
Abstract
One of the biggest challenges in managing head and neck cancers, especially salivary gland cancers, is the identification of secreted biomarkers of the disease that can be evaluated noninvasively. A relevant source of enriched tumor markers could potentially be found in the tumor secretome. Although numerous studies have evaluated secretomes from various cancers, the influence of the cancer secretome derived from salivary gland cancers on the behavior of normal cells has not yet been elucidated. Our data indicate that secretome derived from salivary gland cancer cells can influence the expression of two potential biomarkers of oral cancer—namely, bone sialoprotein (BSP) and dentin sialoprotein (DSP)—in normal salivary gland cells. Using routine immunohistochemistry, immunofluorescence, and immunoblotting techniques, we demonstrate an enrichment of BSP and DSP in human salivary gland (HSG) cancer tissue, unique localizations of BSP and DSP in HSG cancer cells, and enriched expression of BSP and DSP in normal salivary gland cells exposed to a cancer secretome. The secretome domain of the cancer microenvironment could alter signaling cascades responsible for normal cell proliferation, migration, and invasion, thus enhancing cancer cell survival and the potential for cancer progression. The cancer secretome may be critical in maintaining and stimulating “cancer-ness,” thus potentially promoting specific hallmarks of metastasis.
Introduction
The secretome of any group of cells was originally defined to describe the secreted factors, as well as the individual components of the secretory pathway. 1 We now refer to the secretome as being made up of only the secreted factors, specifically, “the global group of proteins secreted into the extracellular space by a cell, tissue, organ, or organism at any given time and condition, through known and unknown mechanisms involving constitutive and regulated secretory organelles.” 2 In addition to proteins, the secretome also contains non-protein constituents, including lipids, micro RNAs, and messenger RNAs, and plays an important role in cell–cell communication that promotes normal cellular function. Analyses of secretomes have revealed that chronic diseases often exhibit changes in their secretome composition, and identification of specific proteins within a pathological secretome may be useful in diagnostics or as prognostic indicators.3-15 In cancer states, secreted factors remain key mediators of cell–cell communication, and alterations in the cancer secretome can be linked to cancer progression that is cell-signaling dependent.16-18 Similar to several other epithelial cancers, various salivary gland cancers have been linked to a family of proteins called SIBLINGs (Small Integrin Binding Ligand N-linked Glycoproteins), which is a group of five proteins, namely, bone sialoprotein (BSP), osteopontin (OPN), dentin matrix protein 1 (DMP1), dentin sialophosphoprotein (DSPP), and matrix extracellular phosphoglycoprotein (MEPE).19-40 Initially discovered as being specific to mineralized tissue such as bone and dentin,23,41-50 all five members of the SIBLING family have now been shown to be present in epithelial tumors, as well as in metabolically active ducts of the kidneys and salivary glands.51,52 The expression of SIBLING proteins in salivary gland cancer cells is known to enhance key cellular features that promote tumor growth and cancer progression via metastasis; SIBLINGs play critical roles in cell proliferation, adhesion, migration, and tumor progression of multiple cancer types, including salivary gland cancers.22,24,26-28,34 What remains unclear is the effect of the cancer secretome on the expression of SIBLING proteins in salivary gland cancers. The current study demonstrates for the first time that the expression of BSP and DSP in normal human salivary gland cells (HSG cells) is enhanced by the cancer secretome derived from human salivary gland cancer cells (HTB-41 cells).
Materials and Methods
Human Salivary Gland Tissue
Normal human submandibular salivary gland tissue (n=6) and cancerous human submandibular salivary gland samples (n=6) were acquired through the National Disease Research Interchange (NDRI; Philadelphia, PA). De-identified, cryofixed, autopsy, and/or biopsy samples received from NDRI were stored at −80C till experimentation. Normal and cancerous human submandibular salivary gland tissues (n=4) were acquired as tissue microarrays from US Biomax, Inc. (Rockville, MD) and stored at room temperature until immunohistochemistry procedures. A total of N=10 samples of normal and cancerous human submandibular salivary gland tissues were used for the current study.
Human Cell Lines
A normal HSG cell line was received as a generous gift (Dr. M. Hoffman, National Institutes of Health [NIH], Bethesda, MA). A submaxillary salivary gland cancer cell line, HTB-41, was acquired from the American Type Culture Collection (ATCC; Manassas, VA). HSG and HTB-41 cells were aseptically cultured in DMEM/F-12 and McCoys5A media, respectively (Corning Cellgro; Manassas, VA), supplemented with 10% FBS and 1% penicillin–streptomycin–amphotericin, in 5% CO2 atmosphere at 37C. All cell culture experiments were carried out in triplicate.
Antibodies
The following primary antibodies were used in the study: anti-BSP (1:100 for IHC and immunofluorescence [IF], 1:500 for WB/LFMb-25/#sc-73630; Santa Cruz Biotechnology, Inc., Dallas, TX), anti-DSP (1:100 for IHC and IF, 1:500 for WB/LFMb-21/#sc-73632; Santa Cruz Biotechnology, Inc.), and anti-tubulin (1:10,000 for WB/#sc9104; Santa Cruz Biotechnology, Inc.). The following secondary antibodies were used in this study: goat anti-mouse IgG antibody—horseradish peroxidase (HRP) conjugate (1:10,000 for WB/#12-349; Sigma-Aldrich, St. Louis, MO), and donkey anti-mouse IgG antibody—Dylight 488 (1:200 for IF; Jackson Immuno-Research Laboratories, Inc., West Grove, PA).
