
Abstract
Cytochemical staining remains an efficient way of identifying females who are heterozygous for the X chromosome-linked glucose-6-phosphate dehydrogenase (G6PD) gene. G6PD is highly polymorphic with certain alleles resulting in low intracellular G6PD activity in red blood cells. Low intracellular G6PD activity is associated with a risk of severe hemolysis when exposed to an oxidative stress such as fava beans, certain drugs and infections. Heterozygous females express the enzyme from both X-chromosome alleles resulting in two red blood cell populations each with G6PD enzyme characteristics representative of each allele; for example, normal and deficient. Cytochemical staining is the only way to determine the relative representation of each allele in red blood cells, a feature that is critical when assessing the risk for severe hemolysis when exposed to an oxidant such as the anti-malarial drug primaquine. This letter discusses red blood cell integrity with respect to the cytofluorometric assays for G6PD activity. An approach to making this test more robust is suggested. The approach makes this test more reliable and extends its use to a broader range of blood specimens.
In some malaria-endemic areas, up to 20% of the population carry variant alleles of the glucose-6-phosphate dehydrogenase (G6PD) gene (Von Seidlein et al. 2013). There are more than 160 diverse variants of G6PD, some of which result in decreased G6PD enzymatic activity. As the gene is on the X-chromosome, males are hemizygous and females are heterozygous or homozygous for the gene. X-chromosome inactivation (or ‘lyonization’) during embryonic development in which one or the other X chromosome becomes inactivated in every somatic cell results in the females essentially producing two red blood cell (RBC) populations, each containing one of the two allele G6PD enzyme products. A female with both a normal allele and a G6PD deficiency allele therefore produces one population of RBCs with normal G6PD enzyme activity and another with the G6PD enzyme that leads to RBCs that are predominantly deficient in G6PD activity. Individuals with G6PD deficiency are at risk of undergoing hemolysis if administered 8-aminoquinoline-based medicines, such as primaquine, which is the only drug available for the radical cure of Plasmodium vivax (P. vivax) malaria. Tafenoquine, another 8-aminoquinoline, is a promising compound (Llanos-Cuentas et al. 2014). Best clinical practice requires testing for G6PD deficiency prior to administering a high dose of any of these medicines. The evaluation of G6PD tests requires the availability of a repository containing blood samples with various types of G6PD deficiency (Kahn et al. 2013). Most tests measure activity in lysed blood, providing only the overall G6PD activity in a blood sample. However, cytochemical staining tests on intact cells or intracellular G6PD activity tests may, at times, be desirable to determine the proportion of deficient RBCs in heterozygotes (Van Noorden et al. 1982; Peters and Van Noorden 2009). Flow cytometry allows for the observation and stratification of RBCs by measuring intracellular G6PD activity (Van Noorden et al. 1989; Shah et al. 2012). In the case of heterozygote females, both the normal and deficient G6PD RBC populations can be observed by cytofluorometric assays (Van Noorden et al. 1989; Shah et al. 2012). The correct interpretation of the cytofluorometric test in terms of the proportions of normal and deficient RBCs is dependent on assay functionality and specimen integrity. The current study describes a formulation that restores the functionality of the cytofluorometric test when using preserved blood specimens. This methodology extends the utility of the cytofluorometric test to field samples and supports the construction of a robust specimen repository and associated quality-control protocols for the evaluation of tests for G6PD deficiency. It is in the context of the construction of a specimen panel that the integrity and stability of blood specimens collected with ethylenediaminetetraacetic acid (EDTA) anticoagulant were measured by both the whole lysate G6PD activity assay and the cytofluorometric assay at the time of collection, as well as after cryopreservation. This was done in the presence and absence of a specific formulation (additives).
All specimens were sourced through Bioreclamation, Inc (Westbury, NY) from voluntary donors after receiving signed consent under the institutional review board protocol 2010–017 IRB. Fresh EDTA blood specimens were divided into two equal portions. For one portion, the EDTA blood specimen was centrifuged at 2200 RPM for 5 min, the plasma and buffy coat removed and replaced with the same volume of additive (2.5% glucose, 0.9% sodium chloride, 0.27% adenine, 0.75% mannitol) (Preservation and Storage of Blood. c2005-2014). The other half of the sample remained without additive. Specimens were stored at 4°C. G6PD activity was characterized using the G6PD Kit (#345; Trinity Biotech PLC, Bray, Co Wicklow, Ireland), as well as the flow cytometry assay previously published (Shah et al. 2012) at several time intervals. For cryopreserved samples, if additives were used, they were washed from RBCs prior to cryopreservation and then replaced after thawing.
