
Abstract
Idiopathic epiretinal membranes are sheets of tissue that develop in the vitreoretinal interface. They are formed by cells and extracellular matrix, and they are considered the expression of a fibrotic disorder of the eye. Confocal and immunoelectron microscopy of the extracellular matrix of excised membranes, revealed high contents of type IV collagen. It was distributed within epiretinal membranes in basement membrane-like structures associated with cells and in interstitial deposits. In both cases, type IV collagen was always associated with type I collagen. Col IV was also coupled with Col VI and laminin. At high magnification, type IV collagen immunolabelling was associated with interstitial deposits and showed a reticular appearance due to the intersection of beaded microfilaments. The microfilaments are about 12 nm in diameter with interbead distance of 30–40 nm. Cells of the epiretinal membranes showed intracellular lysosome-like bodies heavily labeled for type IV collagen suggesting an active role in membrane remodeling. Hence, type IV collagen is not necessarily always associated with basement membranes; the molecular interactions that it may develop when not incorporated in basement membranes are still unknown. It is conceivable, however, that they might have implications in the progression of epiretinal membranes and other fibrotic disorders.
Introduction
Collagens are a broad family of proteins formed by three polypeptide α-chains intertwined together in a triple helix fashion. However, the triple helix domain does not extend for the entire length of collagen molecules as it can range from 96% of the structure to less than 10% depending on collagen type. 1 This diversity forms the basis of the large number of collagen molecules. To date, 28 different collagens have been described. Their classification is not easy for several reasons, including the existence of several α-chains and molecular isoforms. In short, collagens can be grouped into two major categories: collagens that form fibrils (fibrillary collagens) and collagens that do not (non-fibrillary collagens). The latter class of collagens are further divided into several subfamilies including fibril-associated collagens with interrupted triple helices (FACITs), network-forming collagens, membrane anchored collagens with interrupted triple helices (MACITs), microfibrillary interconnecting collagens, and short (developmental) collagens. 2 Type IV collagen (Col IV) belongs to the network-forming collagen subfamily, and it is located within basement membranes (BMs). To form BMs, Col IV is assembled with laminin, entactin and the heparansulfate proteoglycan perlecan. Additional components of BMs are found but are basically expressed in a tissue-specific manner. 3 There are six α-polypeptide chains (α1–α6) that are variously combined together to form three different Col IV isoforms: [α1]2α2, α3α4α5 and [α5]2α6. Whereas [α1]2α2 Col IV is found in all BMs, the other isoforms have a more restricted pattern of expression. Without the presumption to make an exhaustive list, we recall that α3α4α5 isoform has been shown in glomerular BM, 4 retinal inner limiting membrane (ILM), 5 lung alveoli, 6 and seminiferous tubules. 7 In contrast, (α5)2α6 isoform has been found in the parietal layer of Bowman’s capsule, 8 around smooth muscle fibers, 9 and in the epidermis. 10
As a constituent of BMs, Col IV should be found closely associated with cytoplasmic membranes. However, interstitial deposition of Col IV (i.e., not associated with cell membranes as BMs are) has been previously reported in several instances. Interstitial deposition of Col IV has been shown in animal models of kidney fibrosis,11–13 in skin of subjects chronically affected by sclerodermia, 14 and in the interstitial matrix of livers from patients affected by alcoholic cirrhosis. 15 More recently interstitial Col IV deposits have been detected in early fibrotic lesions of idiopathic interstitial pneumonias, 16 and in epiretinal membranes (ERMs).17,18
ERMs are thin sheets of tissue that pathologically develop on the inner surface of the retina, at the vitreoretinal interface. Due to the retraction of ERMs, which is firmly adherent to the underlying ILM, disruption of retinal architecture can occur. They are frequently located at the fovea and affect sight by reducing best corrected visual acuity and by generating distorsion (metamorphopsia). 19 ERMs can be considered the result of fibrotic disorders. Actually, ERMs share with other fibrotic disorders the major hallmarks of tissue fibrosis which are the abnormal and exaggerated deposition of extracellular matrix and the formation of a retracted fibrotic scar tissue which alters the normal architecture and function of the organ. 20
The main collagens involved in fibrosis are type I (Col I) and type III (Col III) interstitial collagens. Interstitial collagens are thought to increase tissue stiffness. This is not only due to the increased amount of collagen fibrils, but also to their crosslinking which in turn induces the development of further tissue fibrosis. 21 Whereas the involvement of Col I and III in tissue fibrosis is a well-established fact, the role played by Col IV is still unclear. In order to shed some light on this matter, the present study analyzed interstitial Col IV deposits in idiopathic ERMs (iERMs) by confocal and immunoelectron microscopy.
Materials and Methods
Antibodies
Rabbit anti-human Col I was from Novus Biologicals (Littleton, CO), goat anti-human Col IV was purchased from SouthernBiotech (Birmingham, AL) and mouse monoclonal anti-human vimentin (Clone V9) was purchased from Sigma-Aldrich (Milano, Italy). Rabbit anti-laminin 1 and 2 and rabbit anti-human type VI collagen (Col VI) were purchased from Abcam (Cambridge, UK), whereas mouse mAb anti-human laminin (γ1 chain-specific, clone 2E8) and mouse mAb anti-human laminin 5 (γ2 chain-specific, clone D4B5) were purchased from Millipore (Temecula, CA). Cy2-conjugated donkey anti-rabbit IgG, tetramethylrhodamine (TRITC)-conjugated donkey anti-goat IgG, Cy5-conjugated donkey anti-mouse IgG and 12 nm gold-conjugated donkey anti-goat IgG were purchased from Jackson Immunoresearch Laboratories (Baltimore, PA), whereas 10 nm gold-conjugated donkey anti-goat IgG was obtained from Abcam and 15 nm gold-conjugated goat anti-rabbit IgG was from British BioCell International (Cardiff, UK).
Samples
Idiopathic ERMs were excised during vitreoretinal surgery by the same surgeon (G.M.T.) between January 2014 and August 2017 at the Ophthalmology Unit of the Department of Medicine, Surgery and Neuroscience of the University of Siena. Patients were treated after being informed of the nature, purpose, implications, and risks of the treatment and after having signed a consent form. A total number of 36 iERMs, 15 of them associated with the ILM, are included in this study and processed as follows. Membranes were fixed with 1.25% glutaraldehyde in 0.1M sodium cacodylate for 24 hr at 4C and postfixed in 1% OsO4 for 2 hr at 4C. After fixation, samples were dehydrated and embedded in Epon following standard procedures. Semithin sections (1 µm thick) and ultrathin sections were, respectively, placed on Superfrost plus slides (Menzel-Glaser, Brounschweig, Germany) and on Neoprene W pre-treated nickel grids (Neoprene W was a kind gift from Safic-Alcan Italia SPA, Garbagnate Milanese, Milan, Italy) or on Formvar-coated copper grids. In addition, three recently sampled iERMs (September/October 2019) were frozen in isopentane prechilled with liquid nitrogen and sectioned with a cryostat as previously reported. 22 Sections of 7-μm thickness were placed on superfrost slides, fixed for 10 min with cold (-20C) acetone or 30 min with buffered formalin and processed for immunofluorescence microcopy.
