evaluated the roles of anaerobic and aerobic glycolysis in ATP synthesis, with lactate as end product or pyruvate as a substrate for OXPHOS (Fig. 1). This investigation was conducted in differentiated cancer cells (dCCs) and cancer stem cells (CSCs) in both human glioblastoma (primary brain tumors) and human leukemia in vivo12,13,26,27, and in human angiogenic tip endothelial cells (ECs) and non-tip ECs in vitro.28,29 Our studies revealed similarities in ATP synthesis pathways between non- or slowly proliferating CSCs residing in their niches and the non-proliferating angiogenic tip ECs. These cell types exhibit a relatively greater reliance on OXPHOS and a relatively lower dependence on aerobic glycolysis for lactate production in ATP synthesis compared with the rapidly proliferating dCCs and non-tip ECs. Besides, energy metabolism in pericytes (PCs) is also an important topic to investigate because of the tight metabolic interactions between ECs and PCs, and OXPHOS is mainly used in PCs for ATP production.30–33 The preferential pathways for ATP synthesis in these different cell types, either anaerobic or aerobic glycolysis and/or OXPHOS, are indicated in Table 1.
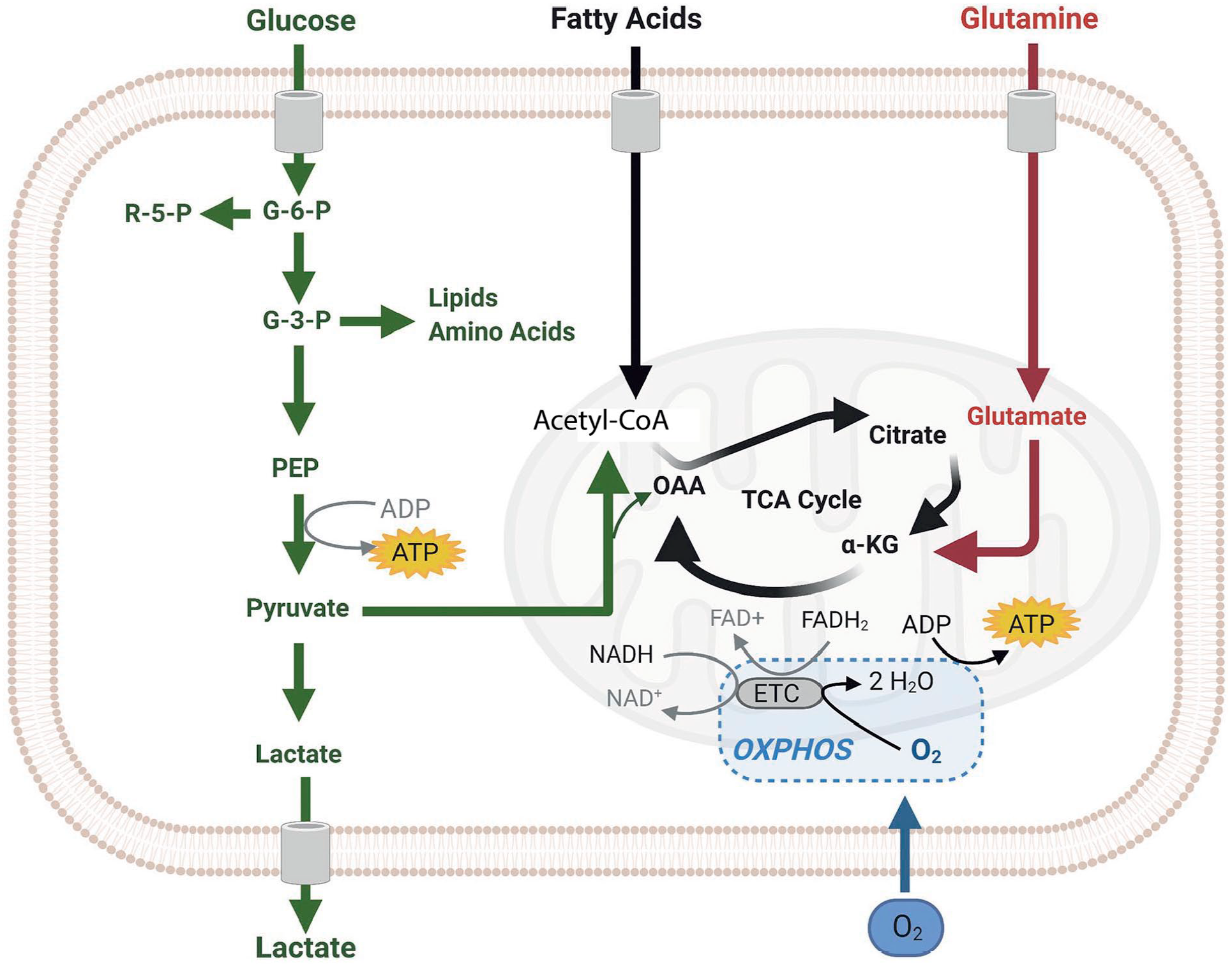
Pathways of cellular ATP synthesis. Simplified scheme of cellular ATP synthesis showing glycolysis and the formation of two molecules ATP and two molecules pyruvate per molecule glucose (green). Pyruvate can then enter mitochondria to become metabolized by oxidative phosphorylation (OXPHOS) using oxygen (O2) to produce 36 ATP molecules per molecule glucose (black). When needed, for example in anaerobic conditions or when lactate and intermediate compounds of the glycolysis pathway are needed for the synthesis of building blocks for macromolecules in proliferating cells, pyruvate is converted into lactate by lactate dehydrogenase A without any further ATP formation (green). Glutamine (red) and fatty acid oxidation (black) are other providers of metabolites to generate ATP in the mitochondria but are not discussed any further in the present article. Image reproduced with permission. 13 Abbreviations: α-KG, α-ketoglutarate; ETC, electron transport chain; FAD+ and FADH2, oxidized and reduced flavin adenine dinucleotide; G-3-P and G-6-P, glucose 3-phosphate and 6-phosphate; NAD+ and NADH, oxidized and reduced nicotinamide adenine dinucleotide; OAA, organic acid oxaloacetate; PEP, phosphoenolpyruvate; R-5-P, ribose 5-phosphate; TCA, tricarboxylic acid cycle.
Cellular Pathways for ATP Production.
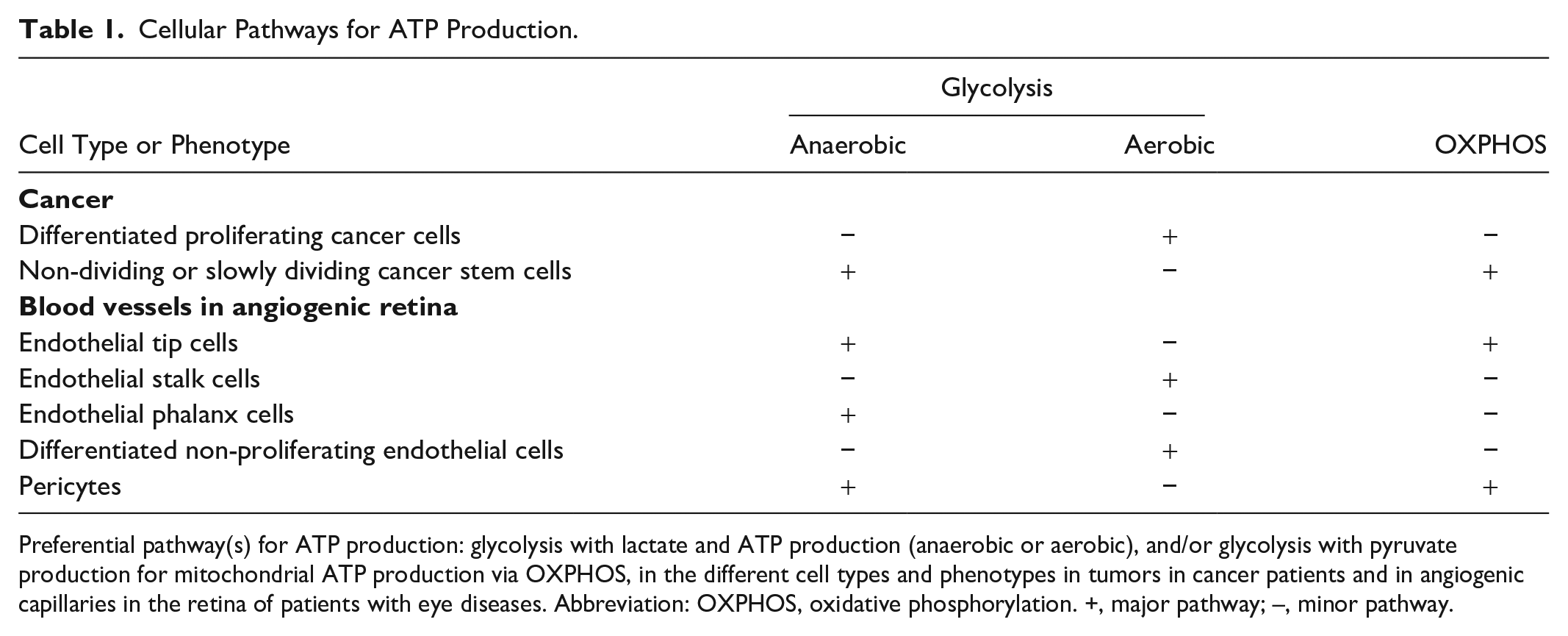
Preferential pathway(s) for ATP production: glycolysis with lactate and ATP production (anaerobic or aerobic), and/or glycolysis with pyruvate production for mitochondrial ATP production via OXPHOS, in the different cell types and phenotypes in tumors in cancer patients and in angiogenic capillaries in the retina of patients with eye diseases. Abbreviation: OXPHOS, oxidative phosphorylation. +, major pathway; –, minor pathway.
The aim of the present review is to link ATP synthesis either via (an)aerobic glycolysis or OXPHOS or both with the various subtypes of cancer cells (proliferating dCCs versus non-proliferating CSCs) and cells active in the angiogenic eye (proliferating stalk ECs versus non-proliferating tip ECs). For this purpose, we have explained the energy (ATP) metabolism of the different cell (sub)types in tumors and the angiogenic eye.
Cellular ATP Synthesis
Figure 1 shows the two major pathways in cells to generate ATP. In the first pathway, a glucose molecule is converted by glycolysis into two pyruvate molecules that are shuttled into mitochondria where pyruvate is converted into acetyl-CoA that is processed by OXPHOS with the use of oxygen to produce ATP. In the second pathway, one glucose molecule is converted by glycolysis into two pyruvate molecules that are then converted into two lactate molecules by lactate dehydrogenase (LDH)A. Glycolysis with subsequent lactate production in normal cells is considered to be a salvation pathway to produce ATP when there is a shortage of oxygen (anaerobic glycolysis), producing only two molecules ATP and two molecules lactate per molecule glucose, whereas pyruvate formation via glycolysis and subsequent transport into mitochondria up to 36 molecules ATP are generated per molecule glucose via OXPHOS in the presence of oxygen.5,13 However, we argue in the next paragraph that glycolysis with lactate production also can occur in normal cells in the presence of sufficient oxygen (aerobic glycolysis). Lactate can be converted into pyruvate, for example in PCs, by LDHB and then channeled into mitochondria.
