
Abstract
Poor survival of high-grade glioma is at least partly caused by glioma stem-like cells (GSLCs) that are resistant to therapy. GSLCs reside in niches in close vicinity of endothelium. The aim of the present study was to characterize proteins that may be functional in the GSLC niche by performing immunohistochemistry on serial cryostat sections of human high-grade glioma samples. We have found nine niches in five out of five high-grade glioma samples that were all surrounding arterioles with CD31+ endothelial cells and containing cellular structures that were CD133+ and nestin+. All nine niches expressed stromal-derived factor-1α (SDF-1α), its receptor C-X-C chemokine receptor type 4 (CXCR4), osteopontin and cathepsin K. SDF-1α plays a role in homing of CXCR4+ stem cells and leukocytes, whereas osteopontin and cathepsin K promote migration of cancer cells and leukocytes. Leukocyte-related markers, such as CD68, macrophage matrix metalloprotease-9, CD177 and neutrophil elastase were often but not always detected in the niches. We suggest that SDF-1α is involved in homing of CXCR4+ GSLCs and leukocytes and that cathepsin K and osteopontin are involved in the migration of GSLCs out of the niches.
Keywords
Introduction
In high-grade glioma, only a fraction of the malignant cells are glioma stem-like cells (GSLCs). These cells are held responsible for tumor maintenance (Hjelmeland et al. 2010; Lathia et al. 2010). GSLCs are highly resistant to therapy because they have an active G2/M checkpoint for DNA repair when damage is induced by irradiation or chemotherapy (Mir et al. 2010) and they express ABC transporters (Rama et al. 2014); this results in recurrence of the tumor (Brooks et al. 2013; Cruceru et al. 2013; Gatti et al. 2013; Hjelmeland et al. 2010; Huse and Holland 2010; Kim et al. 2013; Lathia et al. 2011; Lathia et al. 2012; Lim et al. 2014; Niibori-Nambu et al. 2013; Rao et al. 2013; Schiffer et al. 2014).
GSLCs reside in a specific microenvironment referred to as the GSLC niche, which is considered to be a dynamic and complex milieu in which GSLCs with self-renewal capacities proliferate (Borovski et al. 2011; Brooks et al. 2013; Cheng et al. 2013; Hjelmeland et al. 2010; Kim et al. 2013; Lathia et al. 2011; Lathia et al. 2012; Niibori-Nambu et al. 2013; Schiffer et al. 2014; Wang et al. 2013).
Previous studies have shown that GSLCs reside in close proximity to endothelial cells of blood vessel walls, which are indispensable for the formation of stem cell niches (Borovski et al. 2011; Brooks et al. 2013; Gilbertson and Rich 2007; Medema and Vermeulen 2011; Schiffer et al. 2014). GSLCs can modify the extracellular matrix and non-cancerous cells to create an appropriate microenvironment that facilitates proliferation, invasion and migration of GSLCs (Gilbertson and Rich 2007; Rao et al. 2013). However, it is still unknown which proteins exert what specific functions in the GSLC niche.
Therefore, the aim of the present study was to define GSLC niches that surround CD31+ endothelial cells and contain cells positive for the GSLC markers CD133 and nestin in human high-grade glioma tissue samples. In addition, we aimed to determine whether stromal-derived factor-1α (SDF-1α), its receptor C-X-C chemokine receptor type 4 (CXCR4), osteopontin (OPN) and cathepsin K (CTSK) are expressed in the GSLC niches.
CTSK is one of the highest-expressed proteases in high-grade glioma as appeared in an in silico search of gene expression in human high-grade glioma in publically available data sets, in human high-grade glioma tissue samples, and in high-grade glioma cell lines. The high expression of CTSK in high-grade glioma was validated by RT-qPCR, western blotting and immunohistochemistry (Verbovšek et al. 2014). CTSK can cleave and degrade SDF-1α in human bone marrow (Staudt et al. 2012). SDF-1α is a chemoattractant that is produced by stromal cells and is involved in homing of CXCR4+ stem cells/progenitor cells and leukocytes (Ehninger and Trumpp 2011; Teicher and Fricker 2010). This is evident in bone marrow, where SDF-1α retains CXCR4+ stem cells in stem cell niches. Upon stimulation with granulocyte colony-stimulating factor (G-CSF), macrophages that express the G-CSF receptor are reduced in number. Since macrophages promote the survival of stromal cells, reduction of the numbers of macrophages results in lower SDF-1α levels, which in turn results in mobilization of cells away from the niche (Ehninger and Trumpp 2011; Kucia et al. 2005). In high-grade glioma, hypoxia-inducible factor 1α (HIF-1α) and vascular endothelial growth factor (VEGF) upregulate SDF-1α and CXCR4 (Zagzag et al. 2008; Zagzag et al. 2006). Previous studies have shown that GSLCs express CXCR4 and that they are attracted by SDF-1α (Bleul et al. 1996; Duda et al. 2011; Kenig et al. 2010; Kryczek et al. 2007; Takabatake et al. 2009; Terasaki et al. 2011; Uemae et al. 2014; Wang et al. 2012; Zagzag et al. 2005).
OPN is highly expressed in high-grade glioma as an extracellular glycophosphoprotein with chemokine and cytokine properties that acts as a stem cell regulator in human bone marrow (Nilsson et al. 2005) and can induce hematopoietic progenitor cell mobilization in bone marrow in mice (Kollet et al. 2006). Besides, OPN can act as a survival factor in endothelial cells by interacting with the αVβ3 integrin (Atai et al. 2011; Mazzali et al. 2002; Rao et al. 2013; Sreekanthreddy et al. 2010). It has been shown that OPN increases vimentin expression in high-grade glioma cells, which is an intermediate filament protein and a marker for epithelial-to-mesenchymal transition (Jan et al. 2010; Lee et al. 2006). Furthermore, OPN promotes migration of cancer cells and leukocytes, such as neutrophils and macrophages, in high-grade glioma. These leukocytes secrete serine proteases, including neutrophil elastase (NE) but also matrix metalloproteases, such as MMP-9, and cathepsins. These proteases can all degrade extracellular matrix, resulting in increased invasiveness of cancer cells (Atai et al. 2011; Chantrain et al. 2008; Ehninger and Trumpp 2011; Jan et al. 2010; Mazzali et al. 2002; Mook et al. 2004).
We performed immunohistochemistry to molecularly characterize the GSLC niche in human high-grade glioma samples to visualize CD31 as an endothelial marker, CD133 and nestin as GSLC markers, CD68 as a macrophage marker, CD177 as a marker for activated neutrophils, MMP-9 that is secreted by macrophages (Mook et al. 2004) and NE secreted by neutrophils (Rudolphus et al. 1992) as well as SDF-1α, CXCR4, OPN, CTSK, VEGF, HIF-1α.
Materials & Methods
Patients
High-grade glioma samples from five patients were obtained from the Brain Tumor Bank maintained by the Department of Neuropathology at the Academic Medical Centre (AMC, Amsterdam, The Netherlands) and 1 glioblastoma multiforme (GBM) sample was obtained from the VU Medical Centre (VUmc, Amsterdam, The Netherlands). Research was performed on “waste” material that is stored in a coded fashion. Consent for this project was reviewed and waivered, and the project was approved by the Medical Ethics Review Committee of the Academic Medical Center and University of Amsterdam (reference number W14_224 # 14.17.0286). Consent for removal of the tissue and its storage in the tumor bank for research purposes was obtained and documented in the patients’ medical charts. The tumor samples obtained from the Brain Tumor Bank were derived from two males and three females. The age range was 38–74 years. Four patients had GBM (grade IV) and 1 patient had an anaplastic oligodendroglioma (WHO grade III). All five patients underwent surgical resection of the tumor and one patient was also treated with radiotherapy. Clinical information of the VUmc patient is lacking.
Human Brain Tumor Samples
Tissues samples were snap-frozen in liquid nitrogen in the operating room, and stored at -80°C until use. Cryostat sections (7-μm thick) were cut at -25°C on an HM560 cryostat (MICROM; Walldorf, Germany), picked up on glass slides, and stored at -80°C until use. All staining procedures, including those for controls, were performed on serial sections of each high-grade glioma sample. One GBM sample was not included in the analysis due to tissue damage and large areas of necrosis. Therefore, we excluded that sample and analyzed the remaining five high-grade glioma samples.