Immunofluorescence Microscopy
For Epon-embedded iERMs, immunofluorescence was carried out on semithin sections upon Epon removal and antigen retrieval. Epon was removed from sections with a 1:1 solution of ethanol and a saturated solution of NaOH in ethanol for 7 min. Then, slides were washed in ethanol and distilled water and incubated with 20% donkey serum added with 0.3% Triton X-100. Heat-induced antigen retrieval (HIAR) was carried out according to Yamashita and Okada, 23 autoclaving slides in 0.1M Tris (pH 9.0) at 120° for 10 min. Before incubating slides with each primary antibody, sections were always treated with 5% bovine serum albumin (BSA) in phosphate buffered saline (PBS) while before applying each secondary antibody, sections were incubated with 10% non-immune serum belonging to the animal species employed to raise the antibody. The sequence of antibodies employed was the following: anti-vimentin (dilution 1:150) overnight, Cy5-conjugated anti-mouse IgG (dilution 1:100) for 45 min, anti-Col I (dilution 1:100) for 90 min, Cy2-conjugated anti-rabbit IgG (dilution 1:100) for 45 min, anti-Col IV (dilution 1:200) for 2 hr, and TRITC-conjugated anti-goat IgG (dilution 1:200) for 60 min. We also carried out experiments with anti-laminin and anti-Col VI antibodies followed by the proper secondary antibody. Slides were observed with a Zeiss LSM510 laser scanner confocal microscope. Negative controls included omission of the primary antibodies or their replacement with non-immune sera.
For frozen sections, immunofluorescence was carried out with the following sequence of antibodies: goat anti-Col IV (dilution 1:400) for 1 hr, TRITC-conjugated anti-goat IgG (dilution 1:200) for 45 min, rabbit anti-Col VI (dilution 1:100), or rabbit anti-laminin 1 and 2 (dilution 1:100) for 2 hr, Cy2-conjugated anti-rabbit IgG (dilution 1:100) for 45 min.
Immunoelectron Microscopy
In order to retrieve antigenicity, ultrathin sections mounted on Neoprene W-coated nickel grids were placed for 60 min at 95° in 0.5M Tris (pH 9.0) according to Yamashita and Okada. 23 Unspecific binding sites were quenched with 0.2% sodium borohydride for 10 min and, after extensive rinsing, with 1% ovalbumin in PBS. Subsequently, grids were incubated overnight at 4C with anti-Col IV antibody (dilution 1:50). Grids were then washed, quenched with 1% ovalbumin and incubated with 10- or 12-nm gold-conjugated donkey anti-goat IgG antibody (dilution 1:100) for 2 hr. Upon washing, grids were stained with uranyl acetate and lead citrate. For double labeling experiments, grids were additionally incubated with rabbit anti-human Col I antibody (dilution 1:50) for 2 hr at room temperature, washed, quenched with 1% ovalbumin and incubated for 2 hr with 15-nm gold-conjugated goat anti-rabbit IgG antibody (dilution 1:100). Additional experiments were carried out with anti-laminin and anti-Col IV antibodies followed by the proper gold-conjugated secondary antibody. Negative controls included omission of the primary antibodies or their replacement with non-immune sera.
Results
Pattern of Type IV Collagen Deposition in iERMs
Double labeling with anti-Col I and anti-Col IV antibodies detected both antigens in all samples. Col I was always more represented than Col IV. However, the immunoreactivity for Col IV in 20 iERMs (55.5% of samples) (Fig. 1A to C) was particularly relevant. Samples containing the ILM were very useful to define collagen arrangement in iERMs. Proceeding from outside inward (from the side facing the ILM to the side facing the vitreous), ERMs were characterized by a layer of Col I of variable thickness in contact with the ILM followed by a second layer of Col IV and Col I intimately mixed together (Fig. 1D to F). Hence, Col IV was always much more condensed on the side facing the vitreous. ERM cells were almost always in contact with the Col IV-rich layer, gathered on the inner face of the ERMs. Therefore, based on the constant arrangement of Col I and Col IV, the inner and the outer sides of iERMs could be easily identified even in samples without the ILM: the layer on the outer side (retinal side) was formed only by Col I, whereas in the inner side (vitreous side) Col IV was preferentially gathered where the cells are located (Fig. 1G to J). The deposition of Col IV followed four different patterns but the same ERM frequently displayed areas with diverse patterns. In 23 samples (64% of cases), Col IV was found deposited interstitially (i.e., large amounts of collagen were not in direct contact with cells) (Fig. 1A to C); in 4 samples (11.1% of cases) it was deposited in a dot-like fashion around cells (Fig. 2A and B); in 27 iERMs (75% of cases) it showed a continuous BM-like arrangement (Fig. 2C and D), and in 18 membranes (50% of cases) it formed tracts of a discontinuous BM-like structure (Fig. 2E and F). The continuous appearance of the BM-like Col-IV deposition was maintained even when observed at higher magnification (data not shown). Considering single membranes, we observed that nine samples (25% of cases) displayed an interstitial deposition, as well as continuous and discontinuous BM-like arrangements; five samples (13.9% of cases) showed exclusively an interstitial deposition with no identifiable BM or BM-like structure; five samples (13.9% of cases) had interstitial deposition associated with a continuous BM-like arrangement; five samples (13.9% of cases) were provided with only a continuous BM-like structure; four samples (11.1% of cases) showed alternated tracts of continuous and discontinuous BM-like arrangements; two samples (5.5% of cases) displayed interstitial deposition associated with a discontinuous BM-like structure; two samples (5.5% of cases) had dot-like deposition, as well as continuous and discontinuous BM-like arrangements; one sample (2.8% of cases) displayed interstitial accumulation associated with a continuous BM-like structure and areas with dot-like deposition; and one sample (2.8% of cases) showed all the deposition patterns that could be observed (i.e., interstitial, dot-like, continuous BM-like and discontinuous BM-like).