Production of two molecules ATP per molecule glucose via glycolysis and lactate formation is considerably faster than ATP production via mitochondrial OXPHOS (up to 100× faster).34–37 The higher rate but lower yield of ATP production via glycolysis with lactate production versus the lower rate but higher yield of ATP production via OXPHOS may well have an evolutionary background. 36 Unicellular organisms competing for glucose benefit from fast conversion of glucose to generate ATP, whereas the transition from unicellular to multicellular organisms may have been facilitated by compartmentalized low-speed but high-yield ATP formation by OXPHOS. 36
ATP Synthesis in Cancer Cells
It is still a dogma, formulated by Otto Warburg almost one hundred years ago, that cancer cells produce ATP via glycolysis with lactate formation from pyruvate via LDHA whether oxygen is present or not, the so-called aerobic glycolysis, also known as the Warburg effect.2,5 Part of the dogma was that cancer cells were assumed to suffer from dysfunctional mitochondria.5,38 Aerobic glycolysis was added in 2011 to the famous and much-cited list of “hallmarks of cancer” of Hanahan and Weinberg (Table 2), and was proposed to be an attractive novel target for specific or selective anticancer therapy.1,6,7 However, recent convincing evidence shows that aerobic glycolysis, and thus the Warburg effect, is not a specific hallmark of cancer. The evidence is listed here and summarized in Table 3. Despite this convincing evidence, over 2000 articles have been published in the past decade on the Warburg effect as a hallmark of cancer without a critical analysis whether this assumption is correct. 39
Hallmarks of Cancer.
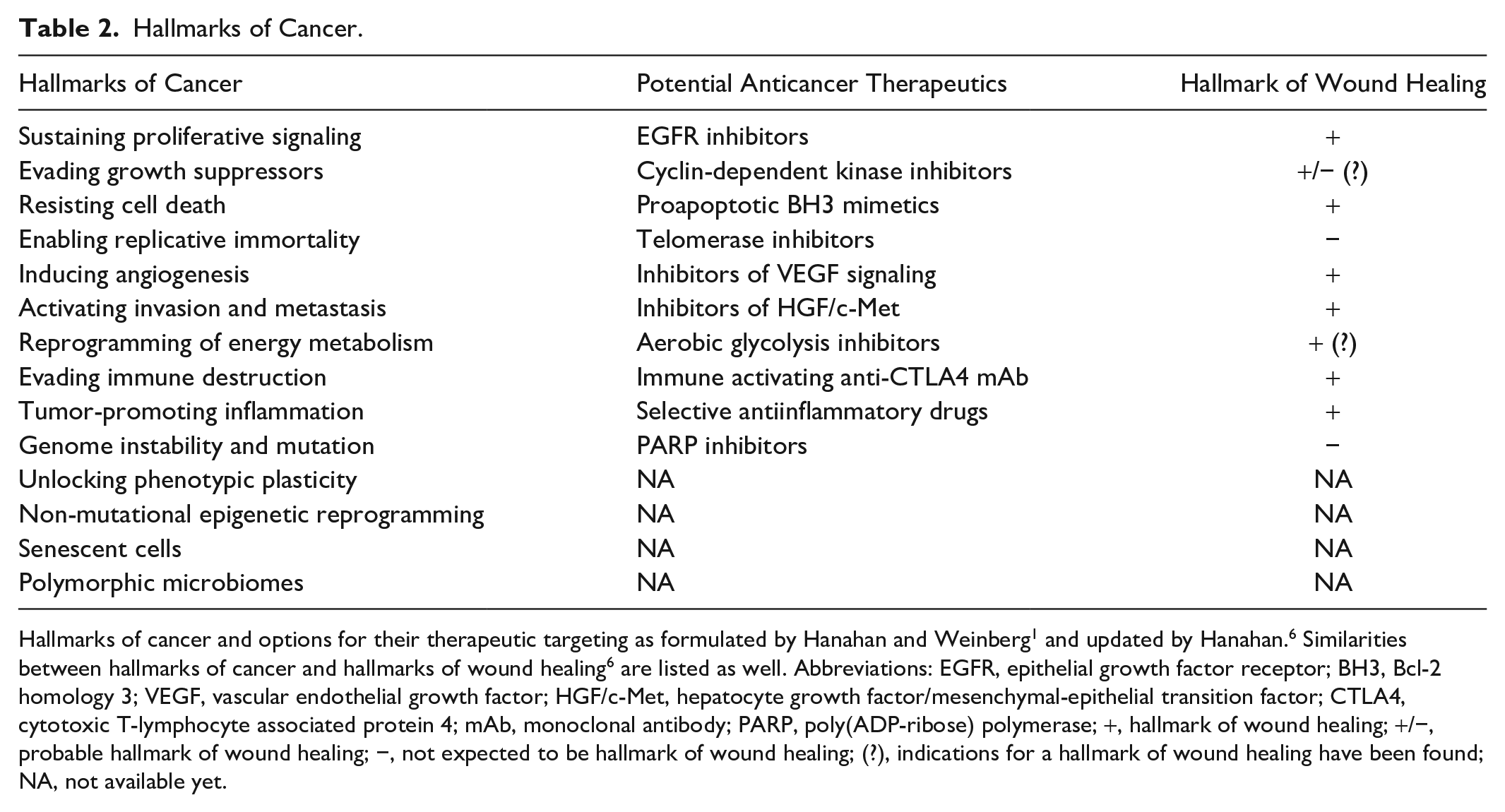
Hallmarks of cancer and options for their therapeutic targeting as formulated by Hanahan and Weinberg 1 and updated by Hanahan. 6 Similarities between hallmarks of cancer and hallmarks of wound healing 6 are listed as well. Abbreviations: EGFR, epithelial growth factor receptor; BH3, Bcl-2 homology 3; VEGF, vascular endothelial growth factor; HGF/c-Met, hepatocyte growth factor/mesenchymal-epithelial transition factor; CTLA4, cytotoxic T-lymphocyte associated protein 4; mAb, monoclonal antibody; PARP, poly(ADP-ribose) polymerase; +, hallmark of wound healing; +/−, probable hallmark of wound healing; −, not expected to be hallmark of wound healing; (?), indications for a hallmark of wound healing have been found; NA, not available yet.
Evidence That Aerobic Glycolysis Is Not a Hallmark of Cancer.

Abbreviation: OXPHOS, oxidative phosphorylation.
First, the success of targeting aerobic glycolysis as anticancer therapy in patients has been limited thus far.11,13–19 Second, cancer cells do not necessarily show impaired mitochondrial function.12,13,40 In fact, functional mitochondria have been shown recently to be transferred from non-cancer astrocytes to primary brain tumor (glioblastoma) cells via tunneling nanotubes (TNTs) that upregulate OXPHOS activity in the cancer cells. 41 Third, aerobic glycolysis is not only found in rapidly proliferating cancer cells but also in rapidly proliferating healthy cells.4,42,43,44 Both proliferating non-cancer cells and cancer cells need the synthesis of building blocks for cell growth which is facilitated by products of glycolysis, such as lactate, 4 and NADPH for biosynthetic reactions.5,12,13,35,38,45 Fourth, normal quiescent differentiated cell types such as erythrocytes, 46 fibroblasts, 45 ECs, 20 and astrocytes in the central nervous system (CNS)13,47–49 also use aerobic glycolysis for ATP and lactate synthesis. Excess lactate produced in ECs and astrocytes is exported to the extracellular matrix (ECM) and imported by PCs and neurons, respectively, to either convert it into pyruvate in the cytoplasm via LDHB that is then channeled into mitochondria or to be directly transferred into mitochondria via transporter proteins in their membranes for its use in OXPHOS.3,31,50,51 Fifth, embryonic development is also associated with the Warburg effect.52–56 Sixth, tumors are heterogeneous and consist of a tumor microenvironment of non-cancer cells, immune cells, blood vessels, and ECM besides a heterogeneous population of cancer cells, such as non-proliferating or slowly proliferating CSCs and rapidly proliferating dCCs, as is shown in a human glioblastoma tumor in Fig. 2.
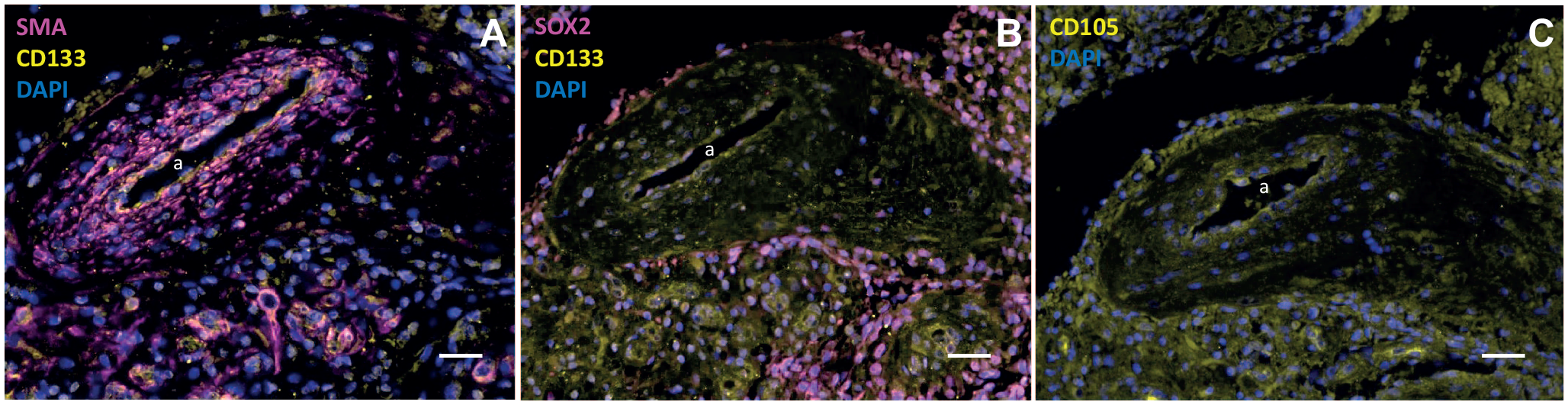
Cancer stem cell niche around an arteriole in a glioblastoma tumor. Immunofluorescence images of paraffin sections of a human glioblastoma tumor showing the heterogeneity of tumor tissue surrounding an arteriole (with the lumen [a] and the thick tunica media around the lumen of smooth muscle actin [SMA]-positive smooth muscle cells) that represents a cancer stem cell niche. The niche contains cluster of differentiation-133 (CD133)- and SRY-box transcription factor 2-positive cancer stem cells, CD105-positive mesenchymal stem cells, and many other undefined cells showing only 4′,6-diamidino-2-phenylindole (DAPI)-positive nuclei. Bars A–C = 100 µm. Immunofluorescence staining and imaging procedures are described in Hira et al.57–59
The ultrastructure of glioblastoma dCCs shows swollen and electron-lucent mitochondria with reduced disorganized cristae60,61 that most likely do not have a functional electron transport chain for OXPHOS activity to produce ATP which is in line with aerobic glycolysis of the Warburg effect. In these studies, tissue samples obtained from patient biopsies were analyzed and thus the bulk tumor dCCs and not the infrequently occurring CSCs in their niches57,58 were studied. We have ultrastructurally compared mitochondria in cultured glioblastoma CSCs and dCCs 13 and observed disrupted swollen electron-lucent mitochondria with disorganized cristae in dCCs and seemingly intact elongated electron-dense mitochondria with highly organized cristae along the longitudinal axis of the mitochondria in CSCs (Fig. 3). These observations are consistent with the concept that dCCs are aerobically glycolytic, whereas CSCs preferentially use OXPHOS for ATP production.12,13,62 At first glance, this is odd because CSCs, like normal stem cells, are found in hypoxic niches, as hypoxic conditions are needed for their stem cell fate and quiescence as has been demonstrated in the subventricular zone in the brain of glioblastoma patients (Fig. 4).63,59 However, quiescent CSCs need only low amounts of ATP that can be synthesized by OXPHOS at low oxygen levels. 12 Furthermore, CSCs use mitochondrial fatty acid oxidation (FAO) that is essential to maintain their stemness2,11–13, which is another indication that mitochondria in CSCs are functional.
