
Abstract
Ionizing radiation (IR) leads to fibrosing alveolitis (FA) after a lag period of several weeks to months. In a rat model, FA starts at 8 weeks after IR. Before that, at 5.5 weeks after IR, the transcription factors Sp1 (stimulating protein 1) and AP-1 (activator protein 1) are inactivated. To find genes/proteins that were down-regulated at that time, differentially expressed genes were identified in a subtractive cDNA library and verified by quantitative RT-PCR (reverse transcriptase polymerase chain reaction), western blotting and immunohistochemistry (IH). The mRNA of the molecular chaperone HSP90AB1 (heat shock protein 90 kDa alpha, class B member 1) was down-regulated 5.5 weeks after IR. Later, when FA manifested, HSP90ab1 protein was down-regulated by more than 90% in lung cells with the exception of mast cells. In most mast cells of the normal lung, both HSP90ab1 and HSP70, another major HSP, show a very low level of expression. HSP70 was massively up-regulated in all mast cells three months after irradiation whereas HSP90AB1 was up-regulated only in a portion of mast cells. The strong changes in the expression of central molecular chaperones may contribute to the well-known disturbance of cellular functions in radiation-damaged lung tissue.
Introduction
Fibrosing alveolitis (FA) is a severe adverse effect after treatment of the thorax with ionizing irradiation (IR). Its final stage is called pulmonary fibrosis (PF). The disease has an inflammatory component but also involves disturbed regulation of cell survival, proliferation and differentiation. These changes are mediated by a number of pro-fibrogenic cytokines and growth factors such as members of the FGF (fibroblast growth factor) family, TNFα (tumor necrosis factor alpha), TGFβ (transforming growth factor beta), PDGF (platelet-derived growth factor) and many more (Crouch 1990; Kovacs 1991; Kovacs & DiPietro 1994; Stone et al. 2004; Fleckenstein et al. 2007).
FA usually occurs after a latency period of several weeks or months. During this latency period, no general disturbance to the lung architecture can be detected with light microscopy (Kasper et al. 1994); however, changes in gene regulation have been detected in a rat model. Whereas the transcription factor NF-κB (nuclear factor kappaB) is continuously up-regulated (Haase et al. 2003), other transcription factors, such as Sp1 (stimulating protein 1) (Haase et al. 2000) and AP-1 (activator protein 1) (Haase et al. 2008), are inactivated during this latency period before the onset of FA. The inactivation of these important regulatory factors might lead to a defective production of protective proteins resulting in a disturbance of cellular functions such as proliferation, differentiation and the production of matrix proteins. One of the protective genes known to be down-regulated is KGF (keratinocyte growth factor) (Haase et al. 2008). In line with its protective function, studies have shown that KGF treatment ameliorates radiation-induced lung injury (Yi et al. 1996; Dorr et al. 2002).
The heat shock proteins (HSPs) comprise a large group of protective genes that are induced after the induction of cellular stressors, such as radiation. These are ubiquitous proteins that function as molecular chaperones, which are proteins that stabilize partially folded or unfolded proteins, or directly facilitate proper protein folding (Lodish et al. 2004; Wegele et al. 2004). Besides some smaller HSPs, two major groups of HSPs have been described: HSP70 and HSP90 (Wegele et al. 2004; Young et al. 2004).
HSPs are highly conserved and highly abundant proteins (Wegele et al. 2004). HSP90 has been described to be essential for cell viability in all eukaryotic cell types (Borkovich et al. 1989; Cutforth & Rubin 1994; Sreedhar et al. 2003). HSP target proteins comprise proteins of virtually all major protein groups, such as receptor proteins, signaling proteins (kinases), polymerases as well as structural proteins (Wegele et al. 2004). For example, HSP90 directly interacts with protein kinases and thereby influences several signal transduction pathways, such as the mitogen-activated protein kinase (MAPK) (Miyata et al. 2001) and the phosphoinositol-3-kinase (PI3K) pathways (Citri et al. 2006). Members of the HSP70 family are involved in the short-term radiation response (Kang et al. 2002). Studies in several cell types suggest that HSP70 proteins mediate radioresistance (Gordon et al. 1997; Calini et al. 2003; Brondani Da Rocha et al. 2004; Matsumoto et al. 2005). This is at least partly mediated by the HSP70-induced stimulation of DNA repair, especially base excision repair (Bases 2006), by inhibiting the Akt pathway (Machida et al. 2005) and by inhibiting apoptosis through its interaction with Bag4 (Bcl-2 associated athanogene 4) in the cellular membrane (Gehrmann et al. 2005). However, HSPs may also increase cell death after radiation by stabilizing p53, as shown in a human T-cell model (Fukumoto & Kiang 2011). Cells with inactivated HSP70 displayed telomere instability and a high frequency of spontaneous chromosomal aberrations, indicating a possible role for HSP70 proteins in the maintenance of genomic stability (Pandita et al. 2004). By stabilizing proteins that arise from mutant genes, HSPs are important players in molecular evolution and in the protection of cancer cells from apoptosis (Rutherford & Lindquist 1998). In this respect, HSP inhibitors have been tested as radiosensitizers. In most studies, HSP inhibitors, such as geldanamycin and radicicol, increased cell killing in the short-term radiation response (Russell et al. 2003; Harashima et al. 2005; Matsumoto et al. 2005).