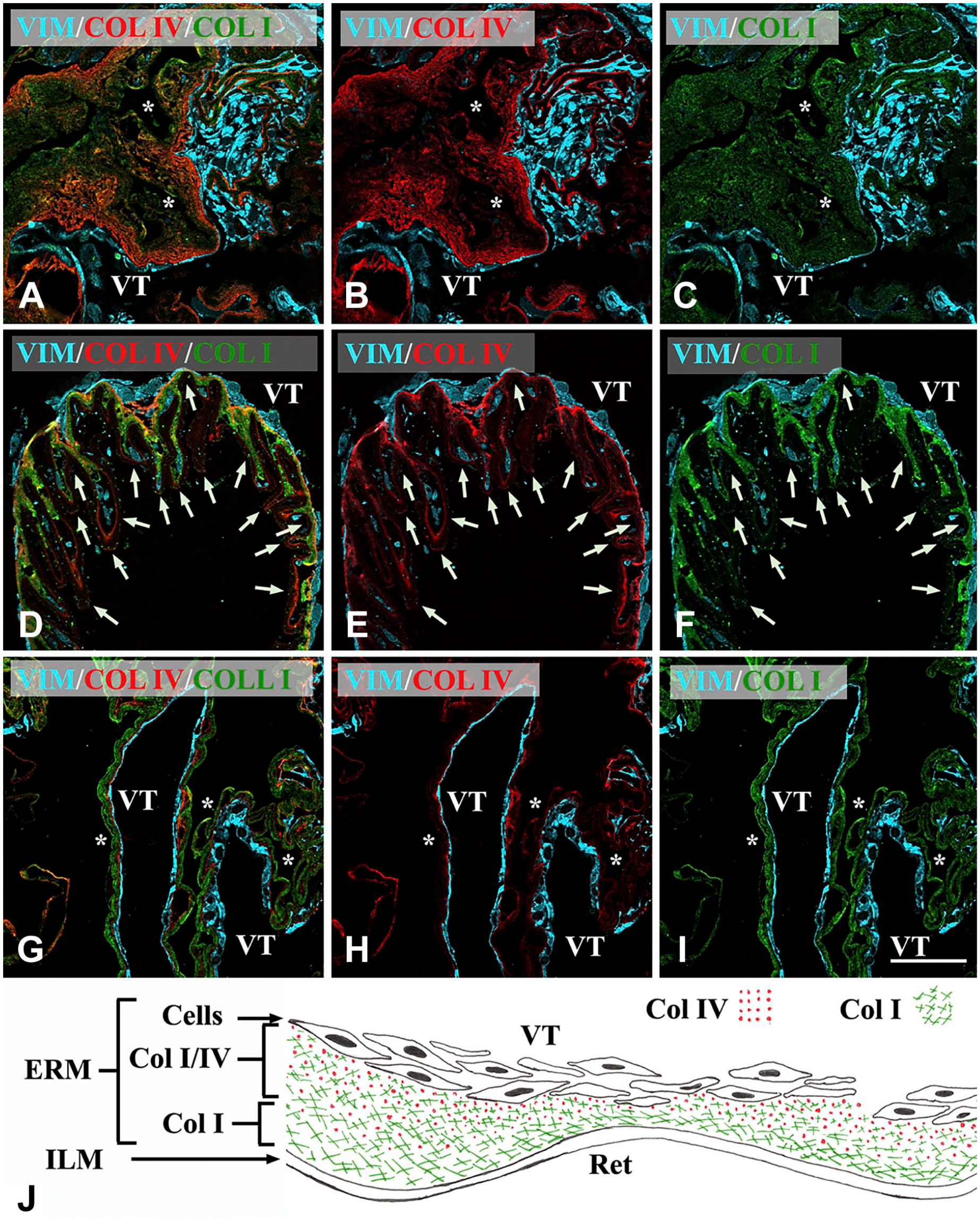
Confocal microscopy of iERMs. Membranes were triple labeled with anti-vimentin, anti-Col IV, and anti-Col I antibodies. Anti-vimentin antibody was employed to stain cells. Figures of the panel are representative of the whole set of samples. (A–C) A thick layer of extracellular matrix is mainly Col I-immunoreactive. However, even Col IV immunoreactivity appears spread through a large portion of the membrane, with the same lamellar arrangement showed by Col I. Cells are all gathered on the same side of the membrane. The sample does not contain the ILM. The membrane is highly convoluted and it is difficult to follow its course. (D–F) iERM associated with the ILM. Whereas ILM is poorly stained (arrows), ERM is strongly fluorescent for both Col I and Col IV. However, a layer almost exclusively Col I-immunoreactive is visible intervening between the ILM and a second layer where both collagens co-localize (yellow-orange color). (G–I) ERM showing the usual three-layered arrangement: one layer exclusively Col I-immunofluorescent (presumably the layer that in situ was adjacent to the ILM), one layer co-stained with anti-Col I and anti-Col IV antibodies, and one layer of cells. Magnification bar = 50 µm; (J) Schematic reconstruction of collagen I and IV arrangements in iERMs. ERMs are attached to the ILM with a layer of Col I. Proceeding toward the VT Col IV appears and increases in concentration. Cells are usually gathered on the vitreous side of the membrane. *Side of the membrane that was detached from the ILM. Abbreviations: iERM, idiopathic epiretinal membranes; ILM, inner limiting membrane; VT, vitreous side of the membrane; Ret, Retinal side; Col I, type-I collagen; Col IV, type-IV collagen.
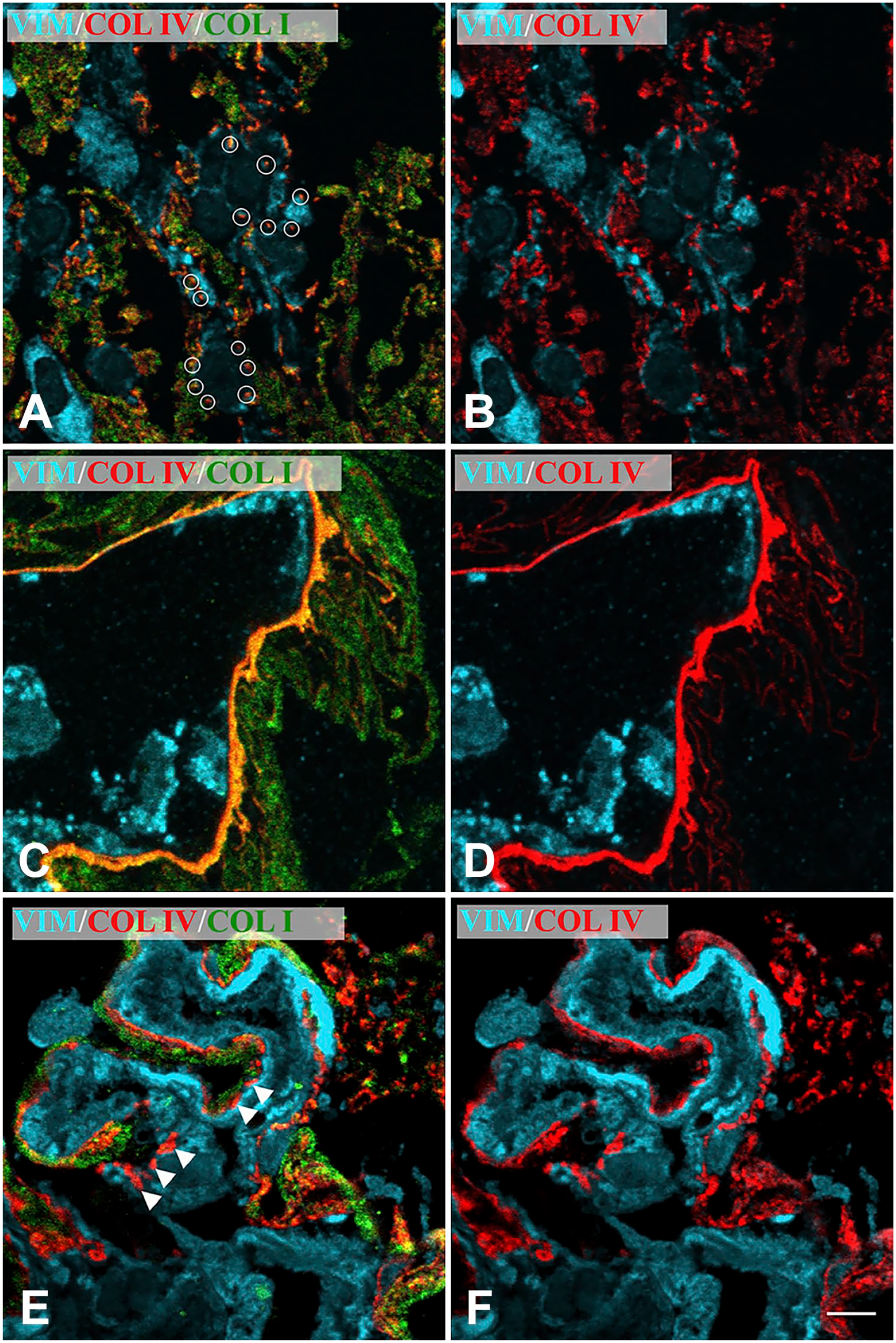
Confocal microscopy of iERMs without associated ILM. BM-like arrangements of Col IV. Membranes were triple labeled with anti-vimentin, anti-Col IV and anti-Col I antibodies. Figures of the panel are representative of the whole set of samples. (A–B) Col IV is deposited around cells in a dot-like fashion (highlighted by white circles) and co-localizes with Col I. (C–D) Col IV is deposited as if forming a true BM. However, even in this case, Col IV co-localizes with Col I. (E–F) Col IV is deposited forming a sort of discontinuous BM (arrowheads). In this case, some deposits of Col IV co-localize with Col I, others seem to be Col I-free. Magnification bar = 5 µm. Abbreviations: iERM, idiopathic epiretinal membranes; ILM, inner limiting membrane; BM, basement membrane.