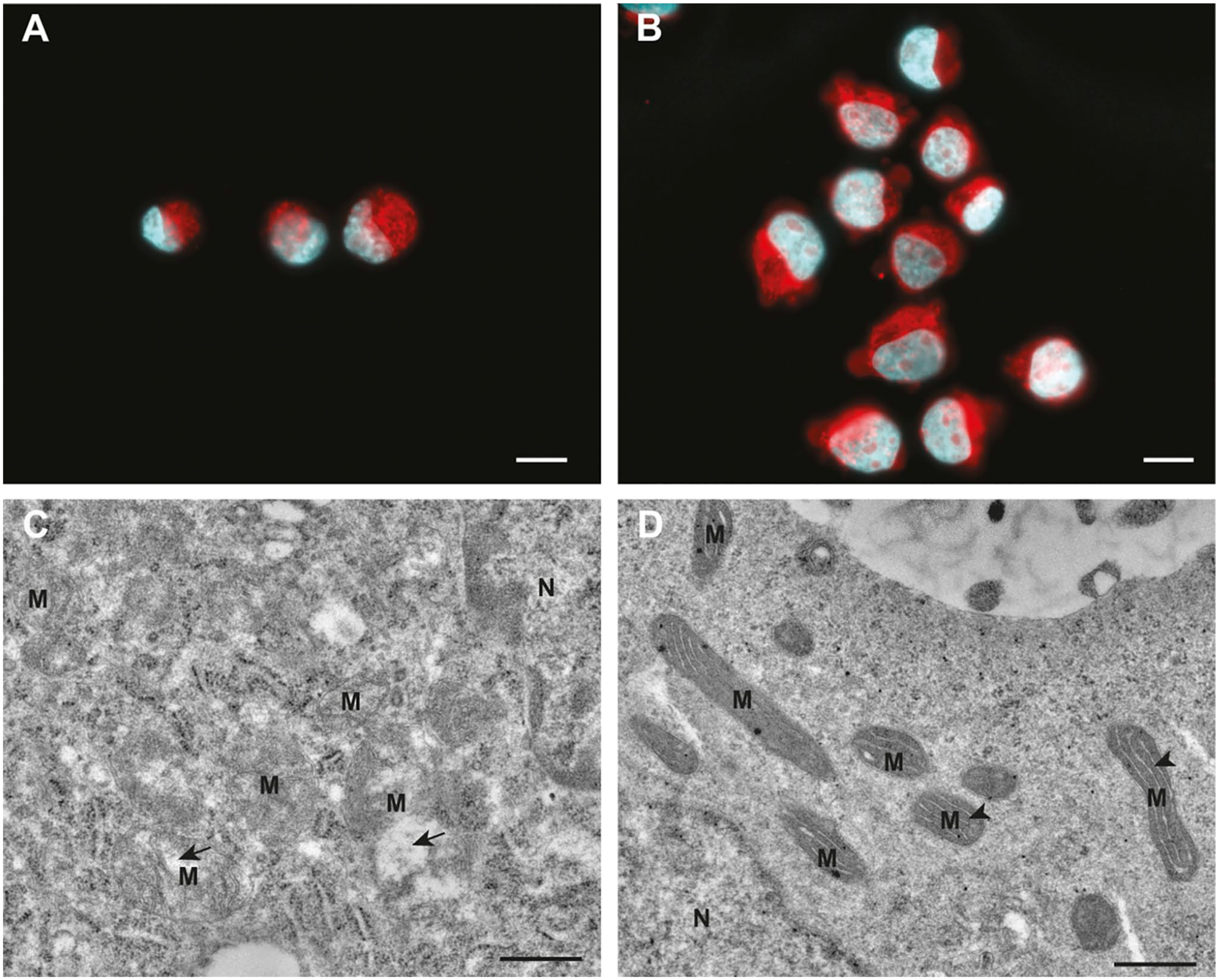
Mitochondria (M) in glioblastoma cancer cells. Differentiated glioblastoma cells (A) and glioblastoma stem cells (B) contain similar large amounts of mitochondrial material as shown here with MitoTracker Red CMXRos labeling of M according to the manufacturer’s instructions (ThermoFisher, Bleiswijk, The Netherlands). However, ultrastructurally M in differentiated glioblastoma cells (C) appear to be swollen, are electron lucent, display disorganized cristae, and often contain distended electron-lucent spaces (arrows). In glioblastoma stem cells (D), the M are elongated, very electron dense, and display highly organized cristae (arrowheads) that are often oriented along the longitudinal axis of M. Cell nuclei in (A) and (B) are stained with 4′,6-diamidino-2-phenylindole (blue). For transmission electron microscopy, cells were fixed in 2.5% glutaraldehyde and 2% formaldehyde followed by postfixation in 1% osmium tetroxide. Cells were embedded in 2% low melting point agarose. The blocks of agarose with embedded cells were solidified on ice and cut into small pieces (<1 mm in smallest dimension). The pieces of agarose with cells were dehydrated in graded series of ethanol and acetone. Finally, samples were embedded in epoxy resin Agar 100. After polymerization for 24 hr at 60C in embedding molds, ultrathin sections were prepared with Reichert Ultracut S (Leica, Amsterdam, The Netherlands) ultramicrotome, using a diamond knife. The ultrathin sections were contrasted with uranyl acetate and lead citrate and imaged with CM100 (Philips, Eindhoven, The Netherlands) transmission electron microscope. The electron micrographs were acquired with Orius 200 (Gatan) camera and Digital Micrograph (Gatan, Pleasanton, CA, USA) software. Bars A and B = 10 µm; bars C and D = 0.5 µm.
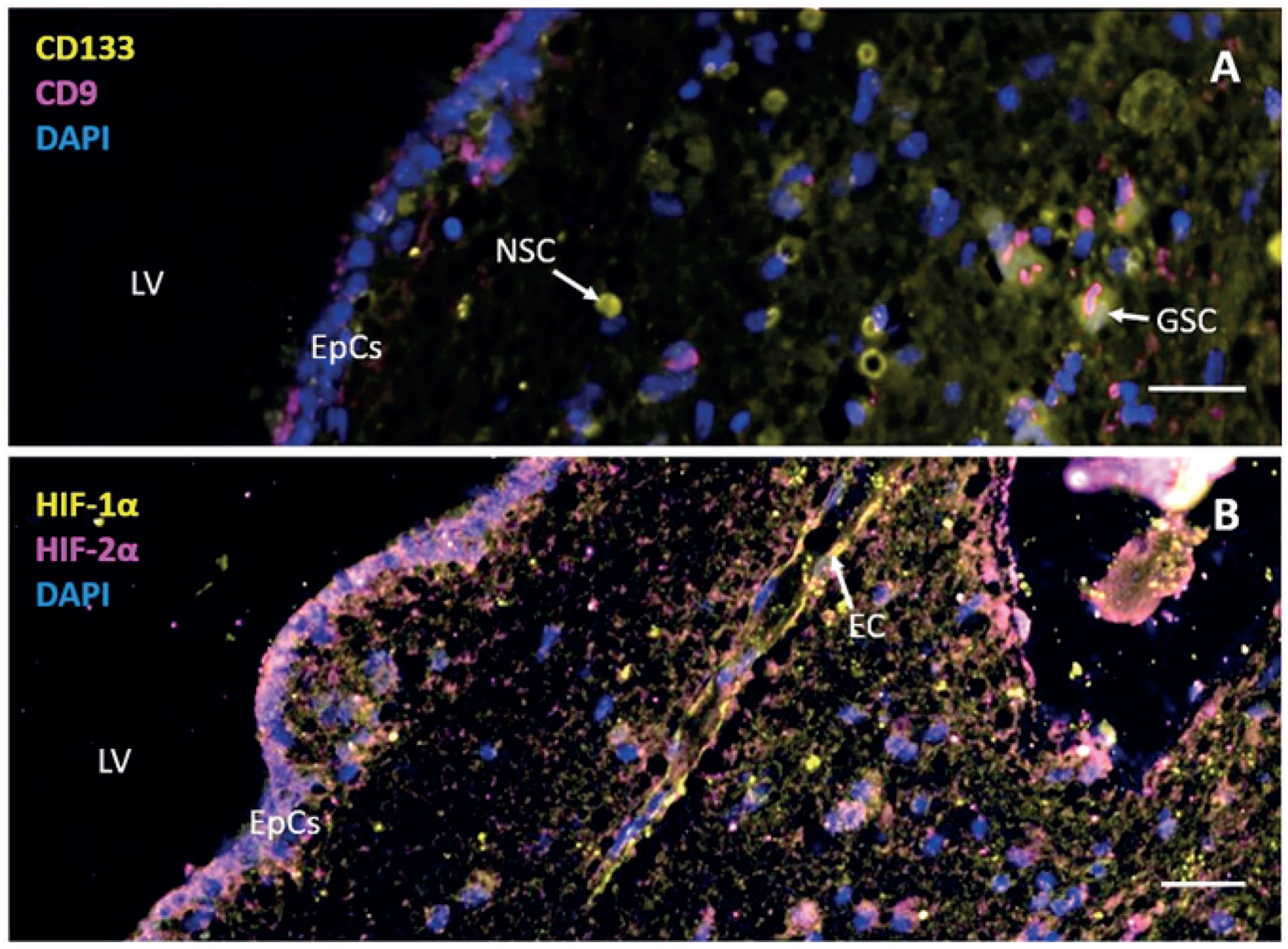
Neural stem cells and glioblastoma cancer stem cells in the subventricular zone of a brain tumor patient. Immunofluorescence images of paraffin sections of the subventricular zone of the postmortem brain of a glioblastoma patient after surgery, radiotherapy, and temozolomide chemotherapy showing (A) the presence of cluster of differentiation-133 (CD133)+ and CD9− neural stem cells (NSCs) and CD133+ and CD9+ glioblastoma stem cells (GSCs) and (B) staining patterns of hypoxia-inducible factor (HIF)1α and HIF2α. 4′,6-Diamidino-2-phenylindole (DAPI) is used for counterstaining of cell nuclei. Bars A and B = 100 µm. Abbreviations: EpCs, ependymal cells; EC, endothelial cell; LV, lateral ventricle. Immunofluorescence staining and imaging procedures are described in Hira et al. 59
This overwhelming evidence obtained in recent years that aerobic glycolysis or the Warburg effect is not a specific hallmark of cancer makes this dogma obsolete and ready for the biochemical museum. 51
A more accurate concept is that ATP synthesis via glycolysis and lactate formation and/or mitochondrial OXPHOS is regulated as a response to the microenvironment of both normal cells and cancer cells.8,15,41 As a consequence, it is reasonable to argue that ATP synthesis via aerobic glycolysis is not a realistic target for systemic anticancer therapy and the lack of success in clinical trials is in line with that.11,16–19 We have postulated that systemic treatment of cancer patients with OXPHOS inhibitors is not a realistic option either. 13 OXPHOS is necessary to produce low levels of reactive oxygen species (ROS) needed for the proper folding of proteins in the endoplasmic reticulum as was argued by Watson, 64 and OXPHOS-dependent exercise is beneficial for the quality-of-life of cancer patients.13,50,51,65,66,67 The necessary low cellular levels of ROS are generated by OXPHOS from 0.1% to 2% of the electrons that escape from the electron transport chain. 13
In conclusion, rapidly proliferating dCCs are preferentially glycolytic, whereas CSCs preferentially use OXPHOS for ATP production as an energy source, and inhibition of ATP production via either pathway is not a therapeutic option in cancer patients. Whether ATP synthesis is an attractive target in angiogenic ocular diseases is discussed in the following paragraphs.