Cancer Secretome Collection
Cancer HTB-41 cells were aseptically grown in sterile 100 mm tissue culture dishes to 90% confluence in McCoys5A growth media. All growth media were replaced with serum-free DMEM/F-12 media for 24 hr, and serum-free conditioned media were collected, centrifuged at 10,000 rpm for 7 min, supernatant retrieved, and stored at −20C for use in secretome studies.53-55 Normal HSG cells cultured in the cancer secretome from HTB-41 cells were designated as HSG* (normal HSG cells cultured in cancer secretome) cells.
Immunohistochemistry
IHC procedures were carried out on both normal and cancerous human salivary gland tissue. Tissue sections were immunostained with specific antibodies against BSP and DSP. Briefly, tissue sections were rehydrated through decreasing concentrations of a graded ethanol series, endogenous peroxidase activity was quenched using 3% hydrogen peroxide (Bloxall; Vector Laboratories, Burlingame, CA), and sections were reacted against anti-BSP and anti-DSP primary antibodies (TX/1:100; Santa Cruz Biotechnology, Inc.). Sections were then conjugated with biotinylated secondary antibodies and reacted against an avidin–biotin complex (ABC Kit; Vector Laboratories). The ABC was then reacted against diaminobenzidine (DAB), an HRP enzyme for the specific antigen being tested. Sections were counterstained with hematoxylin, dehydrated through a graded ethanol series, mounted on glass slides, and sealed with glass coverslips. Cancer tissue sections treated with only the secondary antibody and counterstained with hematoxylin were used as negative controls. All IHC images were acquired using a light microscope (Leica DM2500; Leica Microsystems, Buffalo Grove, IL).
Western Blotting
HSG and HTB-41 cells were aseptically cultured in 100 mm sterile culture dishes until they reached 90% confluence. Total proteins were extracted using mammalian protein extraction reagent (M-PER; Thermo Scientific/Pierce, Rockford, IL). Total proteins were also extracted from HSG* cells. Protein estimation was carried out using the Reducing Agent Compatible and Detergent Compatible (RC DC) protein assay (Bio-Rad, Hercules, CA), and equal amounts of protein were resolved by 10% SDS-PAGE under reducing conditions. After electrophoresis, proteins were electrotransferred onto nitrocellulose membranes (Bio-Rad, Hercules, CA), blocked with 5% non-fat milk in 1× PBS, and probed with anti-BSP and anti-DSP antibodies (1:500; Santa Cruz Biotechnology, Inc., Santa Cruz, CA). HRP-conjugated goat anti-mouse IgG was used as secondary antibody (1:10,000; Sigma-Aldrich), and Clarity Western ECL (Bio-Rad, Hercules, CA) was used as the substrate for HRP detection.
To determine equal protein loading, each membrane was carefully washed, treated for 5 min with stripping buffer (Thermo Scientific/Pierce, Rockford, IL) to eliminate the previous reaction, and washed with PBS. Mem-branes containing whole protein were processed as above with antitubulin primary antibody (1:10,000; Santa Cruz Biotechnology, Inc., Santa Cruz, CA) and HRP-conjugated goat anti-mouse IgG secondary antibody (1:10,000; Sigma-Aldrich, St. Louis, MO). Chemiluminescent detection was carried out for HRP detection, as described above.
Immunofluorescence
HSG, HTB-41, and HSG* cells were aseptically grown on tissue culture grade glass coverslips in sterile 12 well plates. On reaching 80% confluence, cells were fixed in 3% paraformaldehyde, and processed for IF and confocal imaging. Fixed cells were permeabilized in 0.2% Triton X-100 and blocked with 10% blocking solution for 1 hr. Cells were incubated overnight at 4C, in anti-BSP and anti-DSP primary antibodies (1:100; Santa Cruz Biotechnology, Inc., Santa Cruz, CA). Following thorough washes with 0.07 M PBS, donkey anti-mouse secondary antibody (Dylight 488; 1:200; Jackson Immuno-Research Laboratories, Inc., West Grove, PA) was used to tag the specific proteins of interest. Cells were incubated with Rhodamine Phalloidin to visualize the actin cytoskeleton (1:50; Cytoskeleton, Inc., Denver, CO). Coverslips were mounted with mounting medium containing 4′,6-diamidino-2-phenylindole (DAPI) for nuclear counterstain (ProLong Gold; Life Technologies, Grand Island, NY). IF images were acquired using a Confocal microscope (Olympus FluoView FV300; Leeds Precision Instruments, Minneapolis, MN).
Results
BSP and DSP Are Enriched in Human Salivary Gland Cancers
IHC on human salivary gland cancer tissue showed enhanced positive reaction against BSP and DSP antibodies as compared with normal salivary gland tissue (Fig. 1). As expected, the stroma of normal salivary gland tissue showed a diffuse brown staining indicating the presence of BSP and DSP (Figs. 1A and 1B). In addition, epithelial cells lining the salivary gland ducts showed positive staining, indicating the presence of BSP and DSP (Figs. 1A and 1B—green asterisks).