In pulmonary fibrosis, studies on HSP expression have mostly focused on HSP47, which is involved in the processing and secretion of collagen. The expression of the HSP47 has been shown to be elevated in idiopathic interstitial pneumonias and in bleomycin-induced lung injury (Razzaque et al. 1998b; Ishii et al. 2003; Kakugawa et al. 2005). Therapeutic intervention with the antifibrotic drug pirfenidone attenuated lung injury and reduced the level of HSP47 in many cell types of the lung (Kakugawa et al. 2004). Until now, no studies exist on the role of the major HSPs of the HSP70 and HSP90 groups in radiation-induced, long-term tissue damage of the lung.
The aim of this study was to identify potential protective genes that are down-regulated before the onset of radiation-induced FA. Using a combined quantification of western blot and immunohistochemistry (IH), we demonstrate in an animal model that HSP90ab1 is down-regulated in the radiation-damaged lung in cells other than mast cells. Furthermore, we show that, according to our knowledge, mast cells are the first cell type described that are able to survive with low to non-detectable amounts of HSP90ab1 and/or HSP70.
Materials & Methods
Animal Model
The right lungs of female Fischer rats (aged, 7 weeks; weight, ~120 g) were irradiated with 20 Gy of ionizing radiation, as previously described (Haase et al. 2003). Lungs were removed at the indicated time points and either snap-frozen or fixed in 4% neutral-buffered formaldehyde for exactly 24 hr. Animal housing and experiments were approved by the local authorities and followed the German and international animal welfare regulations. Three animals were used for each time point for histological analysis (control and 3 months after irradiation) and three animals for each time point were used for RT-PCR and western blotting (0, 4, 6, 8, 12 weeks after IR, for RT-PCR additionally 5.5 and 6.5 weeks after IR). Since previous histological analyses have shown that the radiation-induced changes were distributed uniformly, pieces of lung were chosen randomly for the experiments.
Construction of the Subtractive cDNA Library
Messenger-RNA from rat lungs at 4 and 5.5 weeks after irradiation was isolated with polyA-coupled magnetic beads (Dynal, Life Technologies; Carlsbad, CA). cDNA was constructed with the pBluescript II XR cDNA Library Construction Kit (Stratagene, Agilent Technologies, Santa Clara, CA) according to the manufacturer’s instructions. DNA hybridization was done with the formamide-phenol emulsion re-association (F-PERT) technique (Gulick & Dvorak 1990), with two modifications (Casna et al. 1986): the use of 100 ng tester cDNA (driver:tester ratio, 20:1) and the reduction of the total incubation volume to 1 ml. The obtained clones were partially sequenced with an ABI Prism 310 Sequencer (Applied Biosystems, Life Technologies).
Quantitative RT-PCR
Total RNA was isolated with Trizol (Invitrogen, Life Technologies). Primer sequences for HSP90ab1 RT-PCR were GGCCGGGGTACCAAAGTGAT (forward) and TCGCCGGGGAATGAAGAG (reverse). Reverse transcription reactions were performed with the reverse gene-specific primers. GAPDH (glycerolaldehyde-3-phosphate dehydrogenase) RT-PCR was performed as described (Haase et al. 2003) and served as a control reaction. By comparing the band intensities after different numbers of PCR cycles, the linear range of the PCR was determined. Bands of ethidium bromide stained gels were quantified using the ImageJ software (NIH, Bethesda, MD).
Western Blotting
Cytoplasmic proteins were isolated according to a published protocol (Schreiber et al. 1988). Western blotting was performed as described (Haase et al. 2003). Anti-HSP70 (reactive with HSPa1a and HSPa1b) goat polyclonal antibody (K-20, Santa Cruz Biotechnology, Dallas, TX) and anti-HSP90beta goat polyclonal antibody (D-19, Santa Cruz Biotechnology) were both used at a dilution of 1:7500. A monoclonal mouse antibody to beta-Actin (A4700, 1:500; Sigma-Aldrich, St. Louis, MO) served as a loading control.