Physiological and Pathological Angiogenesis
The formation of the vasculature in humans is completed during embryonic development and the vasculature remains mostly quiescent throughout the entire life span. Only under specific circumstances, such as an hypoxia-driven vascular endothelial growth factor (VEGF) gradient, ECs can switch to an activated state to form new blood vessels by sprouting from existing blood vessels, which can be simulated in vitro (Fig. 5). This post-embryonic formation of new vessels is called angiogenesis and occurs in the ovaries and endometrium at the beginning of each menstrual cycle and in wound healing processes.25,69–71 Angiogenesis is tightly regulated by pro-angiogenic and anti-angiogenic factors.68,72–75 Besides these physiological processes, angiogenesis can also occur under pathological circumstances, for example in tumors1,76,77 and in neovascular ocular diseases, such as proliferative diabetic retinopathy, exudative age-related macular degeneration, and retinopathy of the prematurity.78–84 Both in tumors and in these eye conditions, angiogenesis usually does not occur as a single process, but invariably drives at least partly elements of the wound healing cascade, such as fibrin deposition, influx of immune cells, formation of myofibroblasts, and deposition of matrix components.84–90 The growth of primary tumors and secondary metastases requires angiogenesis when the tumors reach a volume of approximately 1–2 mm3 and diffusion of oxygen from existing blood vessels is not sufficient anymore.91–94 Shortage of oxygen (hypoxia) occurs in cancer cells and non-cancer cells in the center of the tumor inducing expression of hypoxia-inducible factors (HIFs). HIF1 and HIF2 are the most relevant transcription factors that switch on the transcription of a large number of genes. To rescue hypoxic cells, HIF1 regulates the expression of 701 genes and HIF2 regulates that of 1445 genes. 95 The third member of the HIF family, HIF3, has a multitude of splice variants with yet unknown functions and may be involved as well in the rescue of cells from hypoxia. 96
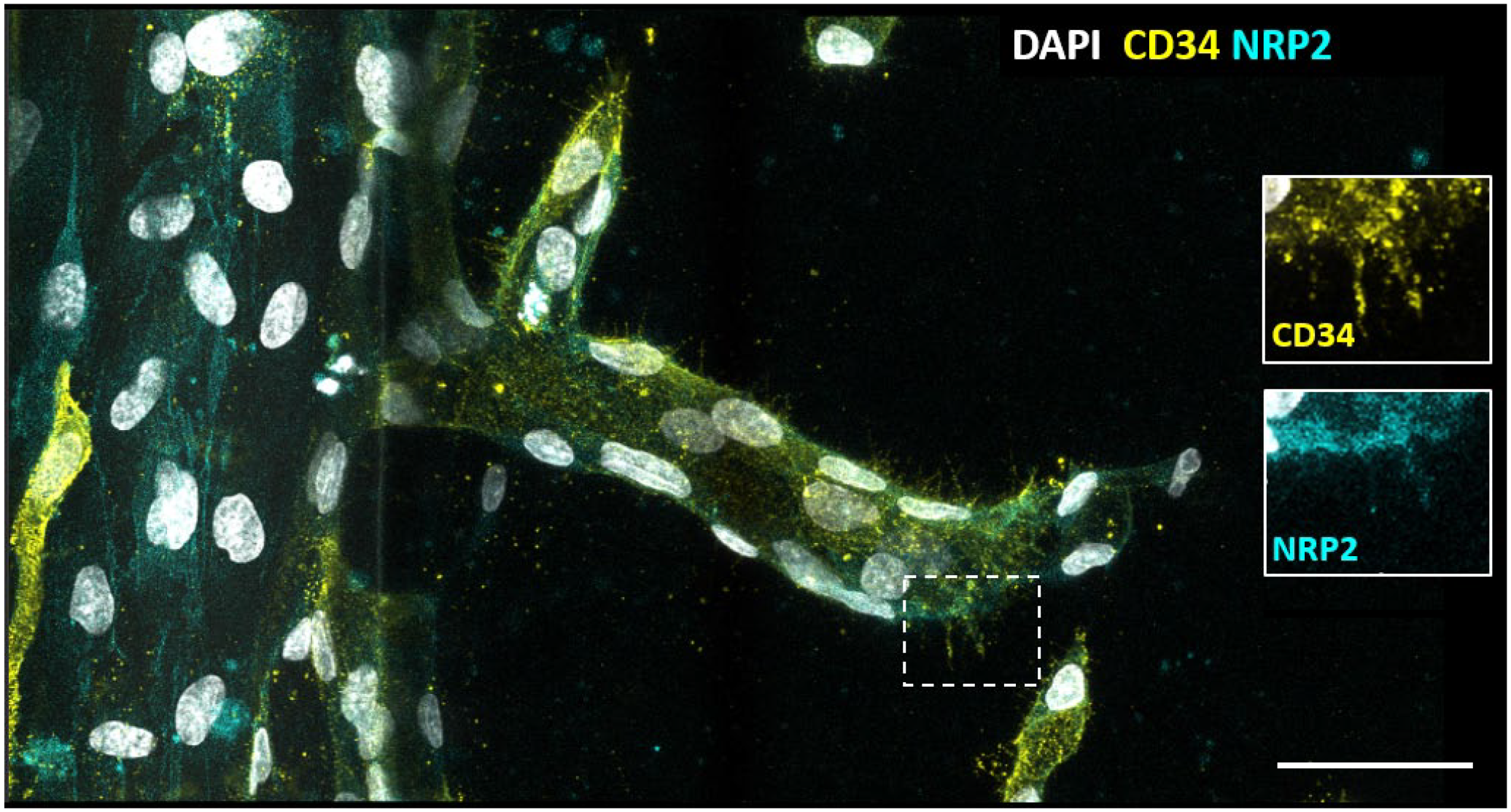
In vitro outgrowth of an angiogenic capillary sprout from an existing capillary induced by vascular endothelial growth factor (VEGF). Sprouting of human umbilical vein endothelial cells in a 3-lane OrganoPlate, a 3D vessel-on-a-chip sprouting assay on a gradient of angiogenic factors VEGF and sphingosine-1-phosphate. A luminized sprout is growing from the capillary on the left toward the right, on a substrate of collagen type 1. Immunofluorescence staining is shown of cluster of differentiation-34 (CD34), an in vitro tip endothelial cell (EC) marker (yellow) and neuropilin 2 (NRP2) as a tip EC-specific marker (blue) and 4′,6-diamidino-2-phenylindole (DAPI) as nuclear staining (white). Inserts show specific staining of CD34 on filopodia. In vitro capillary sprouting, immunofluorescence staining, and imaging are described in Dallinga et al. 68 Bar = 20 µm.
HIF1 and HIF2 are both inducers of the expression of VEGF-A which is a major promoter of angiogenesis.91,92,97–99 Unlike in physiological angiogenesis, ECs of the vasculature in tumors and retina keep proliferating and fail to become quiescent under pathological conditions, leading to continuous growth of tumors and extensive wound healing responses in the eye conditions, respectively.84,94,98,99 During these processes, the new vessels do not mature in time, and remain leaky and prone to hemorrhage which are serious clinical complications of pathological angiogenesis. Consequently, angiogenesis in tumors and neovascular ocular diseases has become the topic of intensive research to develop novel therapeutic anti-angiogenesis approaches.71,93,98,100 Anti-angiogenesis therapy in the eye using antibodies against VEGF-A is reasonably effective, but may induce fibrosis in the eye due to an imbalanced ratio of VEGF-A and connective tissue growth factor 88 and the necessary monthly treatment is a considerable burden for patients. 101 The success of this anti-angiogenesis approach in cancer including clinical trials has been limited thus far.24,25
Treatment of pathological angiogenesis in the retina can be performed locally, for example by intravitreal injections or eye drops, thus avoiding side effects that may occur during systemic treatment of patients such as cancer patients. Nanocarriers may become a useful tool to deliver drugs intra-ocularly with limited invasive technology to minimize the burden for patients. 102 We discuss in the next chapters whether local administration in the eye of inhibitors of ATP synthesis via glycolysis and/or OXPHOS may be a realistic therapeutic approach.