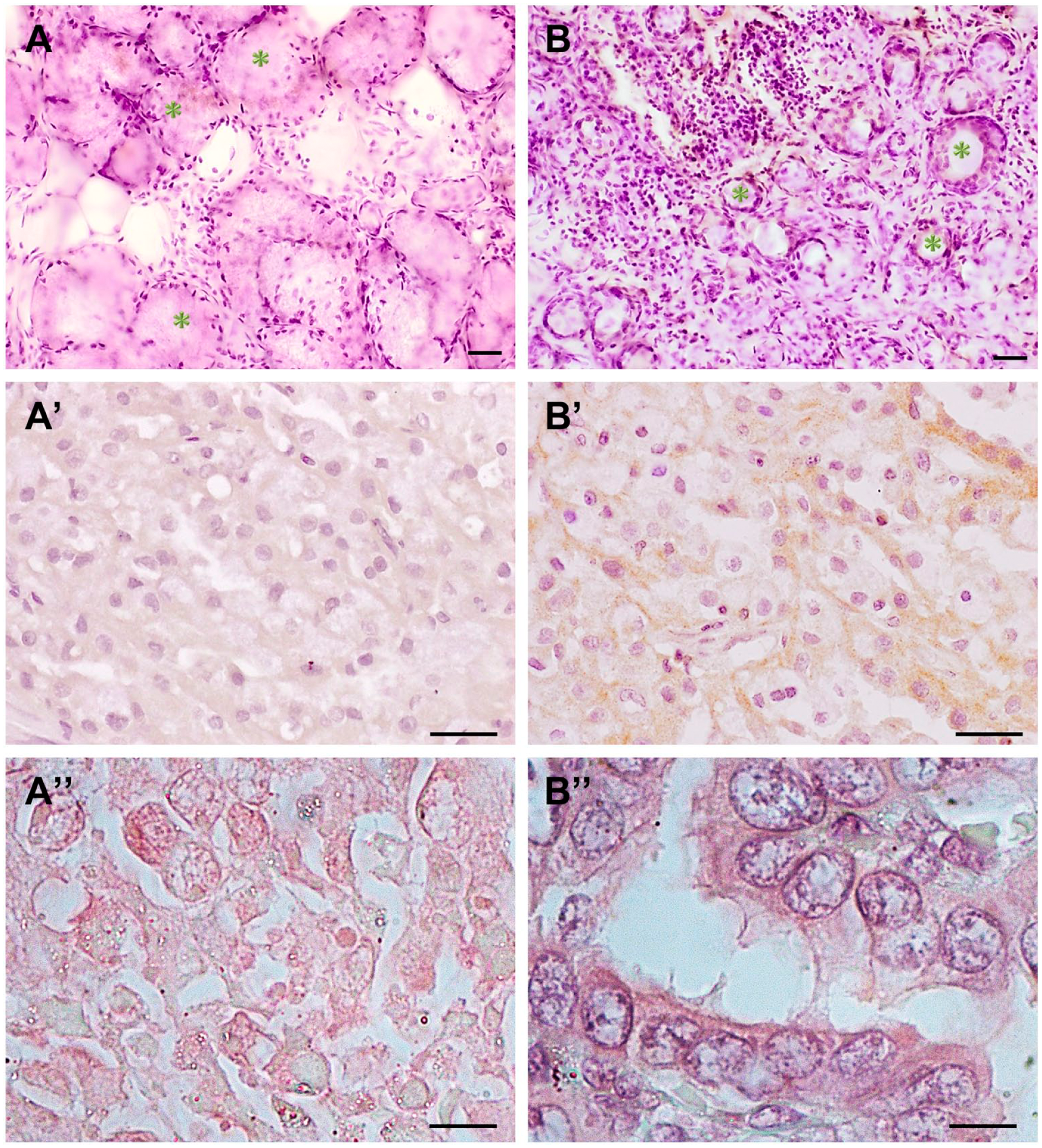
Bone sialoprotein (BSP) and dentin sialoprotein (DSP) are strongly expressed in human salivary gland cancer samples, as compared with normal human salivary gland tissue. All samples are sections of human submandibular salivary glands, and all cancer samples are examples of human submandibular adenocarcinoma. Scale bar = 20µm. A, A′, A″: Immunohistochemistry images showing BSP expression in normal (A), cancer (A′), and negative controls (A″). In normal tissue, nuclei stain dark purple, and there is no positive diaminobenzidine (DAB) staining evident within the intracellular profiles of the acinar cells. The cytoplasm of the ductal epithelial cells (green asterisks indicate stratified cuboidal epithelial ducts) show positive brown staining, indicating BSP expression. In cancer cells, there is diffuse cytoplasmic expression of BSP in epithelial cells, both ductal and acinar, as well as a diffuse positive DAB expression in the stroma/extracellular compartment. The cellular architecture is affected to the point where it is difficult to distinguish ductal cells from acinar cells. Negative controls (A″) did not show positive expression of BSP. B, B′, B″: Immunohistochemistry images showing DSP expression in normal (B), cancer (B′), and negative controls (B″). In normal tissue, DSP expression is seen in the ductal epithelia (green asterisks indicate stratified cuboidal epithelial ducts), as well as in the stroma/extracellular compartment. In cancer tissue, a very robust DSP expression is seen associated with epithelia. Purple nuclear staining is evident with brown staining of the cytoplasm present around the nucleus. Negative controls (B″) did not show positive expression of DSP.