Immunohistochemistry
Two series of serial high-grade glioma cryostat sections were used. Sections were air-dried at room temperature for 15 min before immunohistochemical staining. The sections were fixed in acetone (-20°C) for 10 min and were air-dried afterward for 15 min. The sections were labelled and encircled with a PAP pen (Dako; Glostrup, Denmark), followed by 3 washing steps of 5 min with 0.1 M phosphate-buffered saline (PBS). The sections were treated with 100% methanol containing 0.3% H2O2 for 15 min to block endogenous peroxidase activity and to prevent non-specific background staining, followed by 3 washing steps of 5 min each using 0.1 M PBS. Then, sections were incubated with 0.1 M PBS containing 10% normal goat serum (Dako) and 0.1% bovine serum albumin (BSA) for 45 min to further reduce non-specific background staining. After tapping off the goat serum-containing buffer, sections were incubated overnight at 4°C with primary antibodies (see Table 1), except the sections that were incubated with CD68 and CD177; these were incubated for 2 hr (CD68) and 1 hr (CD177), both at room temperature.
Details of Primary Antibodies Used for Immunohistochemistry.
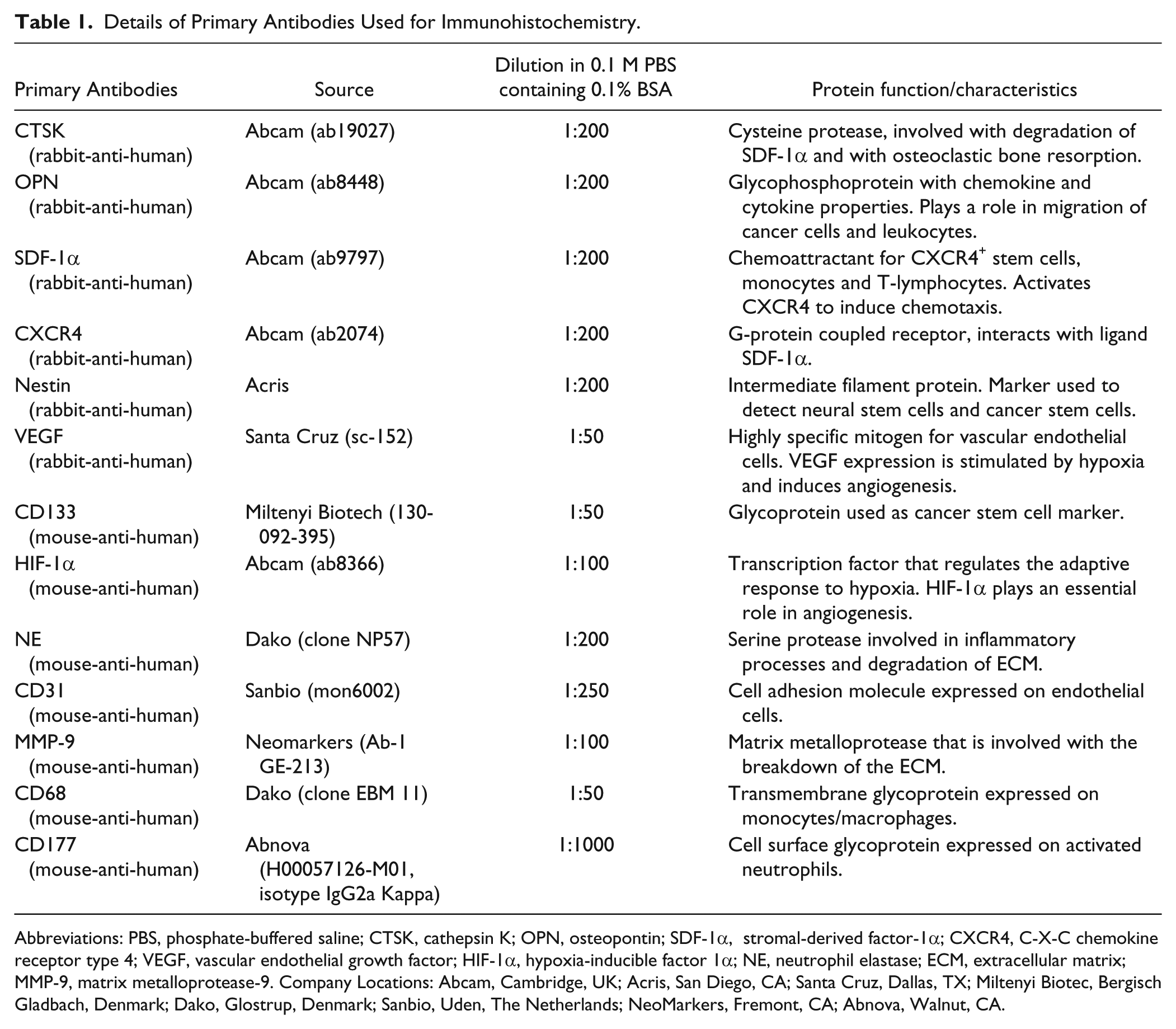
Abbreviations: PBS, phosphate-buffered saline; CTSK, cathepsin K; OPN, osteopontin; SDF-1α, stromal-derived factor-1α; CXCR4, C-X-C chemokine receptor type 4; VEGF, vascular endothelial growth factor; HIF-1α, hypoxia-inducible factor 1α; NE, neutrophil elastase; ECM, extracellular matrix; MMP-9, matrix metalloprotease-9. Company Locations: Abcam, Cambridge, UK; Acris, San Diego, CA; Santa Cruz, Dallas, TX; Miltenyi Biotec, Bergisch Gladbach, Denmark; Dako, Glostrup, Denmark; Sanbio, Uden, The Netherlands; NeoMarkers, Fremont, CA; Abnova, Walnut, CA.
After incubation with primary antibodies, the sections were washed 3 times for 5 min in 0.1 M PBS containing 0.1% BSA. Sections incubated with antibodies against CTSK, OPN, SDF-1α, CXCR4, nestin and VEGF were then incubated with polyclonal goat-anti-rabbit secondary antibody (Dako) in a 1:200 dilution in 0.1 M PBS containing 0.1% BSA for 1 hr. Sections incubated with antibodies against HIF-1α, CD31, CD68 and CD177 were then incubated with polyclonal goat-anti-mouse secondary antibody (Dako) in a 1:200 dilution in 0.1 M PBS containing 0.1% BSA for 1 hr. Sections incubated with antibodies against CD133, NE and MMP-9 were then incubated with Powervision goat-anti-mouse secondary antibodies for 1 hr (Immunologic; Duiven, The Netherlands). Incubation with secondary antibody was followed by 3 washing steps of 5 min with 0.1 M PBS. After that, the sections were incubated with AEC (peroxidase substrate kit; Vector Laboratories, Burlington, CA) for 20 min, followed by one washing step of 5 min using 0.1 M PBS. The sections were then placed in running tap water for 20 min and then in distilled water followed by incubation in hematoxylin (#51275; Fluka Biochemica, Sigma-Aldrich, St. Louis, MO) for nuclear staining for 3 min. The sections were again placed in running tap water for 20 min and then in distilled water. All incubation steps were performed at room temperature, except for the overnight incubations with primary antibodies that were performed at 4°C. Finally, the sections were covered with glycerin/gelatin mounting medium (Sigma-Aldrich).
Control incubations were performed in the absence of primary antibody. The second control panel was performed with rabbit serum or mouse serum in the same concentration as the primary antibody, to determine the effect of the serum on non-specific background staining (Suppl. Figs. S1 and S2).
Hematoxylin-eosin (H&E) Staining
High-grade glioma cryostat sections were air-dried for 15 min after removal from the freezer (-80°C). The sections were then fixed with Formol-Macrodex containing 4% formaldehyde, 7.2 mM CaCl2, 0.12 M Dextran-70, 0.12 M NaCl2 and 7.96 mM CaCo3, for 10 min, followed by a washing step in distilled water for 5 min. Nuclei were stained with hematoxylin for 3 min and the sections were then placed in running tap water for 20 min, after which the sections were placed in distilled water. Sections were then stained with eosin for 3 sec, dipped 5 times in distilled water, 15 times in 70% alcohol, 15 times in 96% alcohol and 10 times in 100% alcohol. Afterwards, the sections were rinsed three times for 5 min in xylene. The sections were covered with the synthetic mountant Pertex (#00801; Histolab, Götenburg, Sweden). All incubation steps were performed at room temperature.