Endothelial Cell Differentiation During Angiogenesis: Tip Cell, Stalk Cell, and Phalanx Cell Phenotypes
When angiogenesis is induced, quiescent mature ECs of existing blood vessels transdifferentiate into a heterogeneous activated population of ECs to form new blood vessel sprouts. The sprouts consist of at least three specialized EC phenotypes (Fig. 5). The leading cell on top of a sprout is a tip EC that possesses probing filopodia. This single tip EC is followed by non-tip ECs, which are rapidly proliferating stalk ECs which differentiate into non-proliferating maturing phalanx ECs (Fig. 5).10,25,72,73,103–105
First, a single tip EC grows out from a pre-existing vessel, migrates into the ECM, and guides the developing sprout consisting of stalk ECs and phalanx ECs along the gradient of pro-angiogenesis factors, such as VEGF-A.28,106 Second, the highly proliferative stalk ECs follow the leading tip EC, form the vessel lumen, and ensure vessel elongation. Third, proliferating stalk ECs become maturing non-proliferating phalanx ECs that tightly align and organize themselves in, fourth, a continuous monolayer of quiescent mature ECs around the lumen.30,105 Simultaneously, tip ECs of two sprouts originating from different existing vessels can merge (anastomosis) to form a new vascular loop.72,73,103,104
Endothelial Tip Cells, Stalk Cells, and Phalanx Cells as Therapeutic Targets in Pathological Angiogenesis
Tip ECs are an attractive target for anti-angiogenesis therapy, as they initiate the angiogenesis process as leader ECs of newly formed blood vessel sprouts by assimilating directional cues from their microenvironment. Tip ECs have a unique molecular and functional signature and quite a large number of growth factors, cytokines, and other signaling molecules play a role in regulating their functions.72,107 Besides, animal models and in vitro models have shown the involvement of metabolic pathways in the regulation of angiogenesis. However, the specific metabolic pathways of tip ECs and their regulation had not been studied until recently because specific or selective biomarkers for tip ECs were not available.
We have identified a selective biomarker for tip ECs, cluster of differentiation-34 (CD34), to enable discrimination of CD34-positive (CD34+) tip ECs from CD34-negative (CD34−) non-tip ECs in vitro.73,74,108 CD34+ tip ECs in vitro show similar characteristics and are regulated by similar mechanisms as tip ECs in vivo (Fig. 6).73,74,108 Afterward, we have studied energy metabolism in tip ECs and non-tip ECs in vitro.28,29
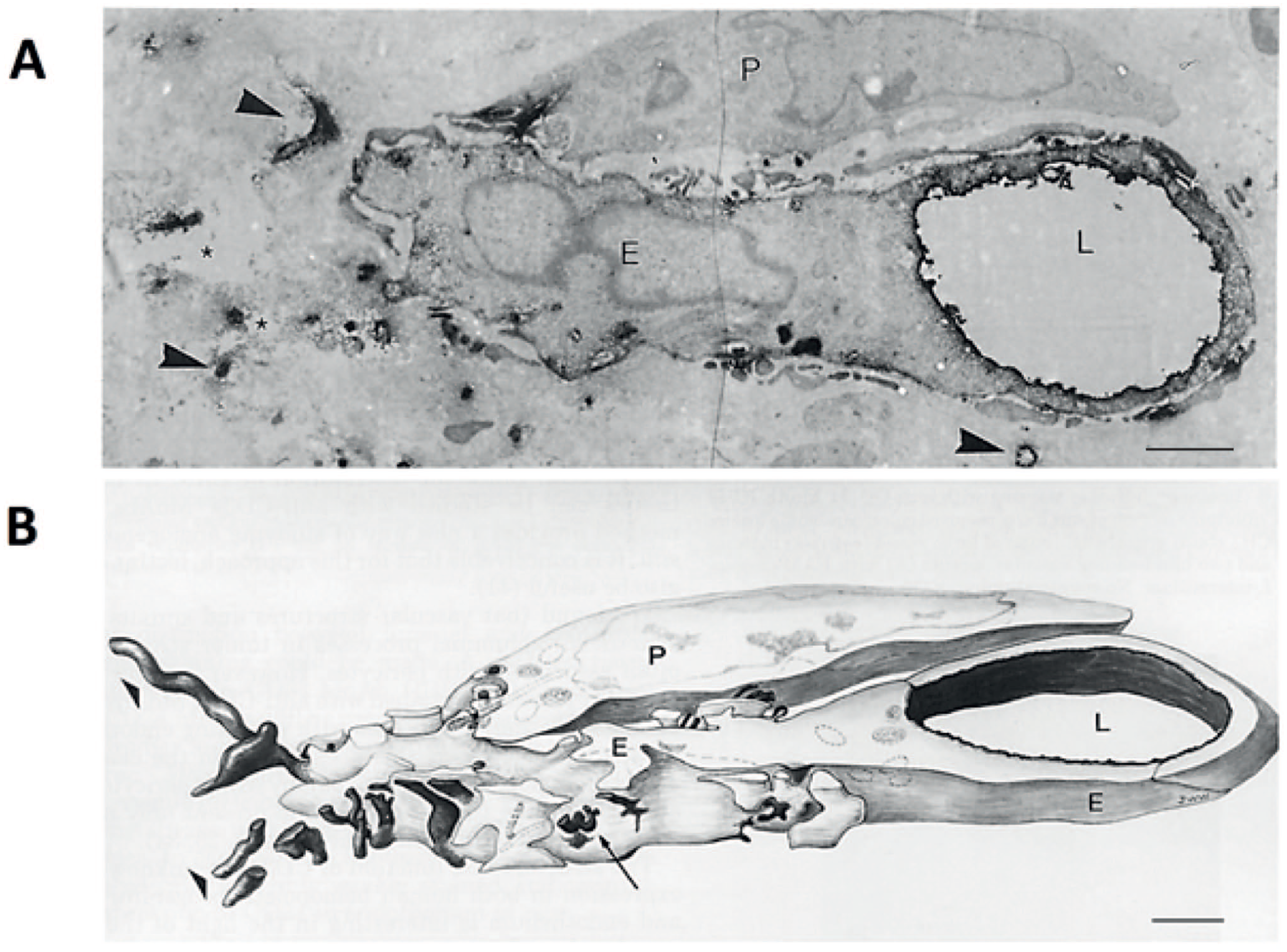
Electron microscopical image and a 3D reconstruction of a cluster of differentiation-34 (CD34)-expressing endothelial cell in a colon cancer biopsy. Electron microscopy image (A) of a 50-µm-thick ultrathin section of after immunohistochemical peroxidase activity staining of CD34 localized with an anti-CD34 antibody (black precipitations) and (B) 3D reconstruction prepared in 1990 (!) of 10 serial ultrathin sections of the same vascular sprout. CD34 positivity is found at the luminal side of an endothelial tip cell with CD34-positive abluminal filopodial extrusions (arrowheads, arrows and asterisks). Bars A and B = 1 µm. Abbreviations: P, pericyte; E, endothelial tip cell; L, lumen of capillary. Image reproduced with permission of the United States and Canadian Academy of Pathology. 109
CD34 is a member of the CD34 transmembrane phosphoglycoproteins family together with endoglycan and podocalyxin.110,111 CD34 is expressed on stem cells and progenitor cells as well as CSCs and enhances cell proliferation and inhibits cell differentiation. CD34 stimulates cell migration and can promote and block cell adhesion.110,111 CD34 is a biomarker of EC precursor cells and of tip ECs in vitro, whereas in vivo it is expressed on the apical membrane of ECs during angiogenesis and plays an anti-adhesive role in lumen formation and possibly in maintaining lumina of capillaries (Fig. 6).73,112 Both anti-adhesive and adhesive properties of CD34 may also facilitate migration of tip cells, particularly of the probing filipodia. 73 The exact roles of CD34 in (cancer) stem cells, progenitor cells, and ECs are still far from understood. 111
In the next section of this review, we evaluate energy metabolism in tip ECs and non-tip ECs (proliferating stalk ECs, maturing phalanx ECs, and quiescent mature ECs), as well as that of PCs as a potential target for anti-angiogenic therapy in ocular diseases.
ATP Synthesis in Tip and Non-Tip Endothelial Cells
Recently, we have reviewed potential anti-angiogenesis therapies in gynecological cancers in parallel with our studies on energy metabolism in tip ECs and non-tip ECs, during vascular sprouting. 25 The similarity between ATP synthesis in tip ECs and non-tip ECs and that in CSCs and dCCs, respectively, was striking. Therefore, we hypothesize that both the non- or slowly proliferating tip cells and CSCs preferentially use OXPHOS for ATP synthesis, whereas proliferating stalk ECs and progenitor cancer cells as well as maturing phalanx ECs and mature ECs and dCCs are preferentially glycolytic. These three EC and cancer cell subtypes are comparable with stem cells, progenitor cells, and differentiated cells in normal tissues such as bone marrow, and intestinal and skin epithelium. Our metabolic studies of purified CD34+ tip ECs and CD34− non-tip ECs in cultures of ECs of different origin, including human umbilical vein ECs, showed that both EC phenotypes are metabolically flexible and are able to use both glycolysis and OXPHOS, but non-tip ECs tend to rely relatively more on glycolysis and less on OXPHOS for ATP production compared with tip ECs.28,29
The transcription factor forkhead box O transcription factor 1 (FOXO1) is highly expressed in quiescent differentiated ECs of capillaries and is downregulated during angiogenesis by the phosphoinositide 3-kinase (PI(3)K)/protein kinase B (AKT) pathway that links cell growth and metabolism. 13 PI(3)K signaling inhibits FOXOs through AKT-mediated phosphorylation that causes nuclear exclusion of FOXOs. 113 FOXO1 suppresses c-MYC signaling that drives energy metabolism, both glycolysis and OXPHOS, and cell proliferation.10,13 It reduces energy metabolism and ROS production in quiescent differentiated ECs of capillaries to a minimum despite the high oxygen levels in their lumen, to optimize their function to deliver oxygen and nutrients to surrounding tissues. 20 During energy-demanding angiogenesis, FOXO1 expression is downregulated to enable the upregulation of energy metabolism and EC proliferation.13,113 Furthermore, downregulation of FOXO1 expression enhances glycolysis-dependent DNA repair. 114 The downregulation of FOXO1 expression in both tip ECs and non-tip ECs has not yet been investigated. However, we hypothesize that FOXO1 expression is relatively less reduced in tip ECs compared with non-tip ECs. This hypothesis is supported by our finding that FOXO1 mRNA levels in tip ECs in vitro are higher than those in non-tip ECs (Miyamura et al., 2024). This observation aligns with the concept that tip ECs exhibit lower proliferation and reduced reliance on glycolysis compared with non-tip ECs (Table 4). Furthermore, after both FOXO1 knockdown and under hypoxic conditions, we observed a relative increase in the number of tip cells (Miyamura et al., 2024). This phenomenon may be explained by a reduced fraction of proliferating non-tip ECs which is in agreement with the recently published article by Miyamura et al. 115 that reported that FOXO1 stimulates the tip ECs.
Glycolysis and OXPHOS in Different Cell Types.