In contrast to normal tissue, BSP showed a stronger reaction in the stroma as well as inside the cellular profiles of the ducts in cancer (Fig. 1A′—red asterisks). Cancer tissue also showed a robust staining against DSP in the stroma and within the duct profiles (Fig. 1B′). Tissue sections stained as negative controls did not show increased positive staining against BSP or DSP (Figs. 1A″ and 1B″).
Western blot analyses to evaluate levels of BSP and DSP in cancer HTB-41 cells as compared with normal HSG cells showed that levels of both SIBLING proteins were significantly elevated in cancer HTB-41 cells as compared with normal HSG cells (Fig. 4).
BSP and DSP Show Distinct Nuclear and Cytoplasmic Localizations in Human Salivary Gland Cancers
IF studies against BSP and DSP antibodies were carried out in normal HSG cells and cancer HTB-41 cells, and are depicted in Fig. 2. As expected, BSP and DSP were present in normal HSG cells. BSP specifically co-localized with the actin cytoskeleton and is seen expressed along the actin cytoskeleton, as well as outlining the cell profiles, co-localized with actin (Fig. 2A—merge). In cancer HTB-41 cells, BSP continues to co-localize with actin, and exhibited a presence along the actin stress fibers (Fig. 2—Panel B merge). DSP expression in normal HSG cells was cytoplasmic (Fig. 2C′). In addition, a discrete accumulation of DSP was seen localized as a cytoplasmic “cap” in relation to the nucleus (Fig. 2C″ and Panel C merge). In contrast, cancer HTB-41 cells showed a robust presentation of DSP in the cytoplasm, as a “perinuclear ring,” as well as translocation into the nucleus (white arrows—Figs. 2D′ and 2D—merge).
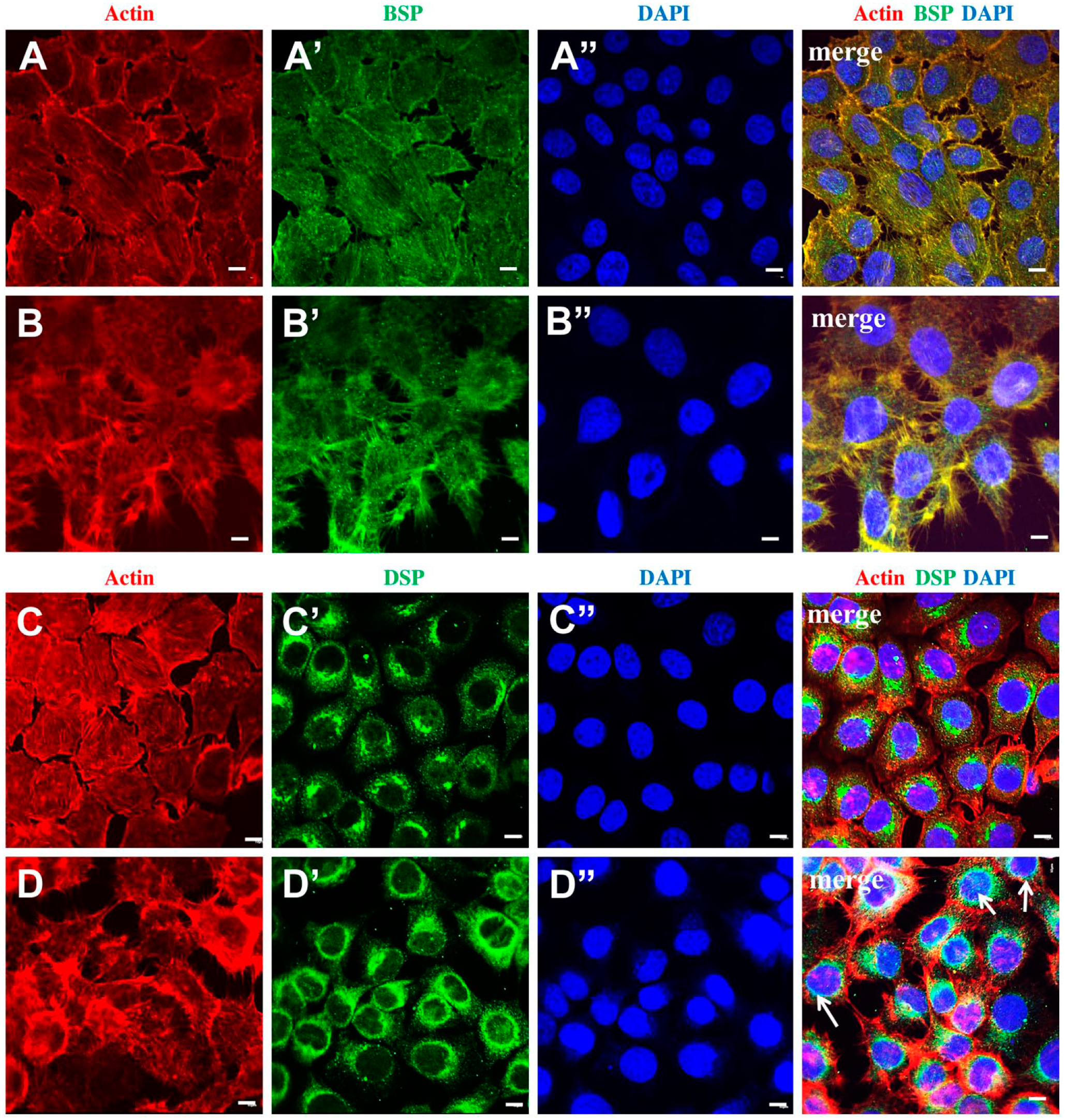