Microscopic Analysis of Staining Patterns
Sections were analyzed by light microscopy and images were taken using the QWin software and the Leica DMLB microscope (Leica; Mannheim, Germany). The staining patterns in the first series of serial cryostat sections were analyzed by two independent observers (V.V.V.H. and W.T.) whereas the staining patterns in the second series of serial cryostat sections were analyzed by three independent observers (F.G., V.V.V.H and W.T.). Niches that expressed CD31, CD133, nestin, SDF-1α, CXCR4, OPN and CTSK were considered as GSLC niches. The expression patterns of the niches found in the first series of sections are shown in Table 2.
Expression of All Investigated Markers in the Five High-Grade Glioma Samples.
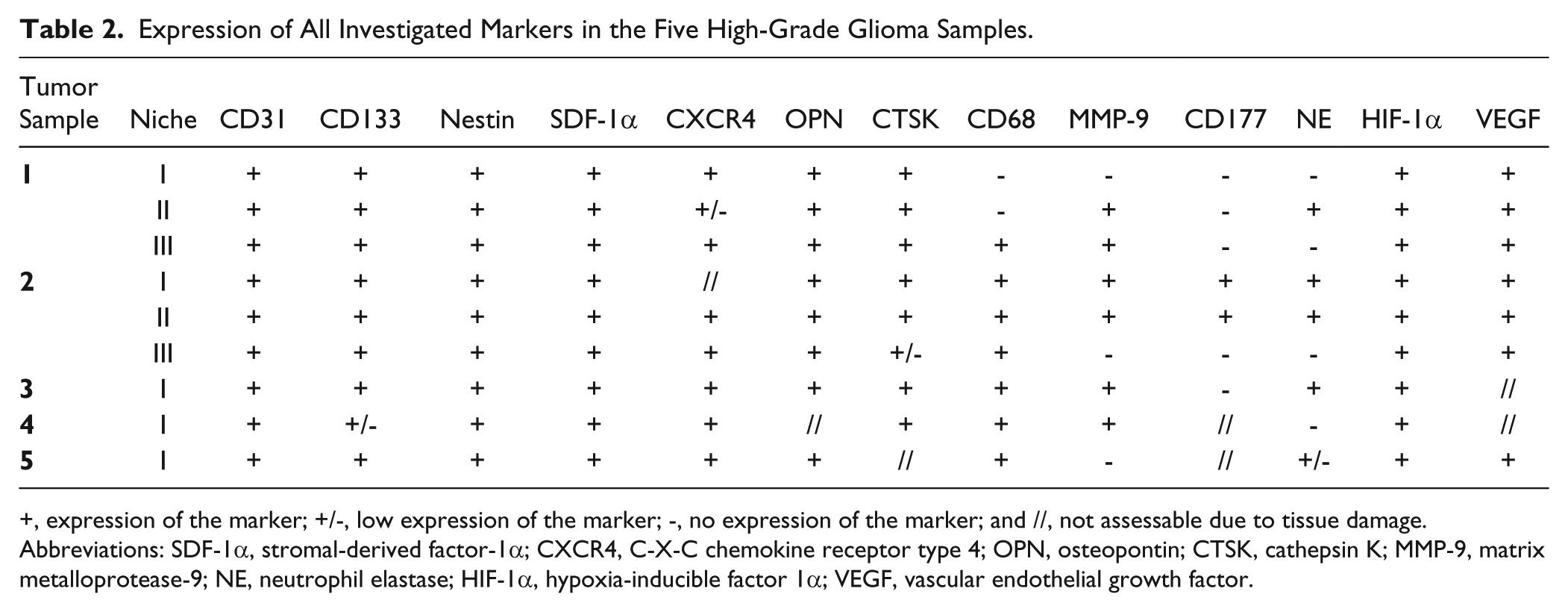
+, expression of the marker; +/-, low expression of the marker; -, no expression of the marker; and //, not assessable due to tissue damage. Abbreviations: SDF-1α, stromal-derived factor-1α; CXCR4, C-X-C chemokine receptor type 4; OPN, osteopontin; CTSK, cathepsin K; MMP-9, matrix metalloprotease-9; NE, neutrophil elastase; HIF-1α, hypoxia-inducible factor 1α; VEGF, vascular endothelial growth factor.
Results
Immunohistochemical Demonstration of Periarteriolar GSLC Niches
We performed immunohistochemistry on two series of cryostat serial sections of human high-grade glioma samples to determine the localization and, in particular, overlap of the protein markers listed in Table 1. The results obtained with the first series of sections are discussed here and were confirmed in the experiments with the second series of sections. H&E staining was performed for histological assessment of the high-grade glioma samples (Fig. 1A). In the high-grade glioma samples investigated, a high number of blood vessels were found (Figs. 1B, 2A, 2D, 2G). Larger blood vessels that appeared to be arterioles were surrounded by necrotic areas (Fig. 1A). Smaller blood vessels, such as capillaries, were not surrounded by necrotic areas (Fig. 2D). In 5 out of 5 tumor samples, we found 1–3 periarteriolar (CD31+, CD133+, nestin+) GSLC niches per 0.25 cm2 tissue and 9 periarteriolar GSLC niches in total. SDF-1α, CXCR4, OPN, CTSK, HIF-1α and VEGF were present in all periarteriolar GSLC niches. The leukocyte-related markers CD68, macrophage MMP-9, CD177 and NE were often expressed, but not in all GSLC niches. Table 2 indicates the expression of all markers in the five high-grade glioma samples. Seven sections from the first serial section series (8%) were not used for analysis because of tissue damage; but, in these cases, analysis was performed in images of sections from the second serial section series. The immunohistochemical images of the first serial section series are publically available at www.tumormetabolism.eu.