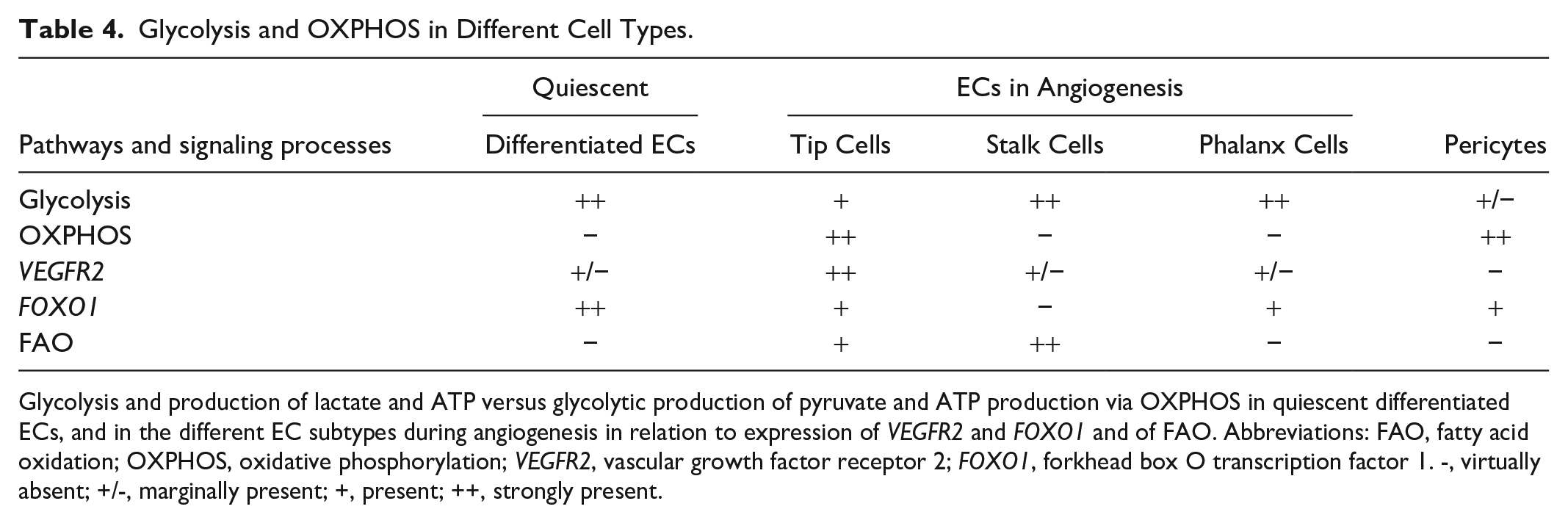
Glycolysis and production of lactate and ATP versus glycolytic production of pyruvate and ATP production via OXPHOS in quiescent differentiated ECs, and in the different EC subtypes during angiogenesis in relation to expression of VEGFR2 and FOXO1 and of FAO. Abbreviations: FAO, fatty acid oxidation; OXPHOS, oxidative phosphorylation; VEGFR2, vascular growth factor receptor 2; FOXO1, forkhead box O transcription factor 1. -, virtually absent; +/-, marginally present; +, present; ++, strongly present.
How tip ECs generate ATP needed for migration is not clear yet. It may well be that tip ECs use OXPHOS, given their high OXPHOS capacity. 116 Alternatively, tip ECs may switch between OXPHOS and anaerobic glycolysis during migration as tip ECs in vitro have shown to be flexible in this aspect to be able to adapt to the local availability of nutrients and oxygen while migrating. This suggests that tip ECs are well equipped to respond to microenvironmental circumstances during migration. 28 De Bock et al. 21 and Rivera and Bergers 117 in a commentary conclude that ECs are glycolytic during angiogenesis without discriminating between tip ECs and stalk ECs. Furthermore, Schoors et al. 118 demonstrated that FAO is essential for DNA synthesis and not for energy supply for migration via OXPHOS in ECs during angiogenesis. In a commentary, Egnatchik and Deberardinis speculated that there must be two different metabolic pools in angiogenic ECs, one for cell proliferation and one for cell migration. 119 However, in these studies tip ECs and non-tip ECs were not separately investigated and because tip ECs are very much in the minority in EC cultures, the metabolic identity of tip ECs was most likely masked in these studies. Based on our studies on the metabolism of purified tip ECs and non-tip ECs in vitro, we conclude that tip ECs use OXPHOS relatively more and aerobic glycolysis relatively less than non-tip ECs for ATP production.28,29 In stalk ECs, FAO appeared to be essential for nucleotide synthesis needed for DNA replication and not for migration. 117 Mechanical regulation of cellular metabolism in response to extracellular environmental cues8,15,28,41 with the cytoskeleton as an intermediate may well be involved, in particular in migrating tip ECs with their flexible metabolism.8,28
In conclusion, during angiogenesis, proliferating non-tip ECs are relatively more glycolytic and rely less on OXPHOS for ATP production and biosynthetic processes, as compared with migrating tip ECs and use FAO for DNA synthesis. Differentiating maturing phalanx ECs are the population of ECs in-between rapidly proliferating stalk ECs and quiescent mature ECs and are also preferentially glycolytic.9,10,21,72,105
Pericytes as Therapeutic Targets in Pathological Angiogenesis
PCs preferentially use OXPHOS for ATP synthesis (Table 4). PCs of the CNS originate from the neuronal crest and interact with ECs as local stromal cells.30,120,121 PCs are present in the wall of small blood vessels but not in lymphatic capillaries. 30 PCs and ECs share a basal lamina (Fig. 7A and B) and have discrete contact points through holes in the basal lamina. Up to 1000 contact points per EC have been found.120–122 These contact points are of the peg–socket type, with PC fingers (pegs) inserted in EC invaginations (sockets; Fig. 7E to G). EC pegs in PC sockets also exist, but occur less frequently. 122 These subcellular structures function as gap junctions enabling the exchange of molecules and charge.30,122–124 Furthermore, PCs and ECs have contacts that function as adhesion plaques. 30 In diabetic patients, these PC–EC contacts are disrupted. 125 In most capillaries in the body, the PC coverage of ECs is in a ratio of approximately 1:100. However, in capillaries that form the blood–brain barrier (BBB) and blood–retina barrier (BRB), the EC:PC ratio is very high, 1:1 to 3:1 with a 30–99% coverage of the apical surface of ECs.30,126
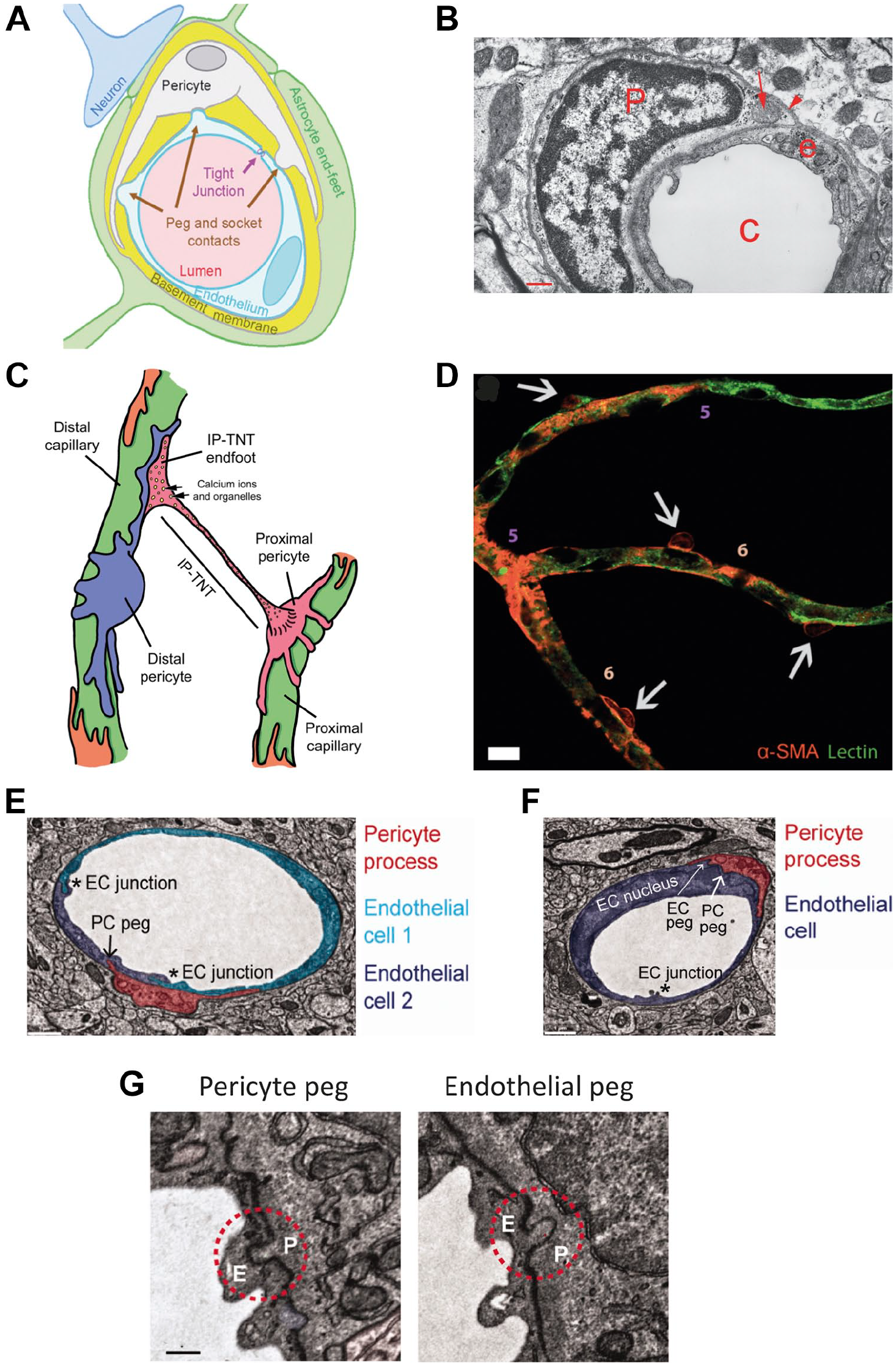
Structural interactions between endothelial cells (ECs) and pericytes (PCs). (A) Schematic drawing of a PC (white) encapsulating an EC (light blue) surrounded and separated by a basal lamina (basement membrane) around the capillary lumen (pink). Tight junctions and peg-socket contacts are indicated. (B) Electron microscopical image of a PC (P) encapsulating an EC (e) around a capillary lumen (C). The basal lamina is indicated by an arrowhead and a mitochondrion in the PC cytoplasm by an arrow. (C) Schematic drawing of PCs encapsulating two capillaries with an interpericyte tunneling nanotube (IP-TNT) connecting two PCs for IP communication. (D) Fifth- and sixth-order capillaries in vitro with ECs (green) encapsulated by PCs (orange). (E-G) Electron microscopical images of PCs encapsulating ECs, showing tight junctions between ECs (*) and peg-socket contacts between ECs and PCs. Bar B = 0.5 µm; bar D = 10 µm; bars E–G = 3 µm. Abbreviation: α-SMA, α-smooth muscle actin. (A)-(D) are reproduced with permission under a Creative Commons CC-BY International License. 121 (E)-(G) are reproduced with permission under the 2022 STM Guidelines (https://www.stm-assoc.org/) of which SAGE is a signatory member. 122
PCs modulate vessel diameter, blood flow, vessel permeability and regulate BBB and BRB functions.121,127 Moreover, PCs regulate EC proliferation and differentiation during angiogenesis and stabilize newly formed EC tubes. In the past, it was assumed that PCs surround only the quiescent phalanx ECs during angiogenesis to participate in the stabilization of the new vessel wall, but it is now clear that PCs are also present around tip ECs and stalk ECs.109,128–131 Platelet-derived growth factor (PDGF)-B-PDGF receptor(R)-β signaling plays a role in the recruitment of PCs by ECs for vessel maturation and stabilization. PDGF-B is released by ECs and binds to PDGFRβ on the membrane of differentiating PCs. Tip ECs express higher levels of PDGF-B than stalk ECs and thus may attract PCs to the front of sprouting EC tubes.30,120,109 Other signaling pathways that are involved in PC recruitment to ECs are angiopoietin 1 (Ang-1)-tyrosine-protein kinase receptor 2 (Tie)-2 and stromal-derived factor 1 (CXCL12)-C-X-C chemokine receptor type 4 (CXCR4). Ang-1 is expressed by PCs and its receptor Tie-2 is expressed by ECs and their binding mediates the maturation and stability of the vessel sprout to reduce vascular leakage.132,133 CXCL12 expressed on ECs binds CXCR4 on PCs and is involved as well in the maturation of the vessel sprout and assembly of the basal lamina.134,135 On the other hand, CXCR4 is also enriched in tip cells73,136 and may well be necessary during migration by binding to CXCL12.