When considering interstitial deposits, Col IV could show a lamellar arrangement or a more compact organization (Fig. 3). Though in diverse areas, both structural arrangements could be present within the same ERM. For this reason, it was difficult to make a valid estimation over their incidence. Certainly the compact arrangement was by far much rarer.
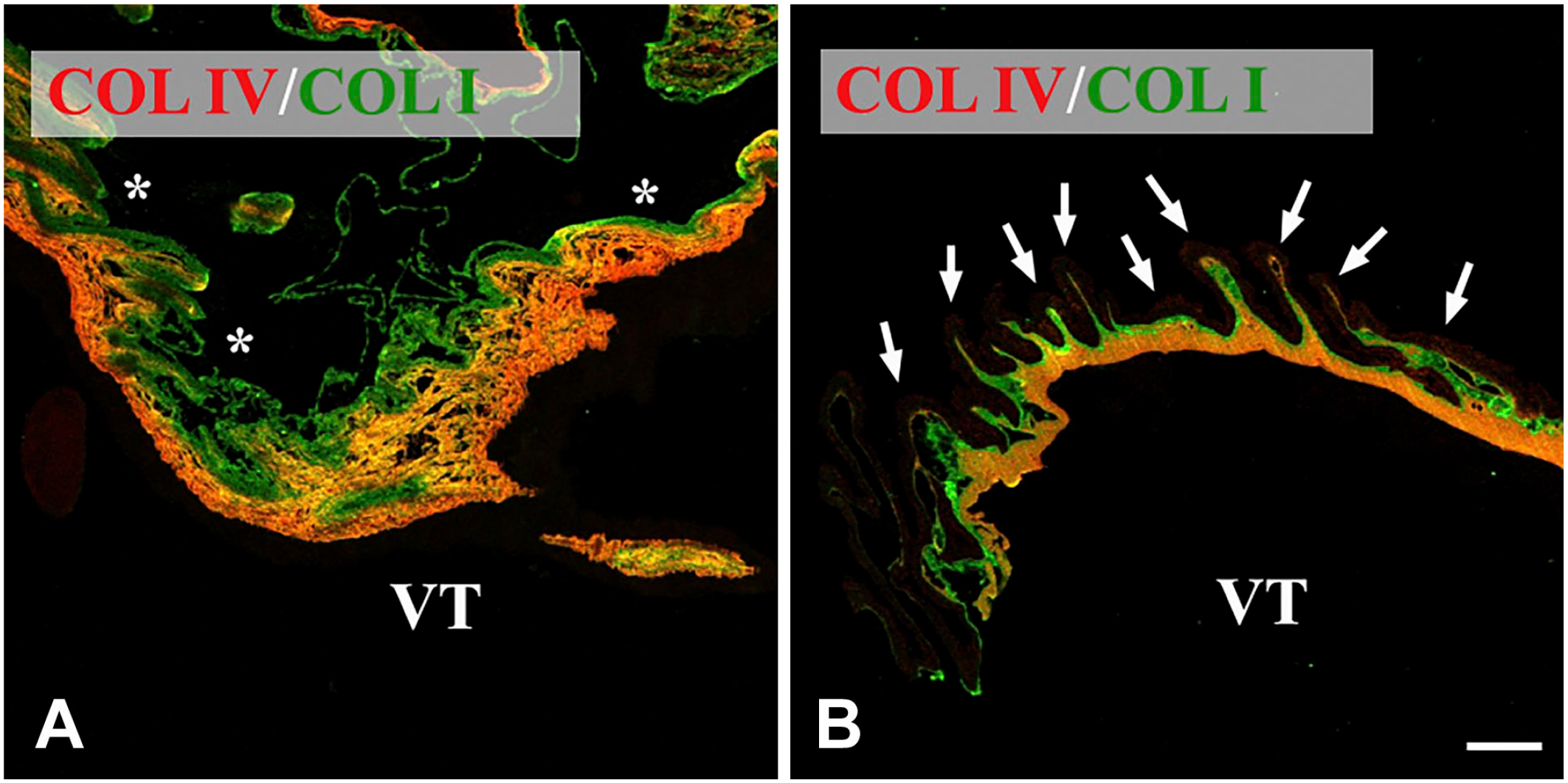
Confocal microscopy of iERMs. Membranes were double labeled with anti-Col I and anti-Col IV antibodies. Col IV deposition closely follows Col I arrangement. Figures of the panel are representative of the whole set of samples. (A) Lamellar arrangement of Col I and Col IV. Note the usual progression of Col IV deposition: on one edge of the membrane Col IV does not co-localize with Col I. Then, proceeding through the membrane, Col IV appears. Though always intermingled with Col I (yellow color), its immunoreactivity progressively increases toward the opposite edge of the membrane (the color turns increasingly orange). The sample does not contain ILM. (B) Compact arrangement of Col I and Col IV. The iERM is associated with the ILM (arrows) which is poorly stained. Once again, the usual distribution of collagens is visible: a layer of Col I is adjacent to the ILM and then a thick layer of intimately mixed Col I and Col IV appears on the opposite edge of the membrane (orange color). *Side of the membrane that was detached from the ILM. Magnification bar = 20 µm. Abbreviations: iERM, idiopathic epiretinal membranes; ILM, inner limiting membrane; VT, vitreous side of the membrane.
We also assessed co-localization of Col IV with Col VI or with laminin. Unfortunately, laminin and Col VI immunoreactivities were negligible on Epon-embedded samples (data not shown). However, a good reaction was achieved on frozen sections where a generalized co-localization of Col IV with Col VI could be observed. Interestingly large tracts of the membrane showed a fine reticular appearance due to intertwined collagen filaments co-stained with both anti-Col IV and anti-Col VI antibodies (Fig. 4A to C). Col IV also co-localized with laminin. Yet, in this case, laminin and Col IV immunofluorescence did not always closely match as in some focal areas laminin or Col IV were apparently alone (Fig. 4D to F).