Stability data (Table 1) is presented for nine donors, two of whom were females with intermediate G6PD activities of 5.39 and 3.01 IU/gHb at Day 0, and with cytofluorometric profiles characteristic of females heterozygous for G6PD activity (two distinct cell populations). These two female donors showed high G6PD activity in 69% and 28% cells, respectively. A pairwise, one-sided Wilcoxon signed-rank test, assuming that overall and intracellular G6PD activity can only decrease in time, was used to test whether there was a significant difference between the aggregate assay result mean values at Day 0 and the days stored at 4°C.
G6PD Activity in EDTA Blood Specimens Stored at 4°C With and Without Additive.
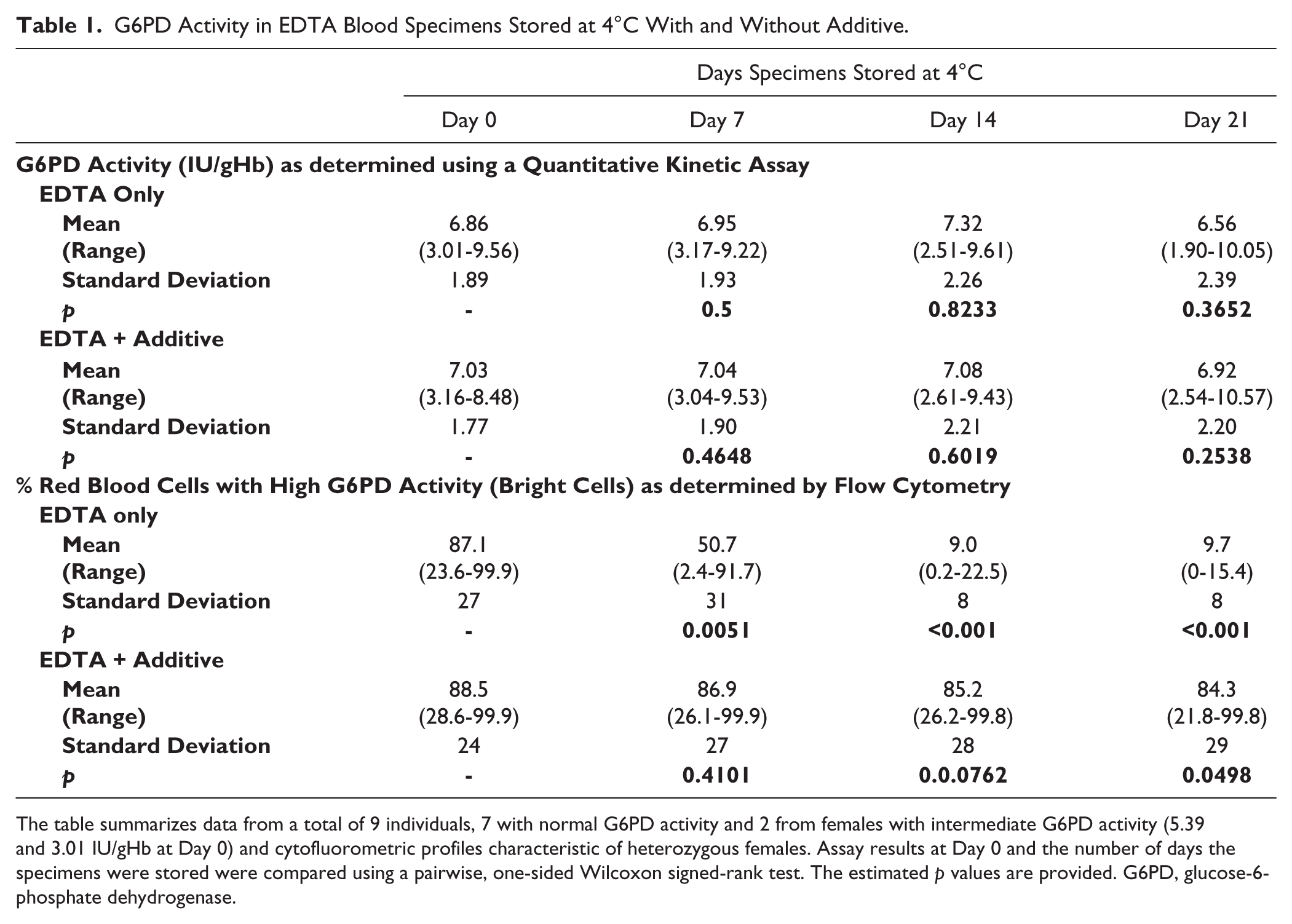
The table summarizes data from a total of 9 individuals, 7 with normal G6PD activity and 2 from females with intermediate G6PD activity (5.39 and 3.01 IU/gHb at Day 0) and cytofluorometric profiles characteristic of heterozygous females. Assay results at Day 0 and the number of days the specimens were stored were compared using a pairwise, one-sided Wilcoxon signed-rank test. The estimated p values are provided. G6PD, glucose-6-phosphate dehydrogenase.