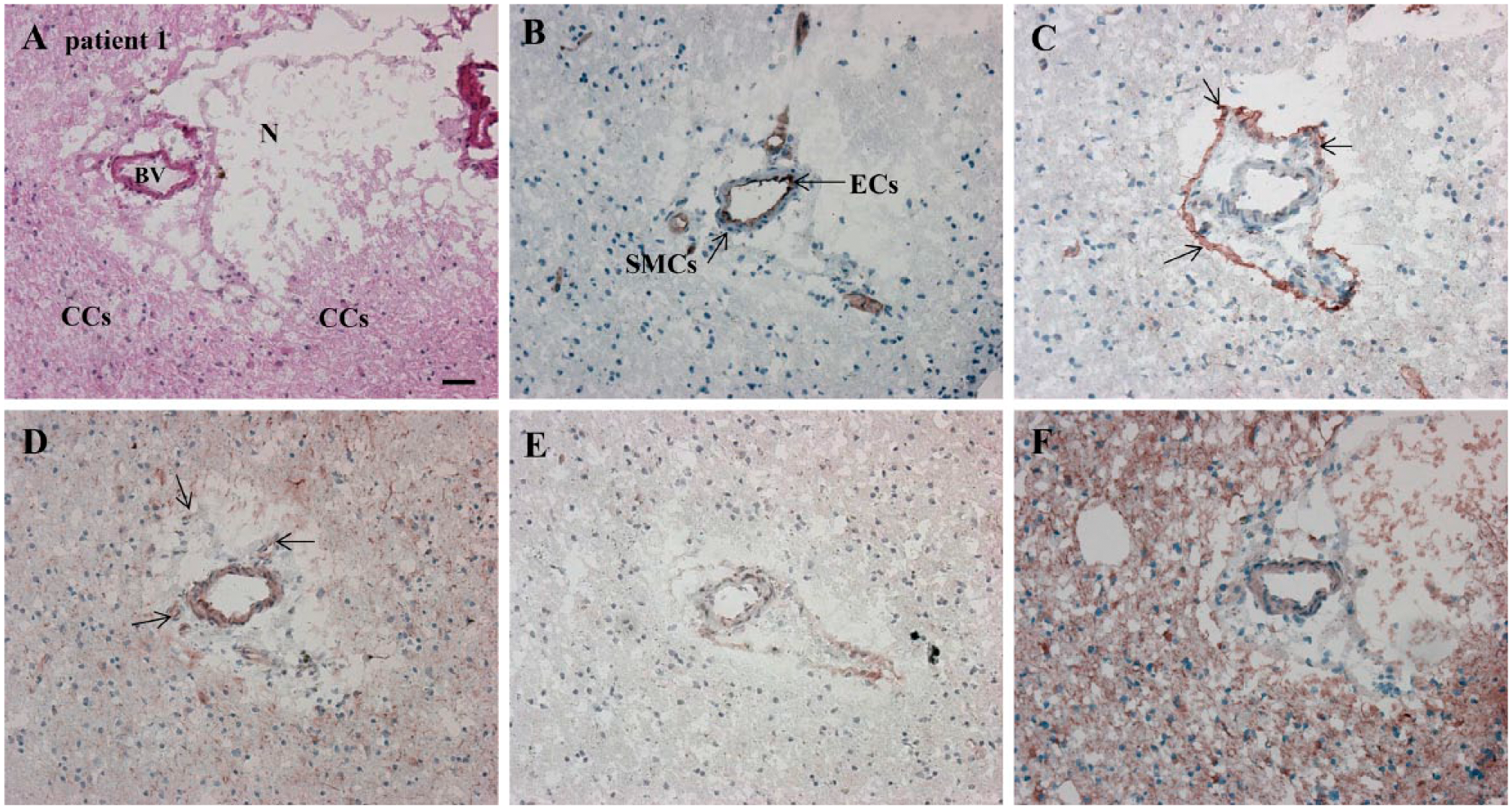
Serial cryostat high-grade glioma sections demonstrating the presence of CD133 and nestin around the arteriole and in the arteriolar wall. HIF-1α and VEGF staining confirms hypoxic conditions in and around the niche. N, necrosis; BV, blood vessel (arteriole); CCs, cancer cells; ECs, endothelial cells; SMCs, smooth muscle cells. (A) H&E staining for the overall histological assessment of the high-grade glioma section. Elements are hypoxic/necrotic regions, the unstructured tissue and blood vessels: (B) CD31 (cytoplasmic, in endothelial cells); (C) CD133 (on the cell membrane, around arterioles, in arteriolar walls and astrocyte-like cells); (D) Nestin (cytoplasmic, around arterioles, in arteriolar walls and astrocyte-like cells); (E) HIF-1α (intracellularly, in arteriolar walls and necrotic regions); and (F) VEGF (extracellularly in necrotic areas, in arteriolar walls and intracellularly in astrocyte-like cells). Scale, 30 µm. Abbreviations: HIF-1α, hypoxia-inducible factor 1α; VEGF, vascular endothelial growth factor.
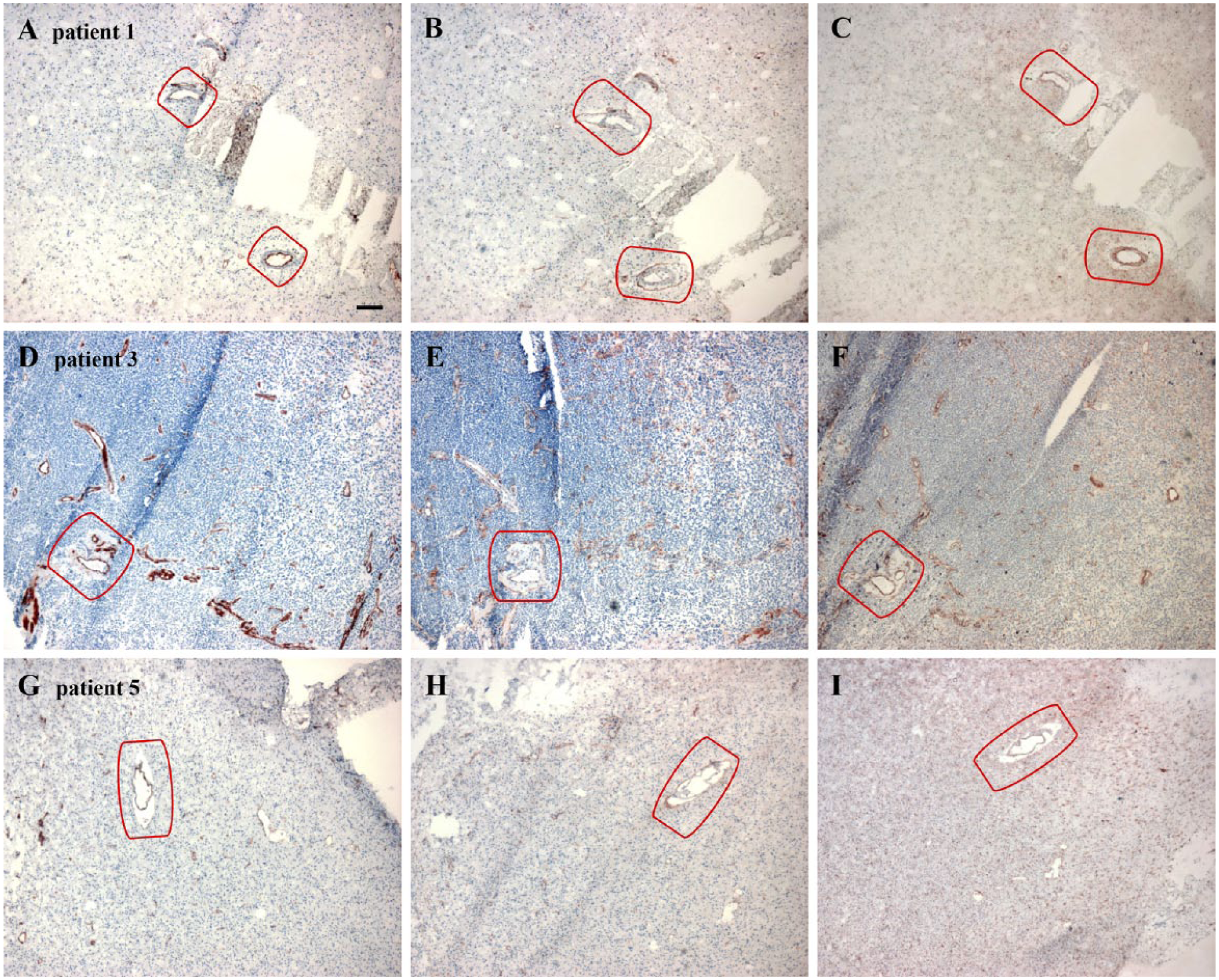
Overview images of CD31 (A, D, G), CD133 (B, E, H) and nestin (C, F, I) staining in periarteriolar glioma stem-like cell (GSLC) niches in three different high-grade glioma samples. The niches are marked in red. Images in (A–C) show two periarteriolar GSLC niches and images in (D–F) and (G–I) show one periarteriolar GSLC niche in the high-grade glioma samples. Scale, 120 µm.
CD31, CD133, Nestin, HIF-1α and VEGF in Periarteriolar GSLC Niches
Since the endothelium is an essential part of the GSLC niche, we sought to localize CD31 on endothelial cells. The endothelial cells were clearly positive for CD31 (Fig. 1B). Subsequently, we localized two markers to visualize GSLCs, CD133 and nestin, which were localized around CD31+ endothelium of arterioles in association with necrotic areas (Fig. 1C, 1D). However, CD133 and nestin were not exclusively localized around arterioles. CD133 staining was also present in astrocyte-like cells (Fig. 1C) around blood vessels, whereas endothelial cells were stained as well, but to a lesser extent (Fig. 1C). Nestin staining occurred even more than CD133 outside the niches in astrocyte-like cells and endothelial cells in all tumor samples (Fig. 1D). However, arterioles were always surrounded by necrotic areas and CD133+ and nestin+ cellular structures.