BSP and DSP exhibit specific intracellular localizations in normal HSG and cancer HTB-41 cells. Scale bar = 5 µm. A, A′, A″, merge: Representative confocal images depict actin cytoskeleton, intracellular localization of BSP, and nuclei in normal HSG cells. The actin cytoskeleton and cortical actin clearly show the normal, expected cell morphology of HSG cells. BSP is localized to the cell membrane as well as the cytoplasm. The merged image shows BSP co-localizing with actin, both in the cell membrane, as well as in the cytoskeleton. B, B′, B″, merge: Representative confocal images depict actin cytoskeleton, intracellular localization of BSP, and nuclei in cancer HTB-41 cells. Cell morphology in the cancer cells is different from that seen in normal salivary gland cells, as evidenced by the extensive actin-lined membrane extensions or filopodia, giving them a “stellate” appearance. BSP continues to be associated with the cell membrane and extends along the filopodia. In the merged image, the strong co-localization between actin and BSP is clearly evident. C, C′, C″, merge: Representative confocal images depict actin cytoskeleton, intracellular localization of DSP, and nuclei in normal HSG cells. In normal HSG cells, DSP is distributed in the cytoplasm, and there is an accumulation of DSP toward one side of the cell. The merged image clearly indicates that in addition to the diffuse cytoplasmic distribution of DSP, there is a cytoplasmic accumulation that is localized as a “cap” in relation to the nucleus. Unlike BSP, DSP is not associated with actin. D, D′, D″, merge: Representative confocal images depict actin cytoskeleton, intracellular localization of DSP, and nuclei in cancer HTB-41 cells. In cancer cells, DSP continues to have a cytoplasmic presentation, but the staining is more intense than in normal salivary gland cells. Unlike normal cells, the cytoplasmic accumulation of DSP is distributed as an intense “perinuclear ring.” Abbreviations: BSP, bone sialoprotein; DSP, dentin sialoprotein; DAPI, 4′,6-diamidino-2-phenylindole.