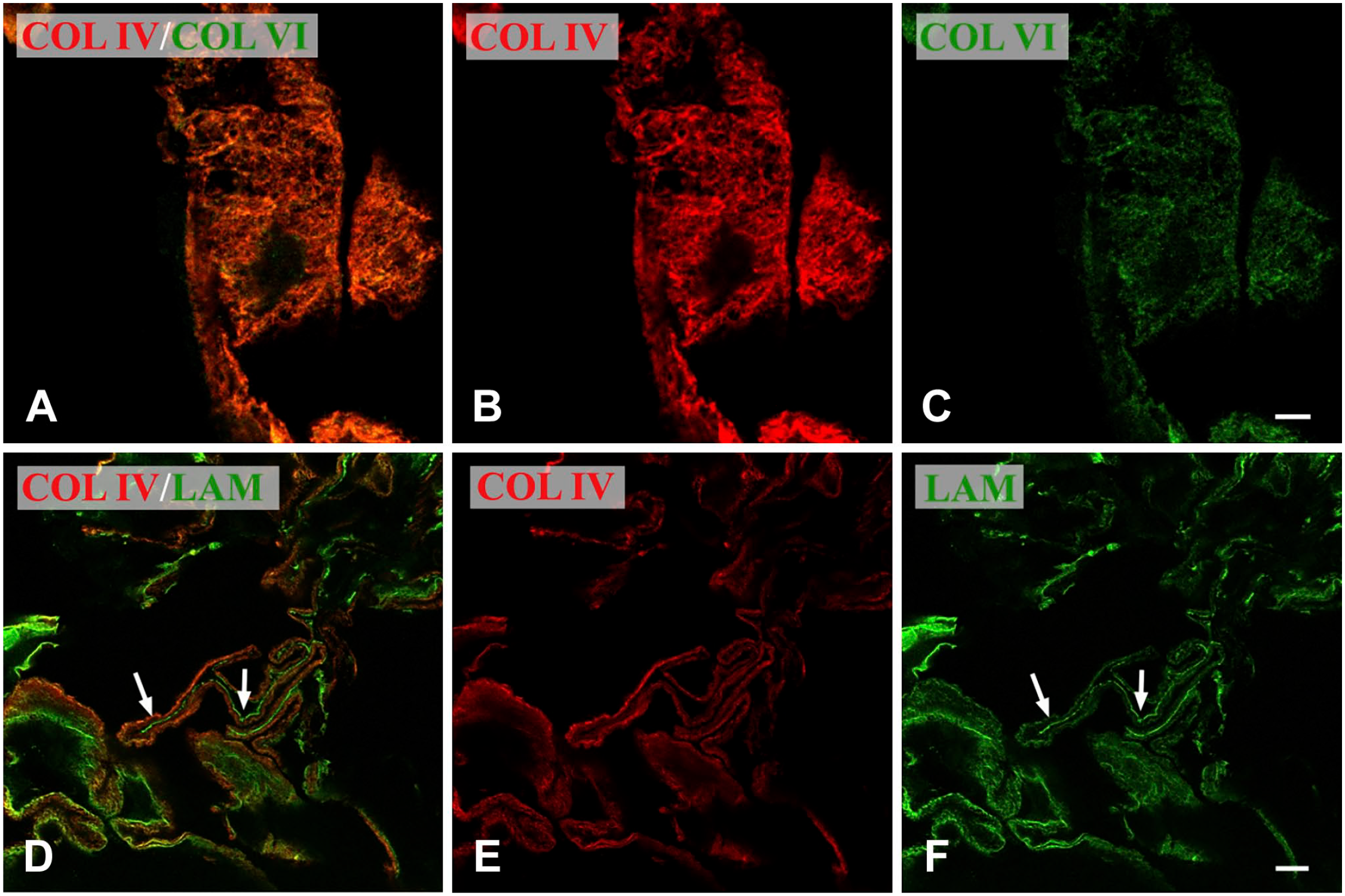
Confocal microscopy of iERM. Figures of the panel are representative of the whole set of experiments carried out on frozen sections. Results were the same on sections fixed with aceton or with formalin. (A–C) The membrane, double labeled with anti-Col IV and anti-Col VI antibodies, shows a fine reticular arrangement of double-labeled filaments (orange color). Formalin-fixed section. Magnification bar = 5 µm; (D–F) The membrane was double labeled with anti-Col IV and anti-laminin 1 and 2 antibodies. A general co-localization of the antigens is evident (orange color). However, some areas of the membrane show a very strong laminin immunoreactivity (arrows) that does not co-localize with Col IV. Formalin-fixed section. Magnification bar = 10 µm. Abbreviations: iERM, idiopathic epiretinal membranes.
Ultrastructural Localization of Type IV Collagen
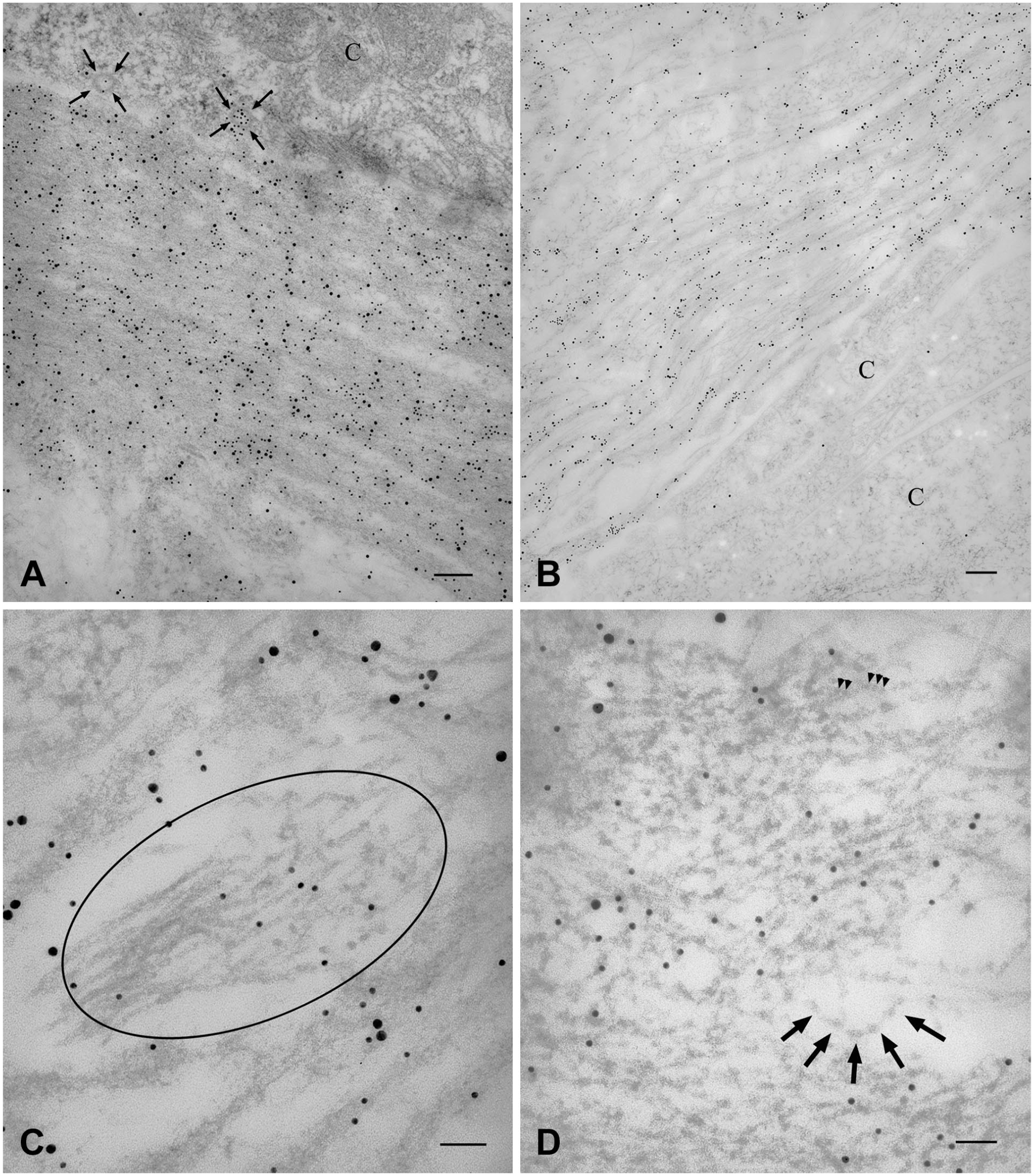
As expected, single labeling of ultrathin sections with anti-Col IV antibody showed a labeling located close to cytoplasmic membranes, in BM-like structures most frequently with a continuous appearance (Fig. 5A). This was in agreement with results obtained with the confocal microscope. However, confocal microscopy also showed that Col IV was almost always co-localized with Col I which, by definition, is not a component of BMs. Indeed, when cut tangentially, Col IV-labeled BM-like structures unveiled the presence of several collagen fibrils extending into the surrounding connective tissue (Fig. 5B). In addition, still consistent with results achieved by confocal microscopy, Col IV labeling was also seen on collagen condensations located far from cells (Fig. 5A). Even such interstitial deposits of Col IV were infiltrated by collagen fibrils (Fig. 5C). In order to confirm that fibrils associated with Col IV were formed by Col I, as suggested by confocal microscopy, we double labeled ultrathin sections with anti-Col I and anti-Col IV antibodies unveiled by gold particles of different size. Type IV collagen-labeled interstitial deposits were indeed also labeled with the anti-Col I antibody (Fig. 5D). As internal controls, areas containing long-spaced collagens, which have been shown to be formed by Col VI,18,24 were not reactive neither with anti-Col IV nor with anti-Col I antibodies and areas containing fibrillary collagens were labeled exclusively by anti-Col I antibody. On the other hand, omission of the primary antibodies or their replacement with non-immune sera resulted in absence of gold-particle deposition (Supplementary Figure 1).