HIF-1α was present intracellularly and VEGF was present extracellularly, especially in necrotic regions and in blood vessel walls, thus confirming hypoxic conditions around CD31+ arterioles (Fig. 1E, 1F). VEGF was also present intracellularly in astrocyte-like cells (Fig. 1F). (See Suppl. Fig. S1A–S1C for the control incubations).
Overview images of CD31 (Fig. 2A, 2D, 2G), CD133 (Fig. 2B, 2E, 2H) and nestin (Fig. 2C, 2F, 2I) show periarteriolar GSLC niches in three different high-grade glioma samples. The GSLC niches were characterized by CD31+ arterioles with surrounding necrosis and surrounding CD133+ and nestin+ cellular structures (encircled in red). CD133 and nestin staining was found outside the niches as well. This suggests that CD133 and nestin are non-specific GSLC markers (See Suppl. Fig. S2 for the control incubations).
SDF-1α, CXCR4, OPN and CTSK in Periarteriolar GSLC Niches
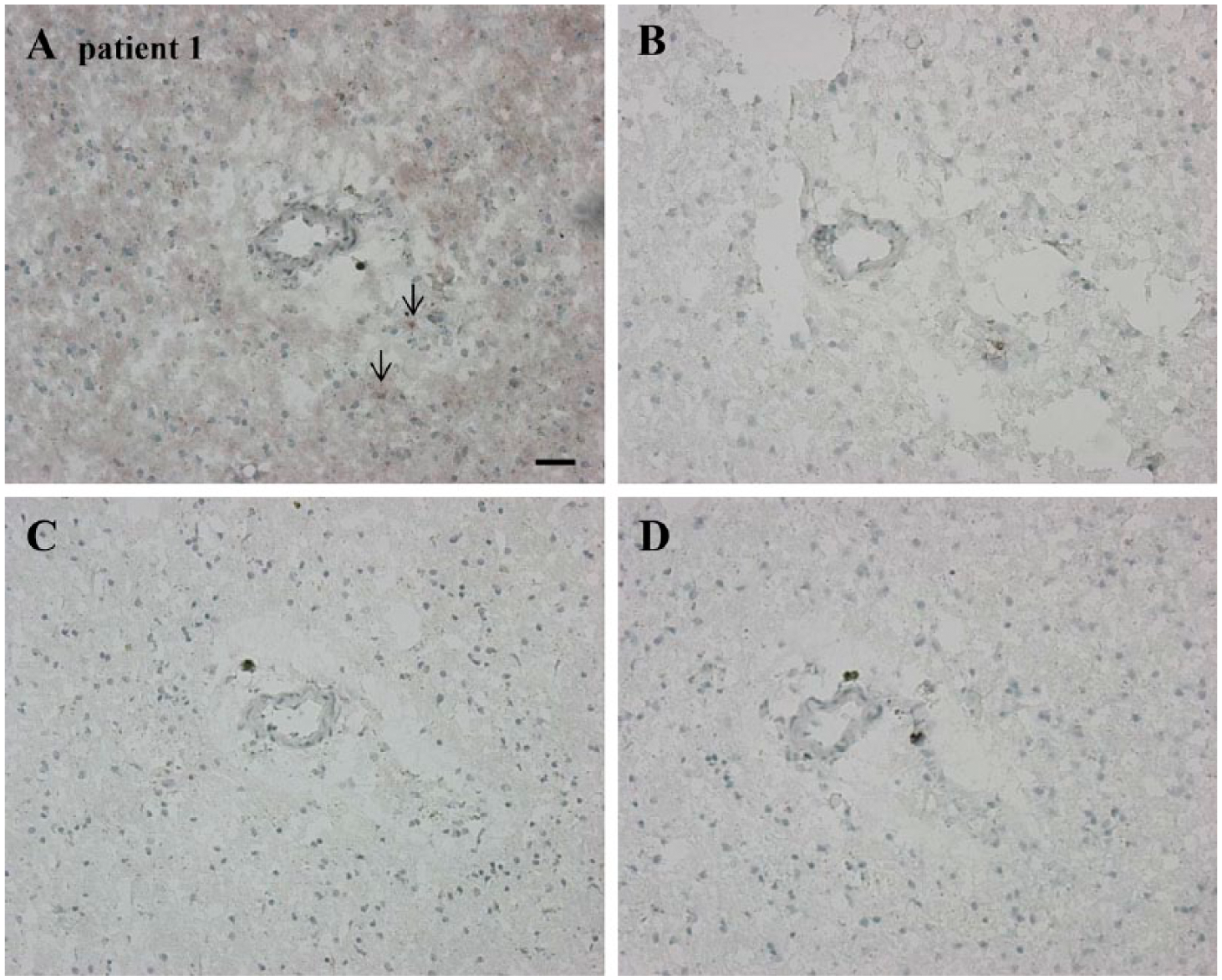
We localized chemokine SDF-1α and its receptor CXCR4 in the high-grade glioma sections (Fig. 3A, 3B). SDF-1α was present extracellularly and was abundantly present in arteriolar walls and necrotic areas (Fig. 3A). There was a clear overlap of SDF-1α, CD133, and nestin staining in GSLC niches (Figs. 1C, 1D, 3A). Similarly, CXCR4 staining overlapped CD133 and nestin staining in the niches (Figs. 1C, 1D, 3B). SDF-1α and CXCR4 staining overlapped in the periarteriolar niches (Figs. 1C, 1D, 3A, 3B). OPN was mainly present extracellularly in blood vessel walls and in necrotic areas and overlapped with CD133 and nestin staining in periarteriolar niches (Figs. 1C, 1D, 3C). CTSK was present in endothelium and overlapped with SDF-1α staining in periarteriolar niches (Fig. 3A, 3D). In addition, we found a correlation between high CTSK expression and low SDF-1α expression, and vice versa, in specific regions in GSLC niches in three different high-grade glioma samples (Figs. 3A, 3D, 4A–4D, 6F, 6H).
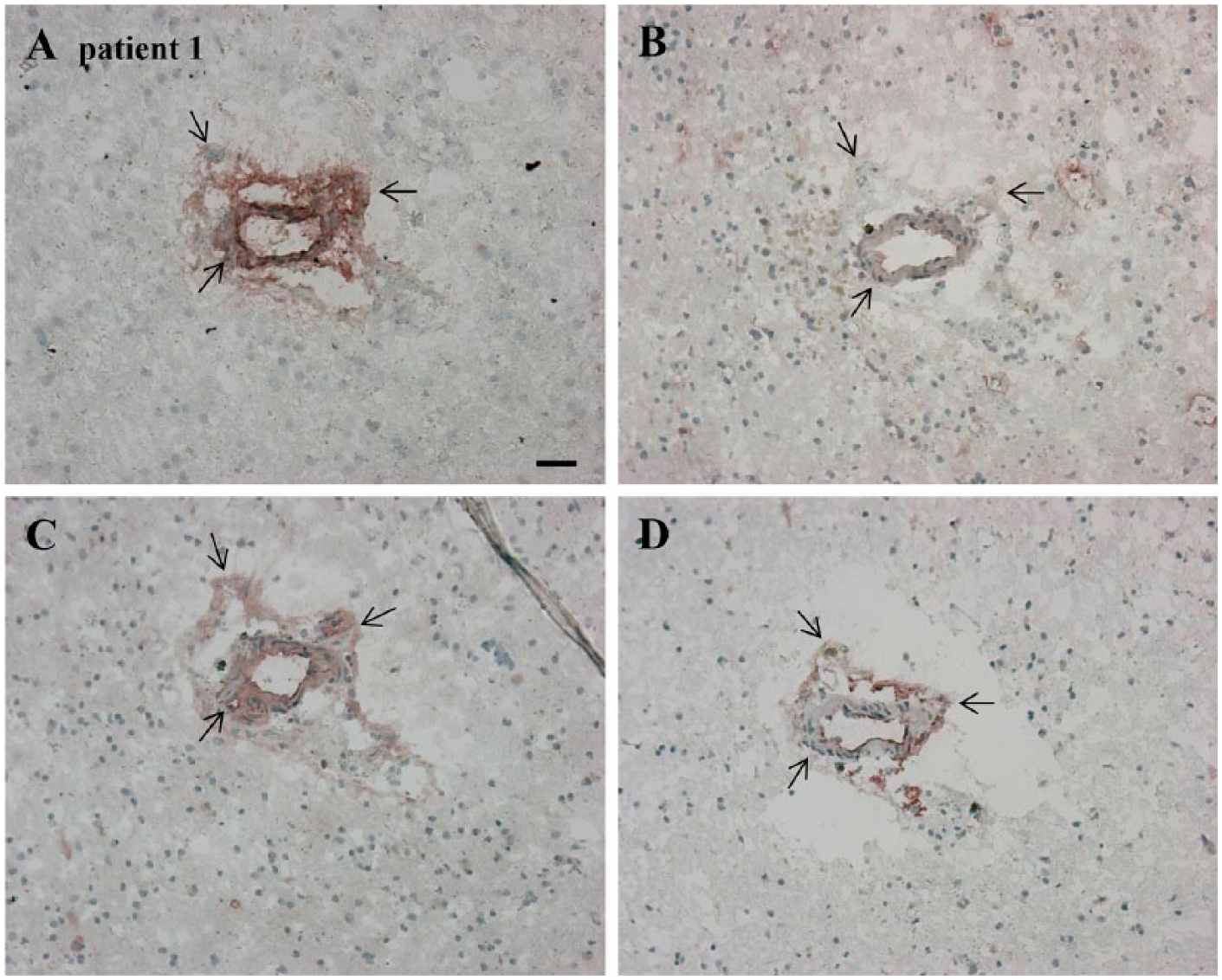
Serial cryostat high-grade glioma sections demonstrating expression and positive overlap of SDF-1α, CXCR4, OPN and CTSK in a periarteriolar glioma stem-like cell (GSLC) niche. Regions of overlap are indicated by black arrows. (A) SDF-1α (extracellularly, in arteriolar walls, necrotic areas). (B) CXCR4 (at cell membranes and in arteriolar walls). (C) OPN (extracellularly, in arteriolar walls, necrotic areas). (D) CTSK (mainly extracellularly and in endothelial cells). Scale, 30 µm. Abbreviations: SDF-1α, stromal-derived factor-1α; CXCR4, C-X-C chemokine receptor type 4; OPN, osteopontin; CTSK, cathepsin K.