As evidenced by the actin distribution, the data also show that cancer salivary gland cells have a very distinct morphology as compared with normal salivary gland cells (Figs. 2B and 2D). Although normal cells exhibit a polyhedral shape, cancer cells exhibit a “stellate” configuration as a result of the elaborate filopodial extensions that project from the cells.
Cancer Secretome Enhances BSP and DSP Expression in Normal Human Salivary Gland Cells
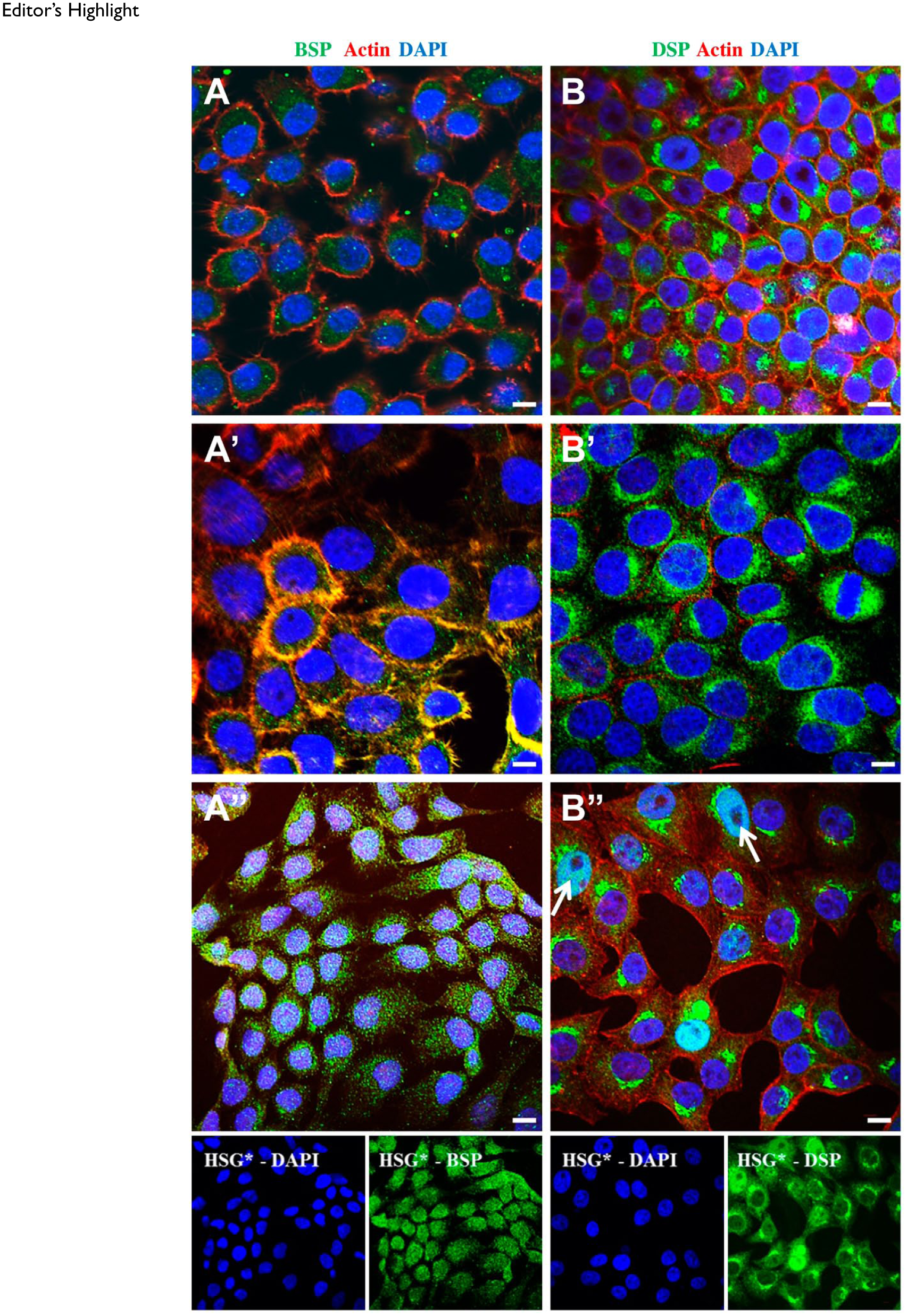
Under the influence of the cancer secretome, BSP expression seemed to be increased in the cytoplasmic and nuclear compartments of HSG* cells (Figs. 3A, 3A′, and 3A″). DSP, which showed the distinct “nuclear cap” presentation in normal HSG cells, showed a more robust presentation in the cytoplasm of HSG* cells, as well as nuclear translocation (white arrows—Figs. 3B, 3B′, and 3B″). The changes observed in BSP and DSP expression seem to indicate a gradual transition of HSG cells from a normal phenotype to a cancer phenotype under the influence of the cancer secretome.
Editor’s Highlight
Western blot analyses supported the IF data that under the influence of the cancer secretome, BSP and DSP expression was upregulated in normal HSG cells (Fig. 4).
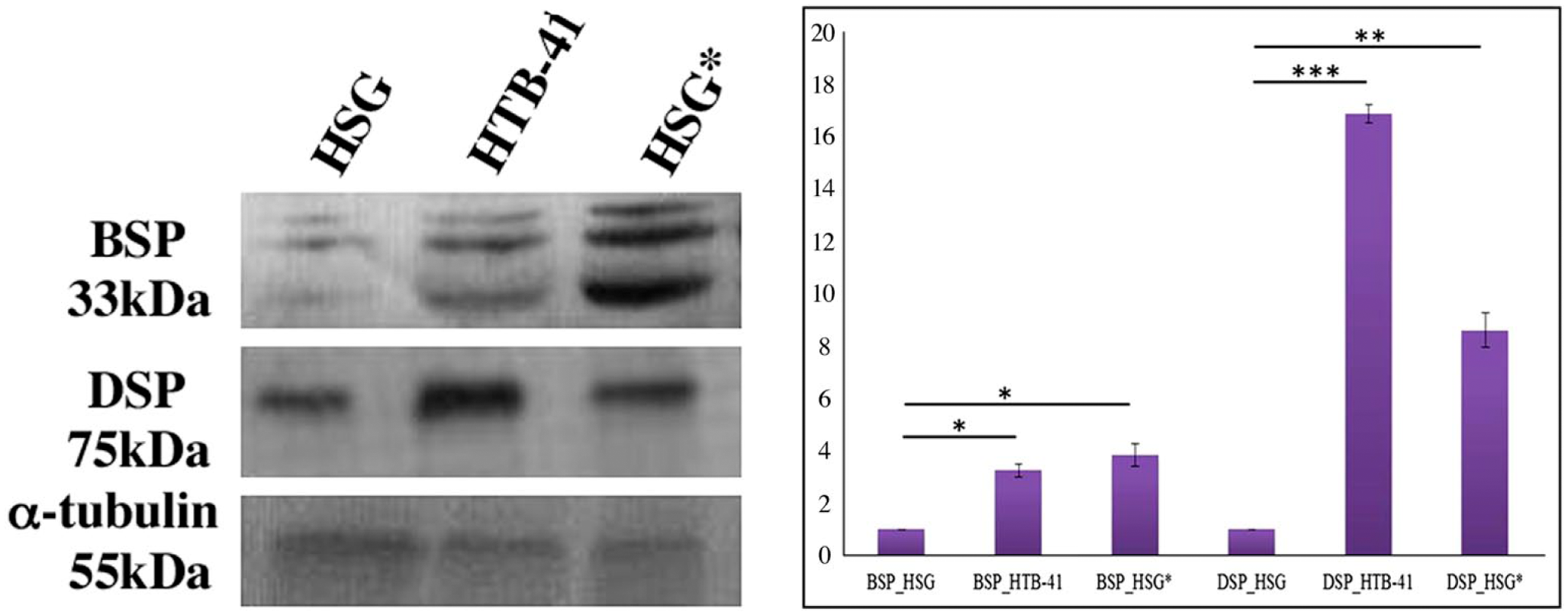
Evaluation of BSP and DSP expression in HSG, HTB-41, and HSG* cells in Western blot detection at 72 hr. Western blot revealed that BSP and DSP levels were elevated in HTB-41 cells, as well as HSG* cells as compared with their normal counterpart. Analyses of relative band intensities followed by a student’s t-test revealed that both BSP and DSP were significantly elevated in HTB-41 and HSG* cells compared with HSG cells (*p≤0.02, **p≤0.01, ***p≤0.001; data expressed as mean ± SD). Abbreviations: BSP, bone sialoprotein; DSP, dentin sialoprotein; HSG, human salivary gland; HSG*, normal HSG cells cultured in cancer secretome.