PC neurogenic locus notch homolog protein (NOTCH)3 and EC NOTCH1 are necessary for vessel stabilization. Delta-like ligand 4 expression in PCs is NOTCH3 dependent and it is a ligand for NOTCH1 in ECs. 137 In addition, CNS PCs are also thought to play a crucial role in establishing a low transcytosis rate and strengthening cell–cell junctions. 30
Armulik et al. 30 defined a mature PC to be a cell that is embedded in the vascular basal lamina. However, this definition is difficult to apply during angiogenesis. It is assumed that, during angiogenesis, PCs are localized at least partly extramurally and proliferate.
PCs are metabolic sensors that can induce increased blood flow when local substrate levels are low. 123 This PC-dependent blood flow regulation is strongly coordinated by the presence of large numbers of interpericyte TNTs (IP-TNTs; Fig. 7C) to exchange organelles such as mitochondria and to serve as channels for calcium waves that are needed for blood flow regulation by coordinated dilation and contraction of the lumen of capillaries. 138 Increased intraocular pressure in the eye causes damage to IP-TNTs that impairs the blood flow and represents a risk factor for the development of glaucoma.139,140
An important metabolic aspect of interactions between PCs and ECs was revealed recently. 31 ECs are highly glycolytic and the larger part of lactate that is produced is secreted into the ECM making the lactate available for uptake by PCs. Alternatively, lactate may also be transported via the peg–socket gap junction-like EC–PC connections, 122 but this direct intercellular transport has not been demonstrated experimentally, as far as we know. Lactate is essential for PCs and the BBB. EC lactate export is mediated by monocarboxylic transporter (MCT)1 and MCT5 and PC import of lactate is mediated by MCT12. Lactate is used as fuel for OXPHOS, as also has been demonstrated in neurons that import extracellular lactate produced by astrocytes from glucose. Astrocytes also function as storage capacity for glycogen.13,141,142 It is not clear yet whether this exchange of lactate from astrocytes to neurons and from ECs to PCs occurs via conversion into pyruvate in the cytoplasm that is then transported into mitochondria to be used for ATP generation by OXPHOS, or alternatively via a lactate transporter to mitochondrial LDHB in the inner mitochondrial compartment.50,51,139 PCs are characterized by their large mitochondria to provide ATP via OXPHOS which is needed for their contractile function. 121 Diabetes reduces ATP production and increases ROS production, 143 thus affecting the proper functioning of ECs and PCs.144,145 Diabetes also causes loss of PCs by apoptosis,146,147 leading to vascular leakage. 144
PCs also use glycolysis and express glucose transporter (GLUT)1 and GLUT4 that facilitate both insulin-dependent and -independent glucose uptake. 33 This glycolytic activity in PCs is associated with contractile activity. 32
In conclusion, it can be stated that (1) whereas angiogenic ECs preferentially use glycolysis, PCs preferentially use OXPHOS for ATP production; (2) OXPHOS in PCs is dependent on lactate production by ECs; and (3) PCs form a regulatory network by their IP-TNTs to coordinate local dilation and contraction of the lumen of capillaries to autoregulate blood flow and thus provision of metabolites.
Inhibition of ATP Synthesis in Ocular Diseases
As discussed, systemic application of inhibitors of ATP synthesis in cancer patients is not an attractive option because clinical trials of inhibitors of glycolysis are either showing a lack of effects or intolerable side effects, whereas inhibition of OXPHOS is unwanted because of the quality-of-life of cancer patients. In ocular diseases, the situation is fundamentally different because inhibitors of ATP synthesis can be administered locally by injections in the vitreous of the eye or, preferably, using eye drops. Local treatment of ocular pathological angiogenesis may also be an escape route to avoid resistance to anti-angiogenic therapy. 24
Here, we investigate whether this local route may be a therapeutic option to inhibit ATP synthesis in ocular diseases. The first question that arises is which are the healthy components in the eye that cannot survive without ATP synthesis. The obvious cells in the eye that need huge amounts of ATP are the photoreceptors. 148 The largest amounts of ATP in photoreceptors are needed for ion transport (Na+ and Ca+). 149 A single rod cell consumes 1 × 108 ATP/sec in darkness and 2 × 107 ATP/sec in light. 150 Glucose is the major substrate for ATP production in photoreceptors but other metabolites such as lactate, amino acids, lipids, and phosphocreatine are also used.151,152 Most of the glucose is used in the large amounts of mitochondria that are present in photoreceptors. Approximately, two thirds of the ATP synthesis in photoreceptors occur via OXPHOS and one third via glycolysis.151,153,154 Therefore, photoreceptors also need large quantities of oxygen. Oxygen consumption and ATP synthesis are correlated in photoreceptors. 151 This huge need for oxygen can barely be met by oxygen delivery by the choroid and, therefore, very low oxygen levels occur during dark adaptation in the outer retina and consequently HIF1α is constitutionally expressed in the human retina. 148 Based on these considerations, we like to hypothesize that OXPHOS is not an attractive therapeutic target in ocular diseases, but glycolysis may well be as it is actively used for ATP synthesis in all angiogenic EC phenotypes and in PCs (Table 4). Inhibitors of glycolysis should be investigated experimentally in animal models of ocular diseases, for example mice and zebrafish.72,155
Inhibitors of Glycolysis for Therapeutic Application in Ocular Diseases
Several inhibitors of glycolysis have been tested in vivo and/or in clinical trials. These inhibitors are discussed here briefly for their potential local application in eye diseases (Table 5).
Inhibitors of Glycolysis.

Target of glycolysis inhibitors, their specificity, and their potential for local treatment of angiogenic eye diseases. Abbreviations: lnc, long non-coding; MCT1, monocarboxylic transporter 1; PDK, pyruvate dehydrogenase kinase; PFKFB3, phosphofructokinase-2/fructose-2,6-biphosphatase 3; VEGFR, vascular endothelial growth factor receptor; ND, not defined; GLUT1, glucose transporter 1. Specificity of Inhibition: -, unspecific; +/-, moderately specific; +, specific. Potential: -, not a compound to be tested as therapeutic for eye diseases; +, potential as therapeutic for eye diseases.
2-Deoxyglucose as a synthetic glucose analog inhibits the first enzyme of the glycolytic pathway, hexokinase, that phosphorylates glucose into glucose-6-phosphate and 2-deoxyglucose into 2-deoxyglucose-6-phosphate. The latter intermediate inhibits hexokinase non-competitively. Intracellular effects of 2-deoxyglucose have been reviewed recently by Laussel and Léon. 14 The compound has been studied for decades and it became apparent that it does not only inhibit glycolysis but also interferes with many other cellular processes, such as the glycosylation of proteins and lipids and various intracellular signaling pathways. Mammalian cells may even become resistant to 2-deoxyglucose although this has not been proven experimentally. It has been established that yeast can develop resistance against the inhibitor. We conclude therefore that 2-deoxyglucose is not a promising inhibitor of glycolytic ATP production in the eye.
Lonidamine also inhibits hexokinase activity, with hexokinase type II that is bound to the outer surface of mitochondria being far more sensitive to lonidamine than the soluble type I in the cytosol. 156 Phase III clinical trials in lung cancer and breast cancer patients showed only marginal therapeutic effects. Moreover, the half-life of lonidamine in cancer patients was short, ranging between 2 and 12 hr.18,156 Therefore, lonidamine does not seem to be a promising glycolysis inhibitor to be applied in the eye either.