The EDTA blood specimens stored at 4°C exhibited good stability in G6PD tests for which the blood was lysed and excess substrate added. This has been demonstrated previously (Castro et al. 2005) and shown here (Table 1), with the change in activity at Day 21 relative to Day 0 statistically not significant. The same stability was also observed post-cryopreservation (data not shown). In contrast, when the same specimens were observed for intracellular G6PD activity by flow cytometry, we observed a decrease in the proportion of RBCs with high G6PD activity (bright RBC), with almost no observable bright RBCs by Day 14 (Table 1 and Fig. 1). Similar relative trends and increasing disparity between the lysed blood G6PD quantitative results and the cytofluorometric assay results was observed in thawed cryopreserved specimens. Specimens from deficient donors remained unchanged as the cells were already low in G6PD activity (data not shown). These observations were consistently observed with all blood specimens.
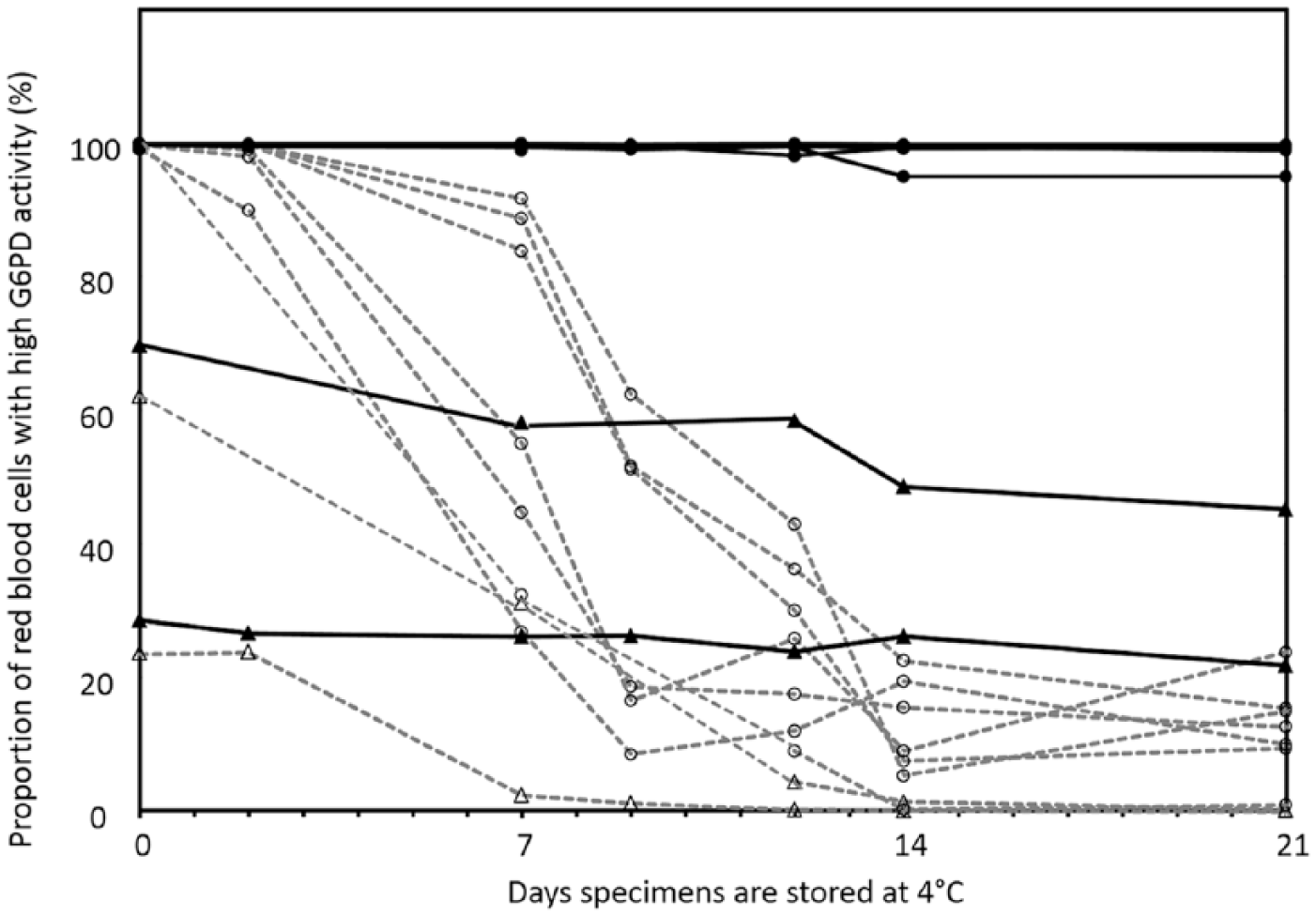
Specimen integrity using a cytofluorometric test for intracellular glucose-6-phosphate dehydrogenase (G6PD) activity. The same specimens, as indicated in Table 1, were submitted to a flow cytometry assay for intracellular G6PD activity. Red blood cells (RBCs) with high G6PD activity are fluorescently bright in this assay in contrast to RBCs with low G6PD activity. The graph shows the percentage of RBCs that were bright at various time intervals since blood collection. Specimens from donors with normal G6PD activity are represented by circles. The solid lines with solid circles represent the specimens with the additive, and dashed lines with empty circles without the additive. Specimens from female heterozygote donors with intermediate G6PD activity are represented by triangles. The solid lines with solid triangles represent the specimens with the additive, and dashed lines with empty triangles without the additive. Data is shown for two specimens from two donors heterozygous for G6PD and seven donors normal for G6PD.
When the blood specimens were supplemented with additive, the same normal and heterozygote donor samples stored at 4°C retained relatively unchanged proportions of bright (G6PD normal) cells over 21 days if the additives were introduced within 48 hr after collection (Table 1 and Fig. 1). With cryopreservation, the integrity again was dependent on the inclusion of additives (for a heterozygote sample, see Fig. 2 and Table 2). Illustrative stability data for specimens previously cryopreserved is shown for two donors, one with normal G6PD activity and a female heterozygous for G6PD (Table 2). We observed deterioration of the sample in the absence of additives, and this was less marked within 1 hr of treatment with the additives (Day 0 post-thaw), and continued to exhibit good integrity three days after thawing. Similar results were obtained for a G6PD normal sample (Table 2). The samples maintained integrity of intracellular G6PD activity for up to 22 days (data not shown).