Discussion
SIBLINGs are a family of five proteins that are encoded by identically oriented tandem genes located on chromosome 4 (BSP, DSP, DMP1, OPN, and MEPE). They are small, soluble, RGD (Arg–Gly–Asp sequence) motif containing integrin-binding ligands, and are unique from larger extracellular matrix proteins such as fibronectin and collagen (Chen et al.).22,23,41-44,49,50,56-67,78 Four of the five members of this protein family (BSP, DSP, DMP1, and OPN) have been shown to be present in mineralized matrices of bone and dentin, whereas various combinations of all five SIBLING proteins have been shown to be significantly upregulated in various epithelial cancers, which metastasize to bone.19,20,23-27,30,34,35,39,51,68-76
A previous study evaluating human salivary gland tissue has shown that SIBLINGs are expressed in metabolically active, high energy-requiring sweat gland ducts as compared with metabolically inactive lacrimal ducts. 52 Major salivary glands produce approximately 700 to 1200 ml of saliva daily. Primary saliva that is produced by the acinar cells is isotonic as compared with plasma. The striated cells of the salivary gland ducts modify the primary saliva by removing sodium and chloride ions from it, and secreting potassium and bicarbonate ions into it. The altered secretion is hypotonic and is referred to as secondary saliva. To support this high-energy requiring metabolic activity of the ducts, salivary glands are supplied by a very rich vascular supply, almost 20 times as much as the blood flow to skeletal muscle. Our studies of normal human salivary gland tissue support the evidence that BSP and DSP are expressed in states of high metabolic activity that typically characterizes the ductal cells of normal salivary glands. Evaluation of the normal salivary gland cell line, HSG, also confirms that BSP and DSP have very specific and unique intracellular localizations. In addition, protein quantification in the current study also shows that there is not a complete lack of BSP and DSP in normal salivary gland cells; rather, a baseline level of BSP and DSP is maintained, presumably to regulate the extraordinarily high metabolic requirements of the salivary gland ductal cells.
In contrast to normal human salivary gland tissue and cells, our data show that salivary gland cancer exhibits a more robust expression of BSP and DSP. In vitro studies of breast cancer cells have shown that BSP is capable of increasing cell proliferation.72,75 Human breast cancer cells transfected with BSP and injected into the mammary fat pad of nude mice stimulated migration, invasion, and growth of primary and secondary tumors. 77 Elevated levels of BSP have also been found in cancers that have a high propensity to metastasize to bone, such as cancers of the breast,20,39,73,76,78 prostate,30,74,79,80 and lung.21,27,40,81 Our evaluations of human salivary gland cancer tissue as well as the human salivary gland cancer cell line HTB-41 indicate very specific intracellular expression of BSP. In tissue samples, BSP was found to be localized to the extracellular matrix/stroma and also had an intracellular presentation in both acinar and ductal cells. Within both acinar and ductal cells, BSP had a very diffuse presentation, completely filling up the cellular profiles, with the nucleus being prominently visible. IF studies of the cancer HTB-41 cell line mirrored the results from IHC, wherein BSP had a cytoplasmic expression. The cytoplasmic expression of BSP is consistent with that seen in other epithelial cell types, as well as non-epithelial cell types such as osteoclasts. 82 A feature of interest was that BSP seemed to co-localize with the actin cytoskeleton. Cancer cells exhibit an inherent capacity to migrate and invade into surrounding tissue, as well as into blood vessels to promote metastasis, and this process requires a dynamic rearrangement of the actin cytoskeleton to form migration-promoting cell membrane extensions like filopodia and invadopodia. It has been demonstrated that actin cytoskeleton organization is affected in BSP-null osteoclasts resulting in the formation of smaller actin-rich membrane extensions as compared with wild type osteoclasts.48,82 Therefore, the co-localization of BSP with the actin cytoskeleton could indicate a potential role for BSP in promoting cell spreading and initiating cell migration via filopodial extensions in human salivary gland cancers. Under the influence of the cancer secretome, normal salivary gland cells not only express significantly more DSP than the untreated cells but also exhibit elongated cell morphology unlike the typical polyhedral morphology of normal salivary gland cells. The data suggest that increased BSP and its potential interaction with the actin cytoskeleton may result in a more migratory cytoarchitecture under the influence of the cancer secretome.