Serial cryostat high-grade glioma sections demonstrating an association between low SDF-1α expression and high CTSK expression and vice versa in specific regions in two glioma stem-like cell (GSLC) niches. Low SDF-1α expression (A) and high CTSK expression (B) in a GLSC niche. High SDF-1α expression (C) and low CTSK expression (D) in a GSLC niche. Scale, 30 µm. Abbreviations: SDF-1α, stromal-derived factor-1α; CTSK, cathepsin K.
We also found three niches in which CD133 and nestin were not present around arterioles, but only in the arteriolar walls. In these niches, CD31, CD133, nestin, SDF-1α, CXCR4, OPN and CTSK staining overlapped in the arteriolar walls (data not shown).
Leukocyte-related Markers in Periarteriolar GSLC Niches
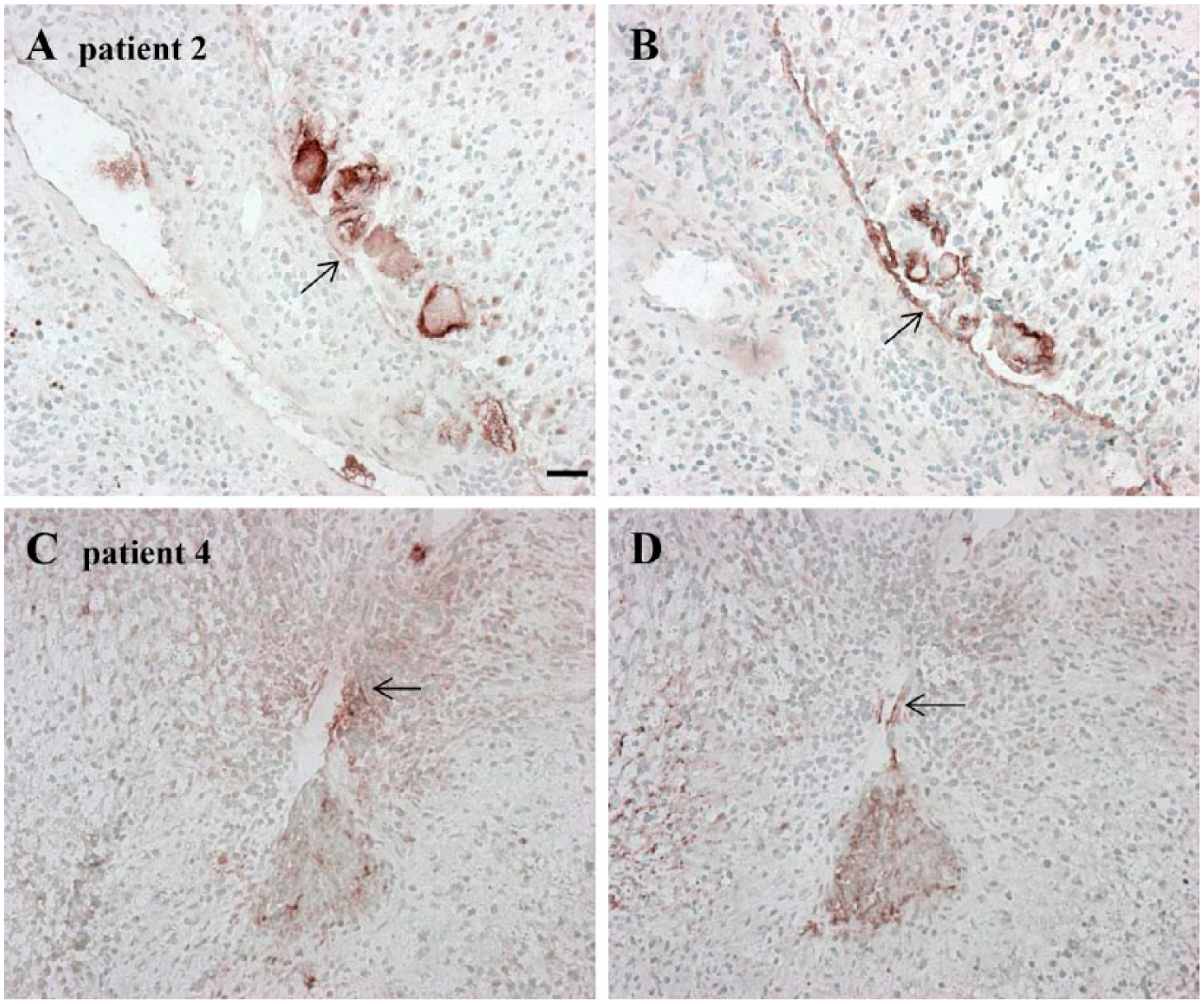
We next localized leukocyte-related proteins such as CD68 as marker for macrophages, macrophage MMP-9, CD177 as marker for activated neutrophils and NE. Macrophage MMP-9, CD177 and NE were not present in all high-grade glioma samples. Macrophage MMP-9 was expressed in 4 out of 5 samples, CD177 in 1 out of 3 samples and NE in 4 out of 5 samples (see Table 2).
These leukocyte-related markers were hardly present in some niches (Fig. 5), but were abundantly present in other niches (Fig. 6). Macrophages and macrophage MMP-9 were present in or in close vicinity to the arteriolar walls (Fig. 6B, 6C). Activated neutrophils and NE were also present in close vicinity of arteriolar walls (Fig. 6D, 6E). We found overlapping staining of macrophage MMP-9 and NE with SDF-1α (Fig. 6C–6F), of SDF-1α staining with CTSK staining (Fig. 6F, 6H), and of OPN staining with macrophages, macrophage MMP-9, activated neutrophils and NE (Fig. 6B–6E, 6G). The abovementioned markers overlapped with CD133+ GSLCs (Fig. 6A–6I). (See Suppl. Fig. S1D–S1F for the control incubations).
Serial cryostat high-grade glioma sections demonstrating the absence of leukocyte-related markers. (A) CD68 as marker for macrophages is hardly present (see black arrows). (B–D) Absence of macrophage MMP-9, CD177 and NE, respectively. Scale, 30 µm. Abbreviations: MMP-9, matrix metalloprotease-9; NE, neutrophil elastase.