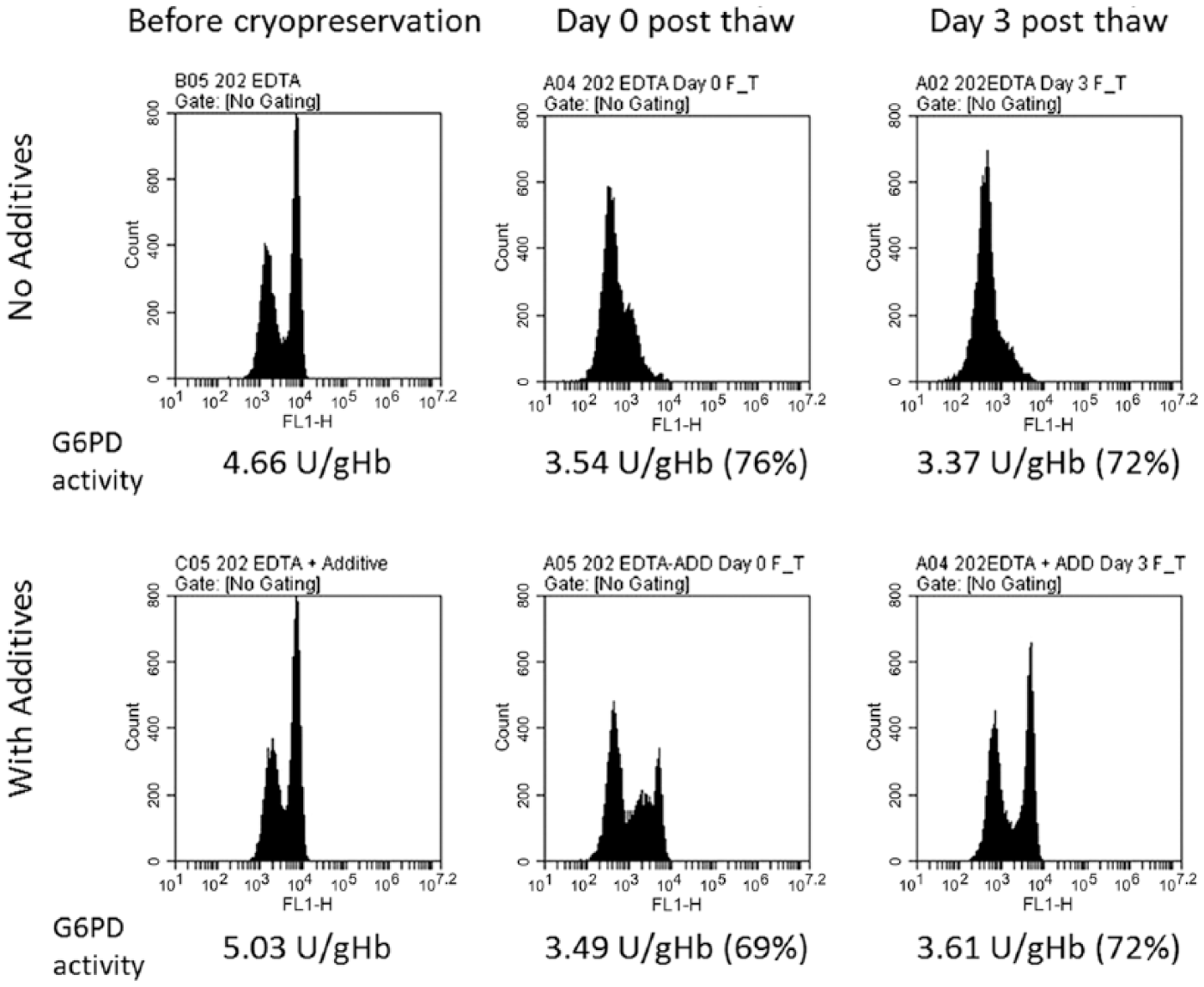
Integrity and stability of a glucose-6-phosphate dehydrogenase (G6PD) heterozygote sample post-cryopreservation. Increasing fluorescence from left to right. Before cryopreservation, the specimen gives a bright peak for cells with normal levels of G6PD and a dim peak for G6PD-deficient cells. The additives have no impact on the quantitative G6PD test performed on lysed blood cells.
Intracellular G6PD Activity as Observed by Flow Cytometry for Specimens Thawed After Cryopreservation With and Without Additive.