Transmission electron microscopy of iERMs. Figures of the panel are representative of the whole set of experiments. (A-C) Sections were labeled with anti-Col IV antibodies (12-nm gold particles). (D) Sections were double-labeled with anti-Col IV (12 nm-gold particles) and anti-Col I (15 nm-gold particles) antibodies. (A) Cells (C) are lined by a Col IV-labeled BM-like structure (arrows). Col IV labeling is not restricted to BM-like structures as it appears located also over interstitial collagen condensations (arrowheads). Magnification bar = 200 nm; (B) Grazing section of a cell-lining BM-like structure. Col IV labeled the BM-like structure which also appears infiltrated by collagen fibrils. Magnification bar = 200 nm; (C) Col IV-labeled interstitial deposit. Twelve-nm gold particles unveiling Col IV immunoreactivity decorated collagen deposits. Such dense collagen accumulations are also heavily infiltrated by collagen fibrils. Magnification bar = 100 nm; (D) A large collagen condensation on the bottom of the figure is co-labeled with 10 and 15 nm-gold particles (respectively unveiling Col IV and Col I). In the center of the figure, a large deposit of long-spacing collagen (LS) displays its typical zebra pattern. Only few 15-nm gold particles can be found on LS deposit as the result of a modest background. On the top of the figure, collagen fibrils are decorated exclusively by 15 nm-gold particles. Magnification bar = 300 nm. Abbreviations: iERM, idiopathic epiretinal membranes; BM, basement membrane.
We also investigated the relationships between Col IV and Col VI or laminin. Unfortunately, under our experimental conditions, we could not achieve any consistent immunogold labeling for either Col VI or laminin (data not shown).
An important result was to find that Col IV immunoreactivity was not restricted to the extracellular matrix. By confocal microscopy, 27.8% of samples showed intracellular staining for Col IV (Fig. 6A). Staining was not diffuse throughout the entire cell body but apparently it was confined to large organelles. Immunoelectron microscopy revealed real intracellular labeling in five cases (13.9% of samples). The intracellular labeling for Col IV was located within large lysosome-like vacuoles (Fig. 6B). Co-labeling, although minor, with anti-Col I antibody was also observed within the same vacuoles. In addition, in some cells Col IV labeling was present within 110 nm large vesicles (outer diameter) that lied close to the cell membrane (Fig. 7A).
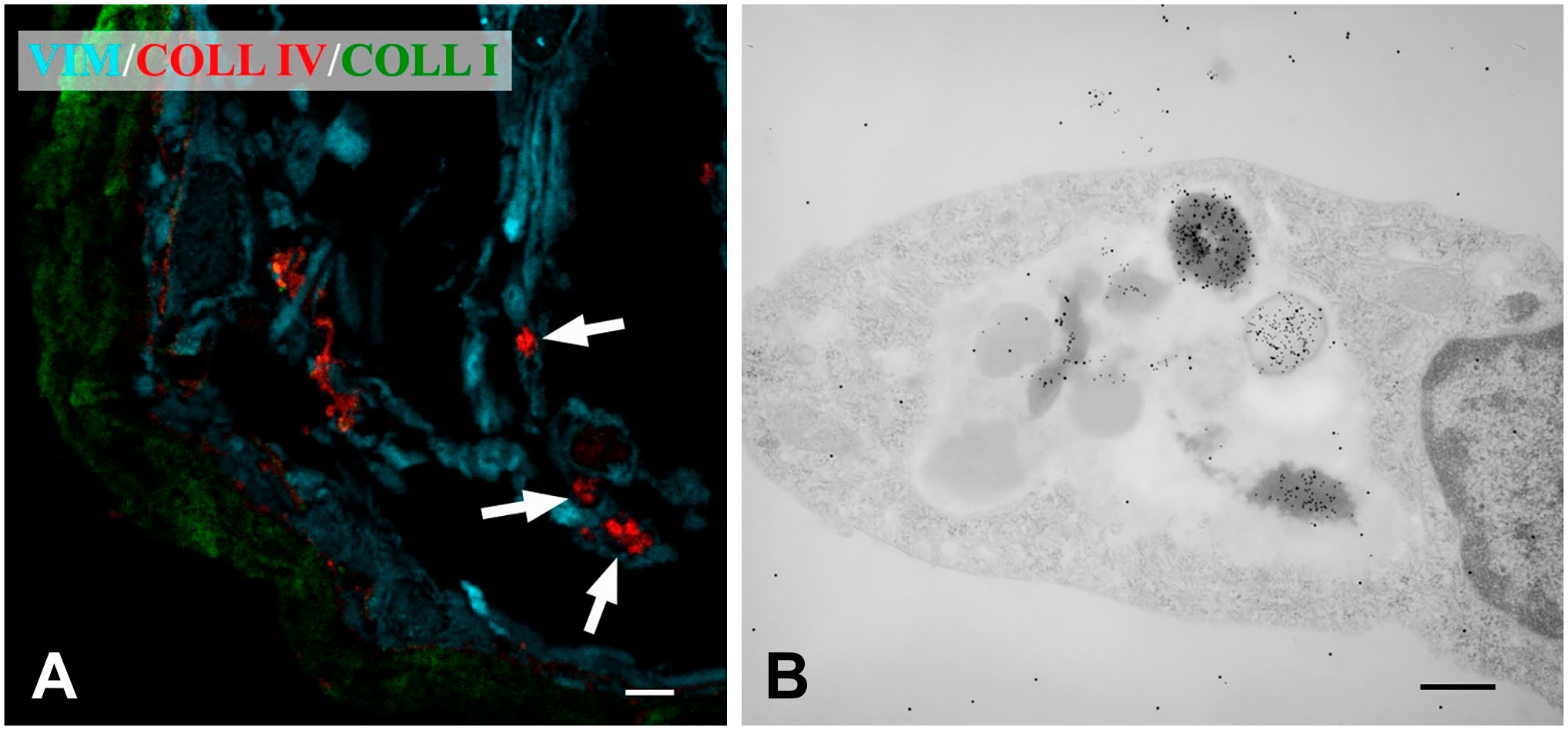
Col IV-containing cells in iERMs. Figures of the panel are representative of the whole set of experiments. (A) Confocal microscopy. Membrane was triple labeled with anti-vimentin, anti-Col IV and anti-Col I antibodies. Large accumulations of Col IV are apparently located within cells. Magnification bar = 5 μm; (B) Transmission electron microscopy of the same membrane confirmed findings from confocal microscopy. Section, double-labeled with anti-Col IV (12 nm-gold particles) and anti-Col I (15-nm gold particles) antibodies, shows cells with large vacuoles containing lysosome-like bodies strongly labeled with 12-nm gold particles and by far less 15-nm gold particles. Magnification bar = 200 nm. Abbreviations: iERM, idiopathic epiretinal membranes.