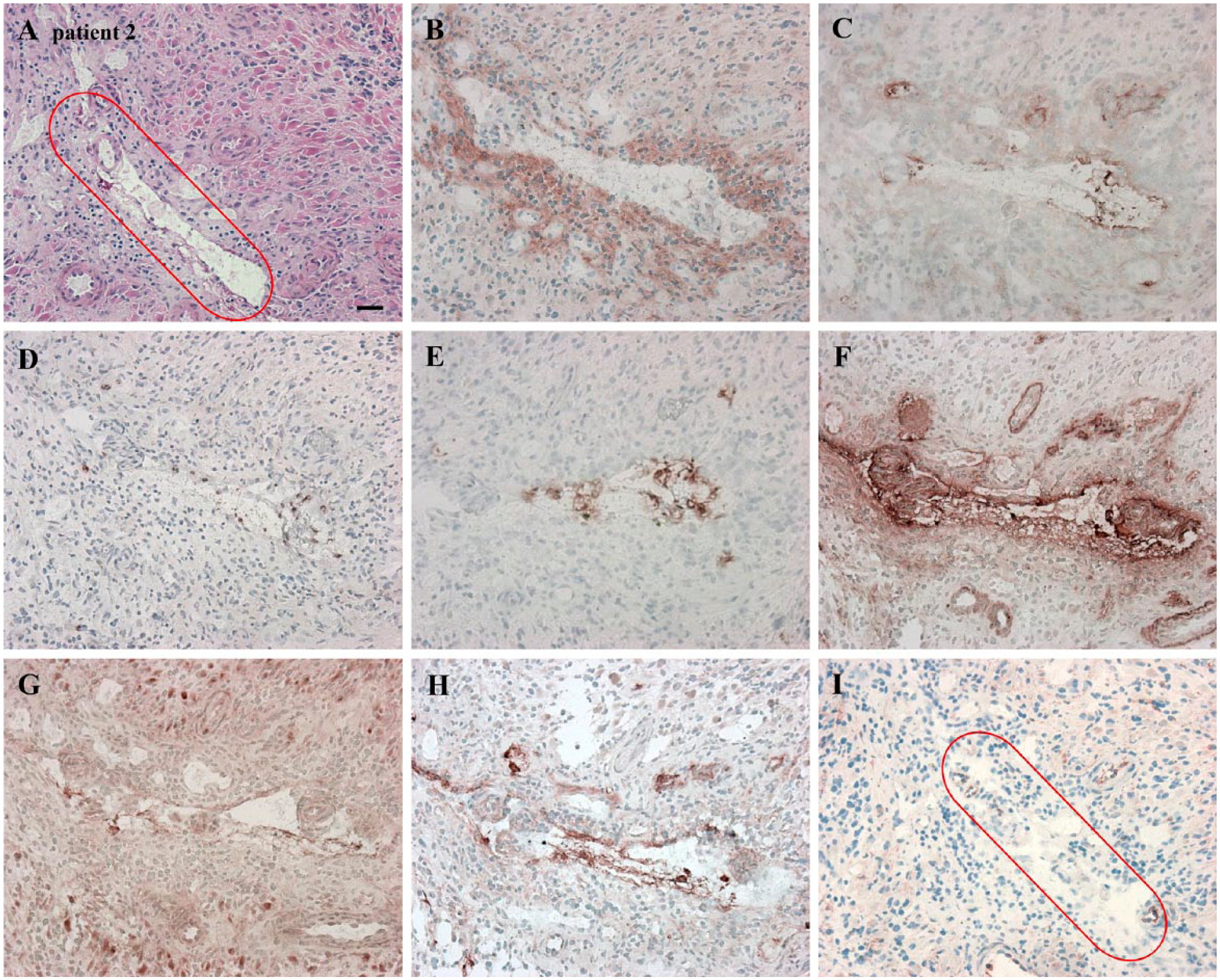
Serial cryostat high-grade glioma sections demonstrating leukocyte-related markers in another periarteriolar glioma stem-like cell (GSLC) niche, in which macrophages, macrophage MMP-9, activated neutrophils and NE were present. CTSK, macrophage MMP-9 and NE overlapped with SDF-1α and OPN overlapped with all leukocyte-related markers. The niche is marked in red. (A) H&E staining for the histological assessment of the section. (B) CD68 as marker for macrophages (on the cell membrane, in the vicinity of arterioles). (C) Macrophage MMP-9 (extracellularly, in arteriolar walls or in the vicinity of arterioles). (D) CD177 as marker for activated neutrophils (on the cell membrane, in the vicinity of arterioles). (E) NE (extracellularly, in the vicinity of arterioles). (F) SDF-1α (extracellularly, in arteriolar walls, necrotic areas). (G) OPN (extracellularly, in arteriolar walls, necrotic areas). (H) CTSK (mainly extracellularly and in endothelial cells). (I) CD133 (on the cell membrane and in arteriolar walls). Scale, 30 µm. Abbreviations: MMP-9, matrix metalloprotease-9; NE, neutrophil elastase; CTSK, cathepsin K; SDF-1α, stromal-derived factor-1α; OPN, osteopontin.
Discussion
We have visualized CD31+ periarteriolar GSLC niches in high-grade glioma samples, which are surrounded by CD133+ and nestin+ GSLCs and express SDF-1α, CXCR4, OPN and CTSK. Expression of leukocyte-related markers, such as CD68, macrophage MMP-9, CD177 and NE was not present in all GSLC niches. Our study supports the notion that SDF-1α, CXCR4, OPN and CTSK are associated with periarteriolar GSLC niches. The overlapping staining of SDF-1α and CXCR4 strengthens our hypothesis that SDF-1α functions in retaining CXCR4+ GSLCs in the GSLC niche, since SDF-1α is a chemoattractant for CXCR4+ cells (Ehninger and Trumpp 2011; Teicher and Fricker 2010). This process may be counteracted by CTSK-mediated degradation of SDF-1α (Staudt et al. 2012). Our observations that CTSK expression is high in regions where SDF-1α expression is low, and vice versa, support this hypothesis. However, immunohistochemistry does not allow the determination of CTSK activity in the high-grade glioma sections, so this is a subject for further mechanistic research. Figure 7 illustrates the molecular image of a periarteriolar GSLC niche to summarize our findings.