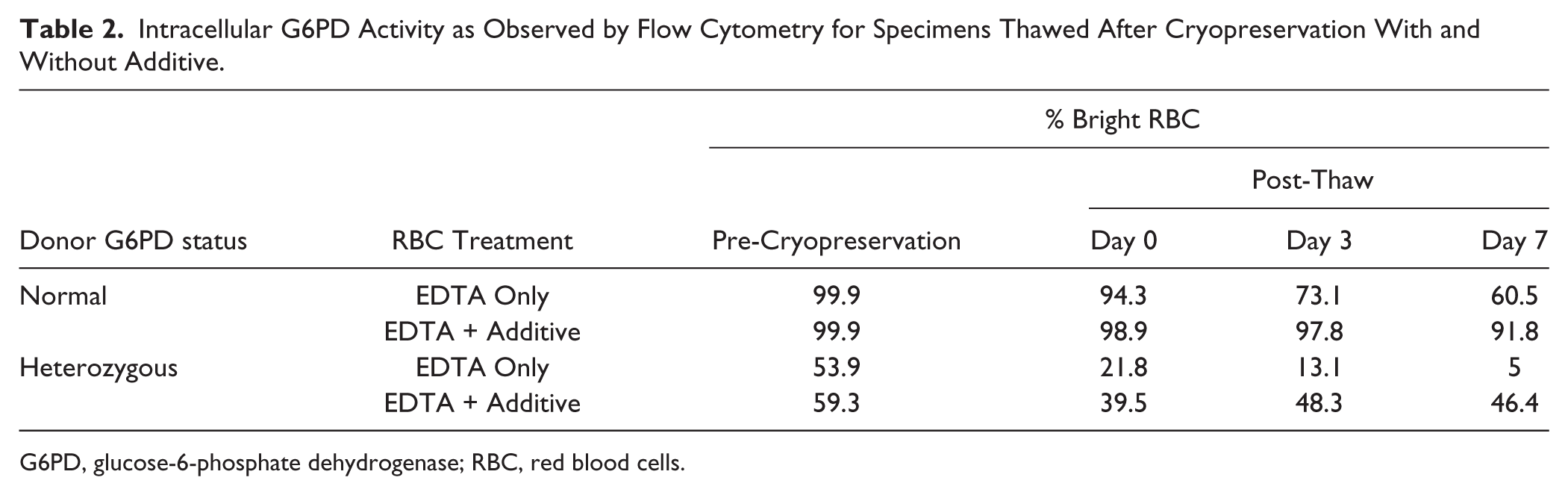
G6PD, glucose-6-phosphate dehydrogenase; RBC, red blood cells.
In an experiment on G6PD normal samples, where the additive was included only 72 hr after collection, the observed compromised cytofluorometric profile before treatment was largely reversed within 1 hr after treatment with the additives. There was no rescue in control experiments in the absence of additives. The additives have no impact on the quantitative G6PD test performed on lysed blood cells (Table 1).
The observed differences between the two G6PD assays are intriguing. Following storage in the absence of additives, the intracellular test gives a false increase in the numbers of deficient cells, despite lysates of the same specimens continuing to exhibit the expected levels of G6PD activity. The enzyme activity in RBC lysates is stable for 21 days if samples are stored at 4°C, either with or without additives. Conversely, treatment with additives is necessary in order to accurately determine G6PD activity inside intact RBCs if the assay is performed more than 48 hr after collection of the sample or after cryopreservation (Figs. 1 and 2).
The assay in RBC lysates requires glucose-6-phosphate and NADP+, both of which are added before there is a change in absorbance at 340 nm, which is primarily due to the G6PD-dependent formation of NADPH. The cytofluorometric method is an indirect assay, relying on additional enzymes and metabolites in order to measure G6PD activity (Shah et al. 2012). Treatment of specimens with additives “rescues” samples from giving false results of a deficiency, indicating that the RBCs retain sufficient activity of each enzyme required for a G6PD normal test result. The assay requires that RBCs contain glucose, ATP, and NADP+, but only glucose is supplied, suggesting that a lack of ATP or NADP+ may be involved in generating false deficiency results. The additives tested during storage include glucose and adenine. Glucose acts as a substrate for glycolysis inside RBCs, facilitating phosphorylation of ADP to ATP. Adenine also enters RBCs, and possibly acts as a precursor for ADP, ATP, and NADP+, when they become depleted during storage. From blood transfusion studies, it has been shown that RBCs can maintain their metabolic activity longer when stored at 4°C when an additive is added (Lion et al. 2010).
The results presented here are encouraging for field studies in which samples may not be analyzed immediately. Specimens can be treated with additives and then robust G6PD data can be measured at offsite laboratories. Previously, we demonstrated the stabilization of G6PD activity inside RBCs by cryopreservation (Kahn et al. 2013). In the absence of additives, integrity in the cytofluorometric assay becomes impaired. The additives extend the functionality of an intracellular G6PD assay, and therefore enhance the utility of cryopreserved samples in the context of specimen panels for the development and evaluation of diagnostic tests for G6PD deficiency, in support of radical cure of P. vivax malaria (Von Seidlein et al. 2013).
Footnotes
Declaration of Conflicting Interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was funded by the Bill & Melinda Gates Foundation, grant number OPP1034534; and the UK Department for International Development (DFID), grant number 204139. The findings and conclusions contained within are those of the authors and do not necessarily reflect positions of the Bill & Melinda Gates Foundation or DFID.