DSPP and its cleaved products, namely, DPP (dentin phosphoprotein) and DSP, play important roles in biomineralization.47,49,50,58,83-87 Although cleaved from the same DSPP holoprotein, DPP and DSP are functionally distinct with respect to biomineralization. 88 DPP with its repeated sequences of aspartic acid (Asp) and phosphorylated serine residues (Pse), and relatively long carboxylate and phosphate groups, regulate the rate and location of dentin mineralization by apatite crystal formation.89,90-92 DSP, on the contrary, is primarily a glycoprotein with minimal phosphate, but very high levels of sialic acid and carbohydrate, 93 thus playing a role in initiation of mineralization and laying down of matrix. In addition to its expression in mineralized tissue such as bone, dentin, and cementum,44,49,50,58,83,85-87,94-96 DSP is found in non-mineralized tissues,52,97-99 and has also been shown to be elevated in several epithelial cancers, of particular note, prostate,24,30 breast,21,27 and lung. 27 Expression of DSP has also been related to aggressiveness in human prostate cancers and oral cancers.24,35 In addition, DSP has been implicated in the transition of oral epithelial dysplasia to oral squamous cell carcinoma, as well as potentially being a prognostic predictor of oral squamous cell carcinoma recurrence.34,100 Our data while they show a very unique cytoplasmic presentation of DSP in normal salivary gland cells as a “polar cap” in close proximity to the nucleus, in cancer cells, however, DSP does translocate into the nucleus. Under the influence of the cancer secretome, DSP shows nuclear translocation in normal salivary gland cells. Although mechanisms of this translocation are unclear, a recent report indicates that full-length DSPP can translocate into the nucleus in Human Embryonic Kidney Cell Line (HEK293), Dental Pulp Stem Cells (DPSC), and mouse osteoblastic cell line (MC3T3) cells. 101 In addition, Teti et al. 102 have showed DSP in the nuclei of human dental pulp cells in vitro, and speculated that DSP in the nucleus could regulate genes that are responsible for dentin matrix synthesis. 102 Although DSP has very specific roles in mineralized tissue, DSP expression in non-mineralized tissue such as salivary glands could indicate important roles, including regulation of cell adhesion and cell migration. A dysregulation of DSP expression and levels could therefore result in altered adhesion and migration dynamics that could potentially enhance cell invasion and eventual cancer progression. The specific partnering of DSP with its matrix metalloproteinase (MMP) partner (MMP-20) has been shown in oral squamous cell carcinomas, wherein DSP was co-expressed with MMP-20 in cytoplasmic, perinuclear, as well as intranuclear locations. 103 Our data support the growing evidence that DSP could potentially have a very distinct role, apart from its role in dentin matrix production, in non-mineralized tissue. Specifically, in tumorigenesis and cancer progression, DSP could function by promoting a cytoarchitecture conducive to cell motility and eventual cell invasion, as evidenced in our data by the “stellate” morphology seen in normal cells under the influence of the cancer secretome.
Several studies that have evaluated various cancer secretomes have revealed a variety of growth factors, proteases, cell motility factors, cytokines, chemokines, and cell surface receptors as their constituents. These secreted factors are key mediators of cell–cell communication and are responsible for regulating cell proliferation, cell migration, cell invasion, evasion of apoptosis, and several other hallmarks of cancer by cell signaling. The cancer secretome has multiple factors that can contribute toward various cellular functions, and our study supports the increasingly evident fact that cancer cells do not act in isolation during cancer progression and metastasis. Our data suggest that under the influence of the cancer secretome, BSP and DSP can potentially alter their cellular localizations, 55 thus indicating the cancer secretome’s potential to transform normal cells into their cancerous counterparts. Thus, in a patient, primary tumor cells have the potential to interact with non-transformed cells to create a microenvironment that is conducive to cancer cell survival, progression, and metastasis. As this close interaction between cancer cells and non-transformed cells can enhance cancer progression and increase malignancy, identifying key membrane and intracellular players such as the SIBLING proteins will allow us to develop strategic therapies targeting either specific factors in the cancer secretome or specific downstream molecules that are involved in the transmission of cell signals.
The cancer microenvironment is particularly suited to promote uncontrolled proliferation, migration, and invasion of cancer cells, thus promoting cancer progression and metastasis. The cancer secretome is an important domain of the cancer microenvironment that has the potential to influence the metastatic potential and local invasiveness exhibited by cancer cells. The gradual increase in BSP and DSP levels indicate that over time, accumulations of these SIBLING proteins could potentially alter signaling cascades that are responsible for normal cell proliferation, migration, and invasion, thus enhancing cancer cell survival and conferring increased metastatic potential. The current study is a first step toward specifically outlining intracellular localizations of these proteins under the influence of the cancer secretome. Although further studies are warranted and ongoing, the data show a clear role of the cancer secretome in maintaining and stimulating “cancer-ness,” thus potentially promoting specific hallmarks of metastasis, namely, cell proliferation, cell migration, cell invasion, and cell survival through downstream cell-signaling pathways mediated via BSP and DSP.
Footnotes
Acknowledgements
The authors acknowledge Dr. Adrianna Marcuzzi, PhD, Southern Illinois University School of Dental Medicine, for technical assistance.
Competing Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Author Contributions
SLH performed the immunohistochemistry and Western blotting. SLH helped in manuscript preparation. BF performed the immunofluorescence (IF). JCY performed the Western blotting and assisted with the IF. ASE and ARJ designed the study. ARJ supervised the study, analyzed data, and drafted the manuscript. All authors have read and approved the final manuscript before submission.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This study was supported by the 2014 Southern Illinois University Edwardsville Assigned Research Time Award (Joy: PI) and the 2014 Southern Illinois University School of Dental Medicine Pilot Project Award (Joy: PI).