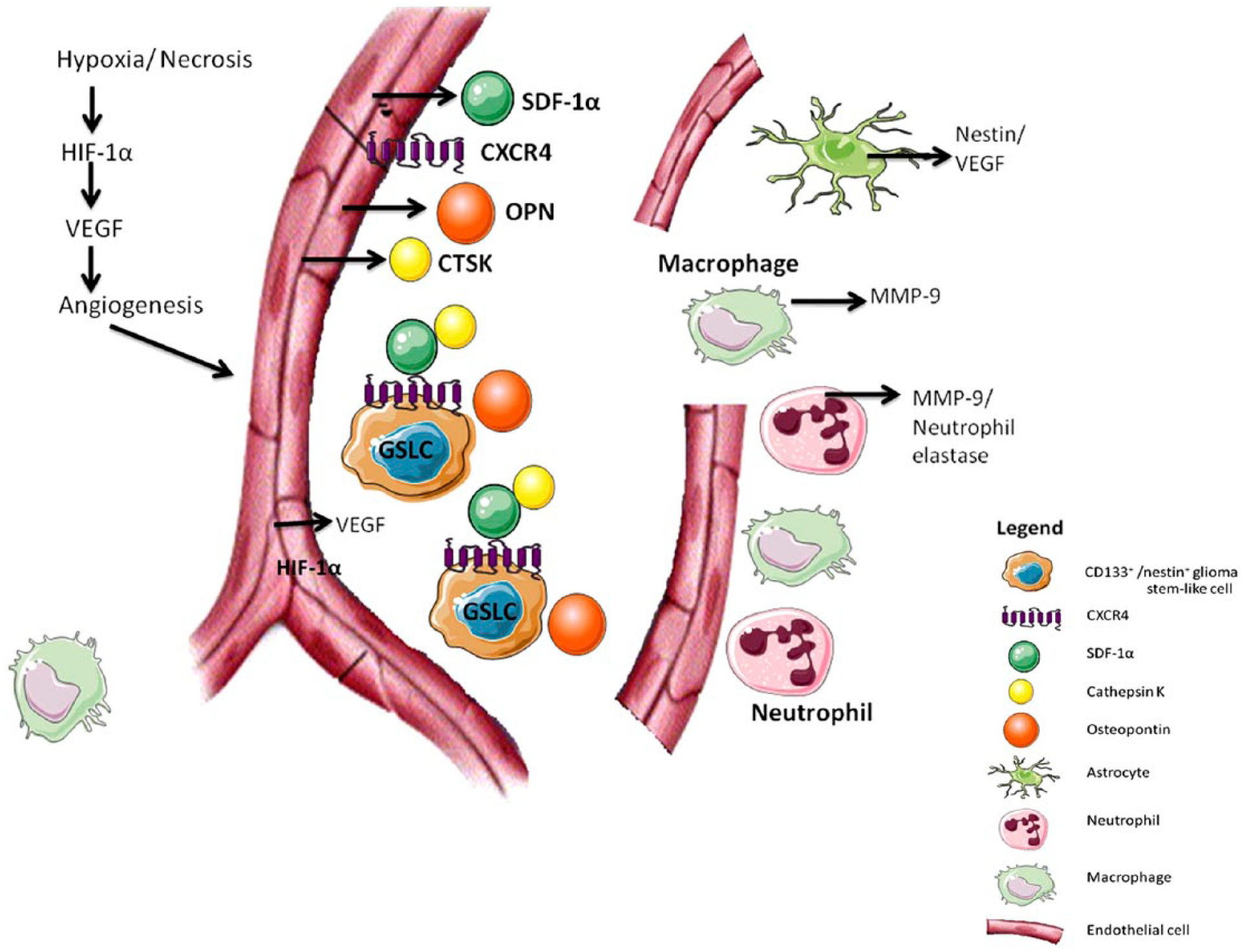
Model of a periarteriolar glioma stem-like cell (GSLC) niche. Hypoxia and necrosis result in the expression of HIF-1α and VEGF, which results in angiogenesis. SDF-1α, CXCR4, CTSK and OPN are functional proteins in the niche. The CD133+ and nestin+ GSLCs reside in close vicinity to CD31+ endothelial cells of arterioles. The GSLCs are CXCR4+ and are retained in the niche by the SDF-1α ligand, which binds to its CXCR4 receptor. OPN overlaps with the GSLCs and causes infiltration with neutrophils and macrophages. NE and macrophage MMP-9 are secreted by these leukocytes. The proteases CTSK, NE and macrophage MMP-9 overlap with SDF-1α and may in turn cleave SDF-1α, resulting in SDF-1α degradation and migration of GSLCs out of the niche. Around the niche, there are astrocyte-like cells that express nestin and VEGF. Abbreviations: HIF-1α, hypoxia-inducible factor 1α; VEGF, vascular endothelial growth factor; SDF-1α, stromal-derived factor-1α; CXCR4, C-X-C chemokine receptor type 4; CTSK, cathepsins K; OPN, osteopontin; NE, neutrophil elastase; MMP-9, matrix metalloprotease-9.
Our results indicate that OPN staining overlaps with CD133+ and nestin+ GSLCs and is present extracellularly, in blood vessel walls, and in necrotic areas. Since OPN functions as a stem cell regulator in stem cell niches in human bone marrow, OPN may have a similar stem cell regulatory function in GSLC niches. Kollet et al. (2006) showed that RANKL-stimulated bone-resorbing osteoclasts reduced OPN and SDF-1α levels in bone marrow in mice, which resulted in hematopoietic progenitor cell mobilization. This suggests that a reduction in OPN levels may result in GSLC mobilization out of the niches. Besides, OPN is a survival factor that may protect cells from apoptosis by OPN interaction with αVβ3 integrin of endothelial cells, which activates the pro-survival transcription factor NF-kB (Mazzali et al. 2002). This phenomenon may be the case for the CD133+/nestin+ GSLCs as well. If OPN protects these cells from undergoing apoptosis, OPN could be an important molecular therapeutic target. It has been shown that high-grade glioma patients with high OPN serum levels had poorer survival than patients with low OPN serum levels (9 months versus 22 months) (Sreekanthreddy et al. 2010). Similarly, a high expression of OPN in cervical carcinoma is associated with poor patient survival (Huang et al. 2015). Our data suggest that the protective effect of OPN on endothelial cells may be one of the causes of poor prognosis of high-grade glioma patients. This poor prognosis is also linked with the infiltration with neutrophils and macrophages via OPN in high-grade glioma. It is suggested that the infiltration with neutrophils and macrophages is induced by the interaction between α9β1 integrin and the OPN arginine-glycine-aspartate (RGD) sequence (Atai et al. 2011).
Interestingly, we found CD133 and nestin staining in astrocyte-like cells and endothelial cells in high-grade glioma sections. This may imply that CD133 and nestin are non-specific markers for cancer stem cells. A recent study validated a novel selective marker for GSLCs, the 25-kDa transmembrane protein CD9, which is over-expressed in some types of cancer and is able to mediate tumor cell invasion, apoptosis and resistance to chemotherapy (Podergajs et al., unpublished data). It was shown that CD9 plays an important role in the maintenance of stem cell characteristics. The validation further indicated that CD9 was expressed in GSLCs and glioma spheroids but not in neural stem cells (Podergajs et al., unpublished data). Another promising biomarker for GSLCs is the transcription factor SOX-2, which is found in neural progenitor cells and is also used as a GSLC validation marker (Molina et al. 2014). Our data, revealing CD133 and nestin as non-specific GSLC markers, underlines the need to discover more specific GSLC markers.
A striking aspect of our data is that we found expression of endothelial marker CD31 in arterioles, surrounded by necrosis and HIF-1α and VEGF expression. This can be explained by the fact that gas exchange does not occur between erythrocytes in the arterioles and surrounding tissues, but rather in the capillaries, resulting in necrosis around the arterioles in the high-grade glioma samples. It has been shown that hematopoietic stem cells (HSCs) also reside in hypoxic niches in bone marrow and, as a consequence, HSCs generate energy by maintaining a high rate of anaerobic glycolysis. Hypoxia appears to regulate hematopoiesis in bone marrow by maintaining important HSC functions, such as long-term self-renewal, cell cycle control, survival, metabolism, and protection against reactive oxygen species (Eliasson and Jonsson 2010; Suda et al. 2011). We suggest that hypoxia in the GSLC niches has similar functions.
A recent study mentioned that the subventricular zone (SVZ) contains the largest neural stem cell niche in the human adult brain (Gupta et al. 2014). It was shown that these stem cells migrate to brain tumors and may serve in this way as a reservoir of GSLCs. Evidence was given that irradiation of the SVZ had a positive effect on high-grade glioma patient survival. It was concluded by the authors that therapeutic targeting of the GSLC niche may have a positive clinical effect.
In conclusion, we have visualized CD31+, CD133+ and nestin+ periarteriolar GSLC niches in high-grade glioma samples from five different patients that all expressed SDF-1α, CXCR4, OPN and CTSK. Critically, there was clearly an overlap in the staining of SDF-1α and CXCR4, which suggests that SDF-1α retains CXCR4+ GSLCs in the GSLC niche, similar to the situation in stem cell niches in bone marrow. We also visualized overlapping SDF-1α and CTSK staining. In addition, we appreciated an association between high SDF-1α expression and low CTSK expression, and vice versa, in specific regions in GSLC niches, which suggests that CTSK cleaves SDF-1α in the GSLC niches, which may result in the migration of GSLCs out of the niches. Furthermore, the clearly overlapping staining of OPN and GSLCs suggests that OPN is involved in GSLC migration. On the other hand, the expression of the leukocyte-related markers, CD68, macrophage MMP-9, CD177 and NE, was not present in all niches.
Footnotes
Declaration of Conflicting Interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
R.J.M. was supported by an AMC PhD scholarship. T.T.L. was supported by program PI-0245, and U.V. by a young researcher fellowship of the Slovenian Research Agency.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.