GLUT1 is the major transporter of glucose in ECs (Fig. 8). 157 Its expression is uncoupled from glycolysis in quiescent ECs that express high levels of GLUT1 whereas their glycolysis rate is low due to FOXO1 expression. In this way, transcellular glucose transport from vessel lumen to surrounding tissues is facilitated. 20 During angiogenesis, FOXO1 expression is downregulated by PI(3)AKT phosphorylation that prohibits FOXO1 to enter the EC nucleus.13,20,113 Afterward, GLUT1 expression in ECs is coupled to glycolysis during angiogenesis. Inhibition of GLUT1 reduces EC proliferation during angiogenesis, whereas the numbers of tip ECs are not altered, indicating that GLUT1 expression and elevated glycolysis rates are found in proliferating stalk ECs. 157 Systemic inhibition of GLUT1 has serious side effects such as seizures and neuronal loss, whereas the BBB and likely the BRB are not affected. 157 It seems therefore that inhibition of GLUT1 by local treatment of diseased eyes with a GLUT1 inhibitor may be a therapeutically effective approach for treating ocular pathological angiogenesis. Potential inhibitors of GLUT1 are BAY-876, 157 phloretin, WZB117, and fasentin.156,159 Recently, it was claimed that GLUT3 instead of GLUT1 is a major GLUT in human ECs, whereas GLUT1 would be a major GLUT in mouse ECs. 160 This may be different for GLUT1 in retinal ECs as it is strongly expressed in these ECs, next to other retinal cell types (Fig. 8). Possibly both GLUT1 and GLUT3 are active as GLUTs in human retinal ECs. 160
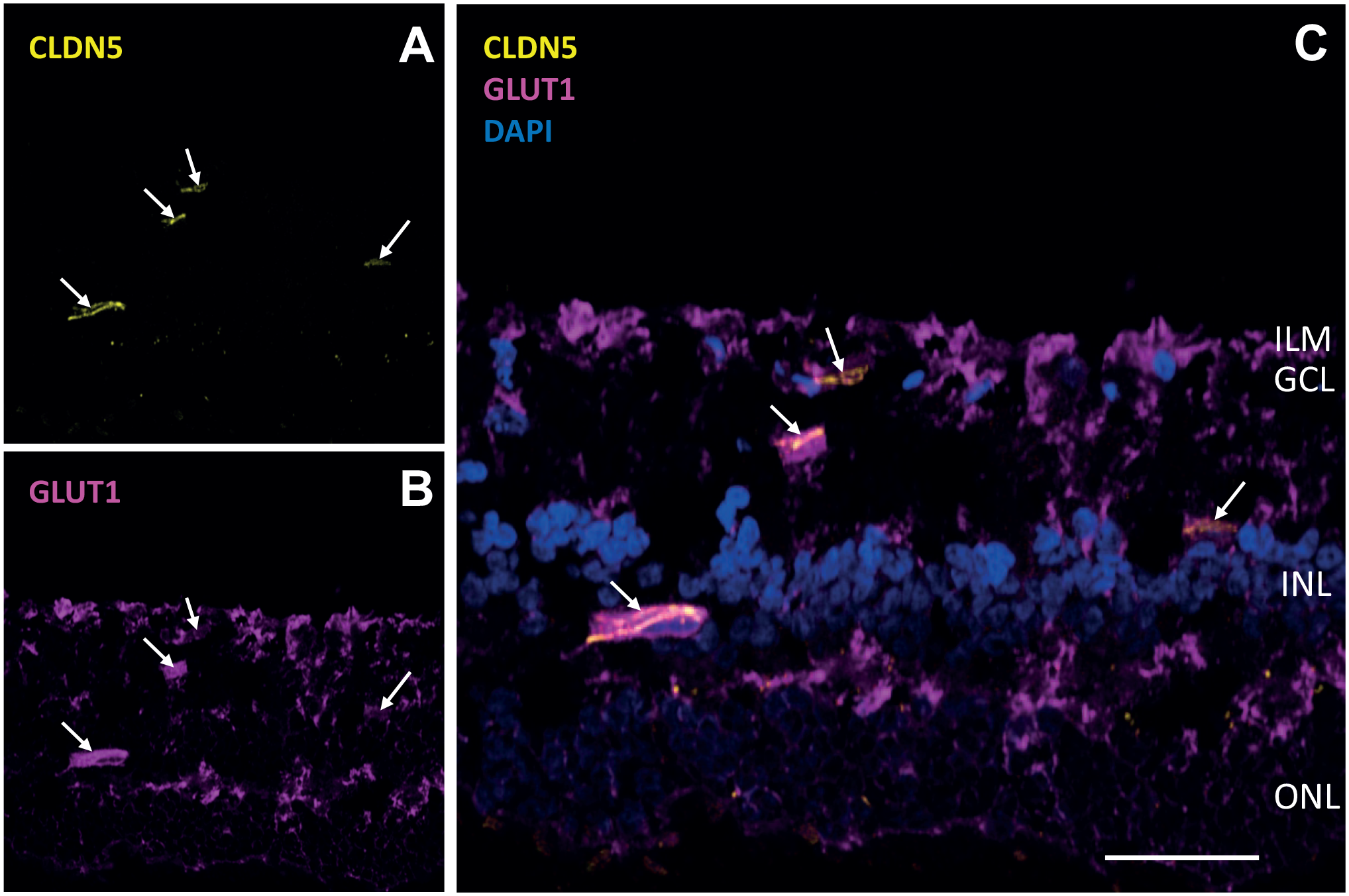
Glucose transporter 1 (GLUT1) and claudin-5 (CLDN5) expression in human retina. Immunofluorescence images of a frozen section of a human retina showing staining of (A) CLDN5 as endothelial cell marker (yellow, arrows), (B) GLUT1 transporter (magenta, arrows), and (C) CLDN5, GLUT1, and 4′,6-diamidino-2-phenylindole (DAPI) as nuclear staining (blue). Bars A–C = 50 µm. Abbreviations: ILM, inner limiting membrane; GCL, ganglion cell layer; INL, inner nuclear layer; ONL, outer nuclear layer. Immunofluorescence staining and imaging procedures are described in Little et al. 158
Potential application of long non-coding RNA (lncRNA)-based inhibition of glycolysis has been suggested to be an alternative anticancer therapeutic approach, despite its reported limitations such as off-target effects, toxicity, and our poor understanding of working mechanisms of lncRNA. 38 Another promising approach for the local treatment of ocular pathological angiogenesis is the application of aptamers. 161 Aptamers are small single-stranded DNA or RNA molecules of up to 100 nucleotides that can bind to a multitude of specific targets that are not necessarily nucleic acids as explained clearly by Iturriaga-Goyon et al. 161 Both lncRNA and aptamers may become new avenues for therapy and in particular for local treatment of ocular pathological angiogenesis.162,163 However, clinical applications of lncRNAs and aptamers are still in their infancy.38,161
MCT inhibitors that interfere with lactate transmembrane transport have been studied for their application as glycolysis inhibitors, including a phase I clinical trial of AZD3965, a potent selective and orally available MCT1 inhibitor. 164 The inhibitor promoted intracellular lactate accumulation that counteracted its use for biosynthetic synthesis resulting in reduced tumor growth. Local administration could become effective as well to inhibit pathological angiogenesis in the eye.
Dichloroacetate is an inhibitor of pyruvate dehydrogenase kinase (PDK). PDK inactivates pyruvate dehydrogenase by phosphorylation and thus prevents pyruvate conversion into acetyl-CoA after it is channeled into mitochondria to fuel OXPHOS. Dichloroacetate is being investigated as an anticancer therapeutic agent because it abrogates glycolysis and stimulates OXPHOS by inhibition of PDK. It has been suggested recently by Schoonjans et al. 165 that dichloroacetate should be tested as an anti-angiogenesis compound and it may well be worthwhile to study its effect locally in ocular pathological angiogenesis.
Phosphofructokinase-2/fructose-2,6-bisphosphatase (PFKFB) isoenzymes activate glycolysis by activating 6-phosphofructokinase (PFK)-1), a rate-limiting step in glycolysis.166,167 Inhibition of PFKFB3 has been studied extensively in pathological angiogenesis8,21,167–170 and cancer.166,169,171 PFKFB3 is the most abundant isoenzyme in ECs.21,167 Several small molecule inhibitors of PFKFB3 have been tested with respect to inhibition of angiogenesis such as AZ67, 172 the cell-permeable selective inhibitor of PFK-2, 3PO, alone or in combination with a VEGFR inhibitor SU4516,10,167 and PFK-158 which is a derivative of 3PO. 8 These PFKFB3 inhibitors have been tested in vitro and systemically in vivo in mice and zebrafish. PFK-158 has been tested in a phase I clinical trial as an anticancer drug and has been suggested to be applied to treat ocular angiogenesis. 101 Therefore, it is a challenging idea to test their application locally in eyes in animal models of pathological angiogenesis.
Recently, Sharma et al.173,174 focused on inhibition of LDHA that converts pyruvate into lactate as therapeutic approach in cancer. There is no reason why inhibition of LDHA and thus of lactate production should not be investigated as anti-angiogenesis agent in the eye. This elegant approach could enhance pyruvate entrance of mitochondria to fuel OXPHOS, whereas synthesis of building blocks for stalk EC proliferation is inhibited. The major obstacle may be the ineffective inhibition of LDHA because of its narrow substrate-binding pocket. 173
In conclusion, rapidly proliferating dCCs and angiogenic ECs mainly use glycolysis for lactate and ATP production. CSCs and angiogenic tip ECs exhibit greater metabolic flexibility, relying relatively more on glycolysis to synthesize pyruvate as compared with their respective counterparts, the dCCs, and stalk ECs. This pyruvate is then channeled into mitochondria to produce ATP by OXPHOS. Inhibition of ATP synthesis via (an)aerobic glycolysis or OXPHOS does not seem attractive as anticancer therapy because efficacy has not been demonstrated in various clinical trials, and because patients need to be treated systemically resulting in unwanted side effects that are affecting the quality-of-life of patients. In ocular pathological angiogenesis, patients can be treated locally in eyes with inhibitors. OXPHOS inhibitors are not an option because of the high ATP demands in, among others, PCs and photoreceptors with OXPHOS as a major source. However, inhibitors of glycolysis for lactate and ATP synthesis seem to be an attractive treatment option as most ECs involved in angiogenesis are glycolytic for lactate and ATP production. This approach should therefore be tested in animal models of pathological ocular angiogenesis. In particular, effects of glycolysis inhibitors on angiogenic ECs and PCs have to be studied because it is not known yet whether inhibition of glycolysis results in unwanted side effects. The most attractive inhibitors to be tested are inhibitors of GLUTs such as GLUT1 and GLUT3, PDK, PFKFB3, and LDHA and inhibitors such as lncRNAs and aptamers that are constructed to inhibit key molecules in glycolysis in angiogenic ECs.
Footnotes
Acknowledgements
This article is my last, first-author article and it is a synthesis of the research in which I participated during the last 10 years on glioblastoma metabolism in the Department of Genetic Toxicology and Cancer Biology at the National Institute of Biology, Ljubljana, Slovenia and during the last 20 years in the research group Ocular Angiogenesis on pathological angiogenesis in the retina in the Amsterdam UMC, Amsterdam, The Netherlands. The article is based on the valuable scientific experiments and discussions in both research groups that ultimately lead to this article. I have always enjoyed participating in the research involved and I still do. I thank all my coauthors sincerely for the fantastic academic activities that we shared in good collegial cooperation.
Competing Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Author Contributions
The authors have contributed to this article as follows: conception and design (CJFVN, VVVH), laboratory experiments, data analysis, and/or assembly of data on glioblastoma cancer cells in primary brain tumors (BM, AH, UB, SKG) and on metabolism in endothelial cells (BYA, NB, MEVBS), electron microscopy (UB, MV) and microscopical fluorescence imaging (PSM, NB, BM, AH, MN, BB, VVVH, MV), scientific discussions (all authors), manuscript writing (CJFVN, BYA, ROS, IK, MV, MEK, MN, BB, VVH), final approval of the manuscript (all authors), and supervision of the entire study (CJFVN, BYA, VVVH).
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: The Slovenian Research Agency (Projects J3-2526, J3-4504, NC-0023, P1-0245, and P3-0108: CJFVN, BM, AH, MN, BB, VVVH, UB, MV), Young Researcher Grant (AH), and the Postdoctoral project Z3-1870 (BB), the European Program of Cross-Border Cooperation for Slovenia-Italy Interreg TRANSGLIOMA (Program 2017; MN, BB), the IVY Interreg Fellowship (VVVH), the EFSD/Boehringer Ingelheim European Research Program in Microvascular Complications of Diabetes 2018 (grant no. BI18_4), and funding through Eye Funds (St. Blinden-Penning, LSBS, St. Oogfonds, RSB) that contributed through UitZicht (UZ2019-23; IK, ROS) and Novartis (Grant 2021-0224; IK, ROS). The funding organizations had no role in the design or conduct of this research. They provided unrestricted grants.