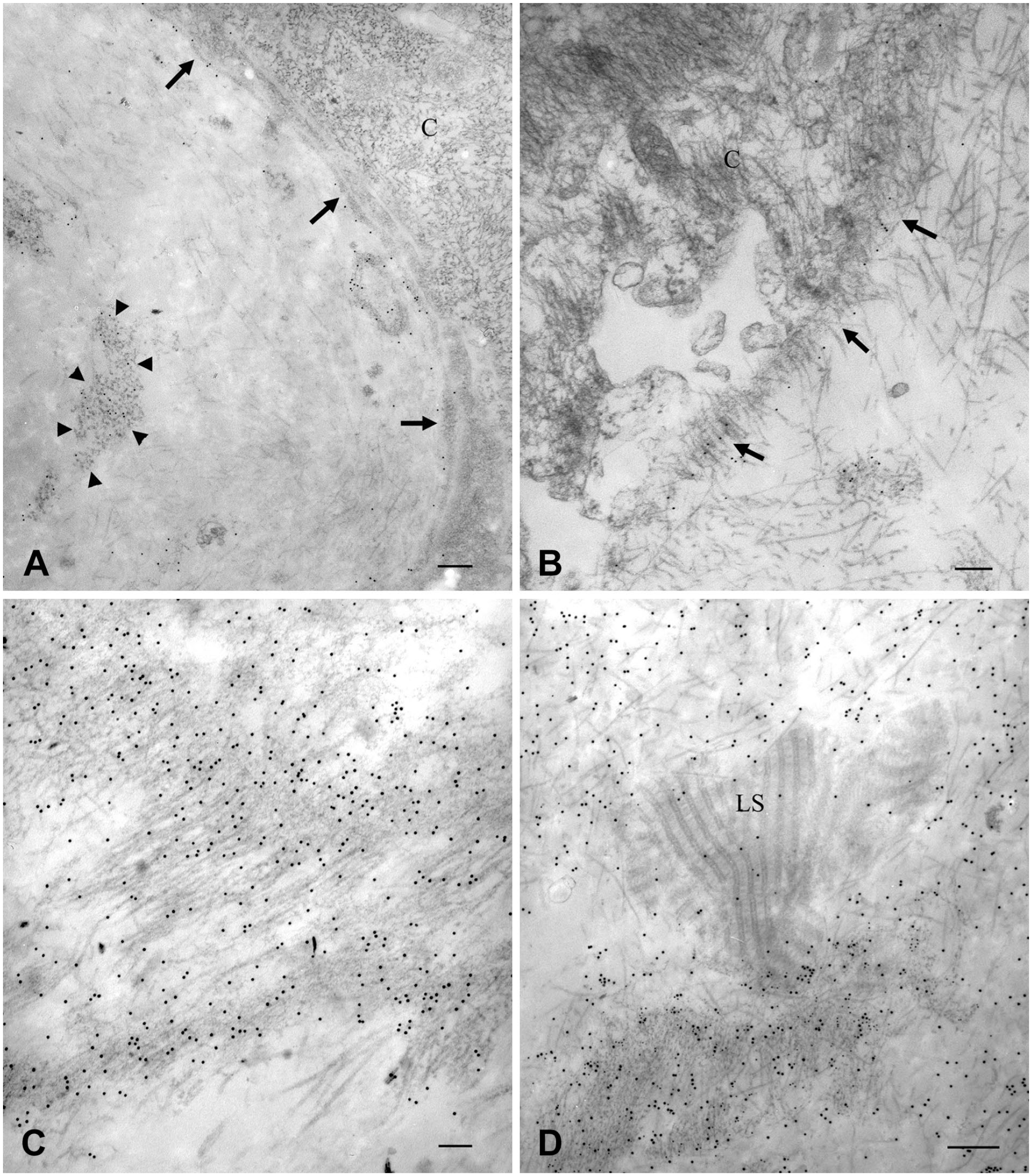
Transmission electron microscopy of the same samples shown in Figure 3. Section have been double-labeled with anti-Col IV (12-nm gold particles) and anti-Col I (15-nm gold particles) antibodies. (A) Compact arrangement of Col IV. The section shows a thick and dense layer of collagen adjacent to cells (C). Anti-Col IV immunolabelled collagen layer is not as compact as it appeared by confocal microscopy and the tendency to form lamellae is evident even in this case. However, collagen lamellae are rather compacted together. This part of the membrane is co-labeled by both antibodies. Cells show 110-nm large vesicles (arrows) close to the plasma membrane labeled with anti-Col IV antibody. Magnification bar = 100 nm; (B–D) Lamellar arrangement of Col IV. B) The figure shows the portion of the ERM which is adjacent to cells (C). Collagen is organized in lamellae with an arrangement looser than the sample shown in (A). Lamellae are double labeled with both antibodies. Magnification bar = 200 nm; (C) The encircled area, labeled exclusively with 12-nm gold particles, shows a tuft of microfibrils. Within the encircled area, microfibrils on the right form a loose network and have a beaded appearance; those on the left of the encircled area are smoother and arranged almost in parallel. Magnification bar = 50 nm; (D) An area heavily labeled with anti-Col IV antibody shows a reticular aspect. The network is formed by beaded microfibrils that join one another in correspondence of the beads. In some areas, microfibrils associate with one another apparently in parallel and in a staggered fashion as revealed by beads arrangement (arrowheads). At the periphery of the network, some beaded microfibrils, not yet entangled in the network, show a linear arrangement (arrows) and the interbead distance of 30-40 nm can be easily measured. Magnification bar = 50 nm. Abbreviations: ERM, epiretinal membranes.
Ultrastructural Features of Type IV Collagen-labeled Microfilaments
Immunogold double labeling of samples showing Col IV compact or lamellar arrangements confirmed the intimate relationship with Col I previously observed by confocal microscopy (Fig. 7A and B). However, even those ERMs that by confocal microscopy showed a more compact texture (Fig. 3B), unveiled a lamellar arrangement by electron microscopy. Yet, lamellae were very close together and partly fused (Fig. 7A). ERMs with a plain Col IV lamellar deposition visible by confocal microscopy (Fig. 3A), showed a looser arrangement by electron microscopy (Fig. 7B). High magnification of the same samples showed that anti-Col IV-labeled microfibrils had two diverse organizations. Labeling was located on microfibrils with a diameter of about 12 nm (Fig. 7C) or on a network of irregularly quadrangular 40- to 80-nm-wide meshes (Fig. 7D). In the last case, the network appeared formed by strands joined one another in correspondence of slightly larger nodes. At their periphery, networks resolved in single strands which appeared as fine beaded microfilaments with an interbead distance of 30–40 nm (Fig. 7D). Beads had a diameter of about 15 nm, and they appeared as the sites (nodes) were microfilaments joined together to generate the network. In some cases, beads could also be seen closely associated in a staggered fashion (Fig. 7D). Areas with smooth microfilaments could also be seen. However, smooth microfilaments were few and continuous with the beaded ones (Fig. 7C). Col I had the usual fibrillary appearance. Though not as thin as Col IV microfibrils, Col I fibrils were very fine as they measured about 15 nm in diameter.
Discussion
Excised ERMs are typically very small samples which make their investigation troublesome. They are difficult to handle, and tissue samples can be so poor that studying them by both light and electron microscopy is quite difficult.17,25 For this reason, we devised an investigation using samples processed for electron microscopy that we also employed for light microscopy. This approach has its advantages but also drawbacks. For instance, a large number of immunolabelling were carried out on the same sample. Yet, in spite of HIAR protocols, some antibodies (anti-laminin and anti-Col VI antibodies in our case) did not yield any labeling either because of the fixation or because of the harsh procedures required for resin removal. For these antigens, we had to carry out immunofluorescence microscopy on frozen samples which, on the other hand, could not be checked with the electron microscope.
In general, ERMs are formed by cells and extracellular matrix. Whereas studies on ERM cells are numerous, investigations on the extracellular matrix are very few.17,18,25,26 ERMs have been shown to contain variable amounts of Col I, type II collagen (Col II), Col III, Col IV, and Col VI. Whereas Col VI deposits were studied in more detail, 18 little attention was given to Col IV which, nevertheless, appears to be highly represented.17,18 Our results confirm that iERMs are highly immunoreactive with anti-Col IV antibodies. In addition, we also show that the pattern of Col IV deposition is quite peculiar as it is not restricted to BMs. Indeed, even the thin Col IV-positive sheet-like structures lining ERM cells do not seem to be conventional BMs as they almost invariably contain Col I. For this reason, we are cautious to consider them as true BMs and we prefer to define them as BM-like structures. Actually, though occasionally, association of Col I with BMs has been previously reported in the nasal mucosa of allergic subjects, 27 in retinal arterioles and venules, 28 in the trabecular beams of human trabecular meshwork, 29 in aged human ciliary body, 30 and in human iris. 31 Curiously, with the exception of the allergic nasal mucosa, all other reports deal with the human eye.
Our results show that Col IV-positive BM-like structures in ERMs can be arranged according to three different patterns of deposition as they are continuous, discontinuous, or organized in a dot-like fashion around cells. The significance of such diverse arrangements is at present unclear.
Idiopathic ERMs are also characterized by Col IV deposited in large interstitial accumulations. Confocal and immunoelectron microscopy are consistent to show intimate associations of Col I and Col IV. Such a relationship is difficult to explain since direct interaction between the two molecules has never been demonstrated. However, as some other extracellular matrix components can bind both collagens, their interaction could be just an indirect one. For instance, vitronectin and Col VI, with the latter also being abundant in ERMs, 18 can bind both Col IV and Col I.32–35 Even nidogen, another mandatory component of BMs, binds both types of collagens, though with diverse affinities. 36 Indeed, as our experiments on frozen sections confirmed co-localization of Col IV and Col VI, Col VI could be a candidate molecule for mediating Col I-Col VI interactions.
Since Col IV co-localizes with laminin in BMs, we tried to demonstrate the same co-distribution in ERMs. Unfortunately, though we tried several antibodies, results were always inconclusive with Epon-embedded material. On frozen sections, however, a strong immunoreactivity could be seen with antibodies raised against laminins 1 and 2. Though laminin immunoreactivity in ERMs was previously reported by others, 18 our experiments show that it mainly co-localizes with Col IV, confirming that deposition of Col IV is coupled with laminin as it regularly occurs in BMs. Yet, focal areas where laminin and Col IV apparently are not associated can also be found. Laminin alone, on the other hand, is a finding to be expected as laminin secretion always precedes Col IV deposition even in BM formation.
Col IV staining without any apparent association with cells was confirmed at the ultrastructural level, where Col IV immunogold is not just restricted to BM-like structures but also involves interstitial collagen condensations. Anti-Col IV-labeled interstitial deposits appear formed mainly by beaded microfibrils with a diameter of 12 nm. At regular intervals along the length of the microfilaments, globular-like expansions (beads) increase the diameter up to 15 nm. This arrangement reminds us of the end-to-end association of Col VI molecules which also gives rise to beaded microfilaments. 37 Failure to label our samples with anti-Col VI antibody by transmission electron microscopy prevents us to rule out co-localization of Col VI and Col IV at the ultrastructural level. However, as the beaded repeat of 105 nm characteristic of Col VI does not match the interbead distance (length 30–40 nm) that is observed in Col IV-labeled microfilaments, we believe safe to exclude that beaded microfilaments are made of Col VI. Formation of linear beaded microfilaments seems to be just a step for the generation of a dense network which is the most frequent finding. This network is characterized by 40–80 nm wide irregular meshes bordered by strands of microfilaments that join together in globular expansions. If this network is the result of just Col IV polymerization or of the interaction with some other protein is unknown. However, lateral association of Col IV molecules can certainly account for the generation of a network with such small meshes as previously reported. 38 What is more difficult to explain is the formation of linear beaded microfilaments. Microfilaments with such short interbead distance should require, at the very least, an unexpected level of lateral association involving the entire length of Col IV molecules assembled in a staggered fashion. However, such a pattern of polymerization for Col IV has never been reported.
In addition to the staining of the extracellular matrix, anti-Col IV labels some ERM cells. Immunoelectron microscopy confirms such labeling and allows to pin-point the precise location of the labeling within large compartments containing lysosome-like bodies. This finding suggests that ERM cells are engaged in the active remodeling of ERMs, reabsorbing both Col IV and Col I. Collagen molecules can be degraded extracellularly by metalloproteases. 39 Fragments generated from the enzymatic activity of metalloproteases can be further digested extracellularly by gelatinases, 40 or can be taken up by cells for degradation within lysosomes. 41 However, collagen fibril direct uptake by phagocytosis is also an option.42,43 In our case, we cannot say with certainty which pathway has been activated for Col IV reabsorption. However, as endocytosed intact collagen fibrils have never been observed in this study and some cells show anti-Col IV-labeled vesicle with a diameter (110 nm) compatible with their identification as clathrin-coated vesicles, the more likely chance is that the uptake involves degraded small collagen fragments that are subsequently moved to the lysosomal compartment. Yet, intracellular Col IV-immunoreactive vesicles could be also secretory vesicles caught in the act of moving toward the plasma membrane. Indeed, data in our hands are not enough to determine with certainty if Col IV-labeled vesicles are steps of endocytosis rather than exocytosis processes.
Intracellular staining with anti-Col IV antibodies was observed in about one membrane out of four. When consecutive sections were checked with the electron microscope, in half cases the labeling appeared only as the result of invagination of the extracellular matrix within the cells. However, the remaining cases were true intracellular labeling. The reason for the lack of intracellular staining in the great majority of the membranes is at present difficult to explain since ERMs, being tissue of new production over the retina, cannot be compared with control samples or intact portion of tissue around nor is there a disease stage for the ERMs which cause a visual reduction and which then require surgical intervention. In summary, we have shown that iERMs are provided with large amounts of Col IV which assemble in BM-like structures and, more interestingly, accumulate in interstitial deposits. Though the relevance of such findings in the development and progression of ERMs is still to be determined, the molecular interactions that Col IV may acquire outside BMs is a matter that deserves attention for their possible implications in other fibrotic contexts.
Supplemental Material
2019-00078R1_Production_Supplemental_Figure_1_online_supp – Supplemental material for The Peculiar Pattern of Type IV Collagen Deposition in Epiretinal Membranes
Supplemental material, 2019-00078R1_Production_Supplemental_Figure_1_online_supp for The Peculiar Pattern of Type IV Collagen Deposition in Epiretinal Membranes by Marì Regoli, Gian Marco Tosi, Giovanni Neri, Annalisa Altera, Daniela Orazioli and Eugenio Bertelli in Journal of Histochemistry & Cytochemistry
Footnotes
Acknowledgements
We are indebted to Dr. Moïse Bendayan for critical reviewing of the manuscript.
Competing Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Author Contributions
E.B. designed the study and wrote the manuscript; G.M.T. and G.N. provided the excised epiretinal membranes for the study; G.M.T. also contributed to the design of the study. M.R. contributed to the design of the study, performed immunoelectron microscopy, and reviewed the manuscript; D.O. and A.A. carried out immunoreactions for confocal microscopy and reviewed the manuscript. All authors have read and approved the final version of the manuscript.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by intramural funds of the University of Siena.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.