Histological Staining
H&E and (Masson-Goldner) Trichrome stainings were performed using standard protocols (Böck 1989). Peroxidase-based IH was done as described (Haase et al. 2003). The antibodies mentioned in the western blotting methods section were used at a dilution of 1:10, in case of labeling of bronchiolar epithelium 1:100. For labeling of mast cells in light microscopy, a rabbit polyclonal antibody to IκBγ (sc-201, Santa Cruz Biotechnology) was used at a concentration of 1:100 (Haase et al. 2004). For immunofluorescence (IF) double labeling, the primary anti-HSP antibodies were used at a dilution of 1:100 and detected with the tyramide signal amplification (TSA) kit (Molecular Probes, Life Technologies). For some double labeling experiments (see figure legends), polyclonal goat antibodies to HSP70 (sc-1060) and HSP90ab1 (sc-1057) were used at a dilution of 1:100 (Santa Cruz Biotechnology). The polyclonal goat primary antibodies were used with a biotin bridge anti-goat antibody (DAKO, Glostrup, Denmark) at a dilution of 1:100 (10 ng/ml). The mouse monoclonal antibody to mast cell tryptase (DAKO) was used at a dilution of 1:100. The mouse monoclonal antibody to surfactant protein D ({SFTPD}sc-59695; Santa Cruz Biotechnology) was used at a dilution of 1:70. The rabbit polyclonal antibody to aquaporin 5 (A4979; Sigma Aldrich) was used at a dilution of 1:70. The nuclei were stained with DAPI (4’,6-diamidino-2-phenylindole, Molecular Probes). Fluorescence images were obtained with the laser scanning microscope LSM 510 Meta (Zeiss, Oberkochen, Germany) or an SP5 upright (Leica Microsystems, Wetzlar, Germany). Control staining included replacement of the primary antibody with non-immune serum of the same species and blocking with the peptide against which the antibodies had been raised. The pre-diluted primary antibody was incubated for 1 hr at room temperature with 4 µg of blocking peptide before adding this solution to the section.
Quantification of Proteins
We had the impression that different cell types had a different HSP expression profile after irradiation. Therefore, we sought to quantify HSP expression in different cell types at one time point as well as over time. Although quantification of IH data is a matter of debate, there have been numerous examples of reliable quantification of such experiments (van der Loos et al. 1994; Matsuo et al. 1995; Taylor & Levenson 2006; Walker 2006; Vilmar et al. 2012). The major source of variation is staining intensity, which is influenced by the multiple steps involved with tissue preparation. However, there is no doubt that the different staining of various cell types within one section represents the real expression of proteins provided that the whole section has been treated homogenously. We circumvented the problem of variation between different tissue samples by a combination of methods. The quantification of total HSP90ab1/HSP70 protein content over time was done by western blotting. This was combined with a quantification of the relative protein expression in different cell types by IH and IF, as follows.
Relative amounts of total HSP proteins were determined using western blotting in three independent experiments with the help of the ImageJ/Fiji software (Schindelin et al. 2012). Relative intensities (RI) of the bands in control lungs were set to 1 and adjusted to the values of the loading control (β-actin). In order to correlate the contribution of different cell types (mast cells versus non-mast cells) to the total amount of protein, IF double staining was performed with HSP70 or HSP90AB1 and the mast cell marker mast cell tryptase (MCT) and recorded with a laser scanning microscope (Leica, LSM SP5). The settings of the laser power and gain were adjusted for the HSP recordings such that the background staining was approximately zero. From each of three independent experiments, three randomly chosen fields of 600625 µm2 were analyzed with Fiji software (Schindelin et al. 2012). This area contained more than 1,400 mast cells. For determination of total lung HSP stained sections, the threshold in the MCT stained sections were set to 15 which highlighted all lung tissue. With these settings, all particles (particle size 0–infinity) were analyzed and transferred to the region of interest (ROI) manager. This mask was transferred as an overlay to the HSP image and the integrated density of the HSP protein in lung tissue was determined. In order to determine the portion of mast cell HSP, the threshold in the MCT staining was set to 150, which highlighted only mast cells. Particles with sizes of 0.001–infinity were analyzed and treated as described above.
Quantification of mast cell numbers was done in separate experiments using histological sections (thickness, ~1 µm) stained with trypan blue using a Zeiss Axioskop microscope equipped with an ocular micrometer. Ten randomly chosen fields of 0.6 mm2 were counted in three different sections of normal lungs and lungs obtained three months after irradiation.
Results
Histopathological Changes in the Irradiated Rat Lung
The normal lungs of Fischer rats had alveoli with thin walls (Fig. 1A, 1B). Assuming the diameter of a red blood cell to be ~7.5 µm, the alveolar walls were usually not thicker than about 10–12 µm. Three months after irradiation with 20 Gy of IR, FA was detectable (Fig. 1C–1F). Tissue architecture had changed considerably. Large amounts of extracellular matrix were accumulated as shown by the turquoise color in the Trichrome stain (Fig. 1D and 1F). Aggregates of foamy macrophages were found in the remaining alveolar spaces (arrows in Fig. 1D–F). In addition, a large number of mast cells infiltrated the lung tissue (this is demonstrated in later figures). These changes started to occur at about 8 weeks after irradiation and were distributed focally in the lung, covering about 5 to 10% of the total lung tissue except for the mast cell infiltration which started at about 6 weeks after irradiation and affected the entire irradiated lung tissue. During the time period of this study (up to 3 months after irradiation), no areas with cell-free scar tissue as a feature of full-blown PF were detectable.
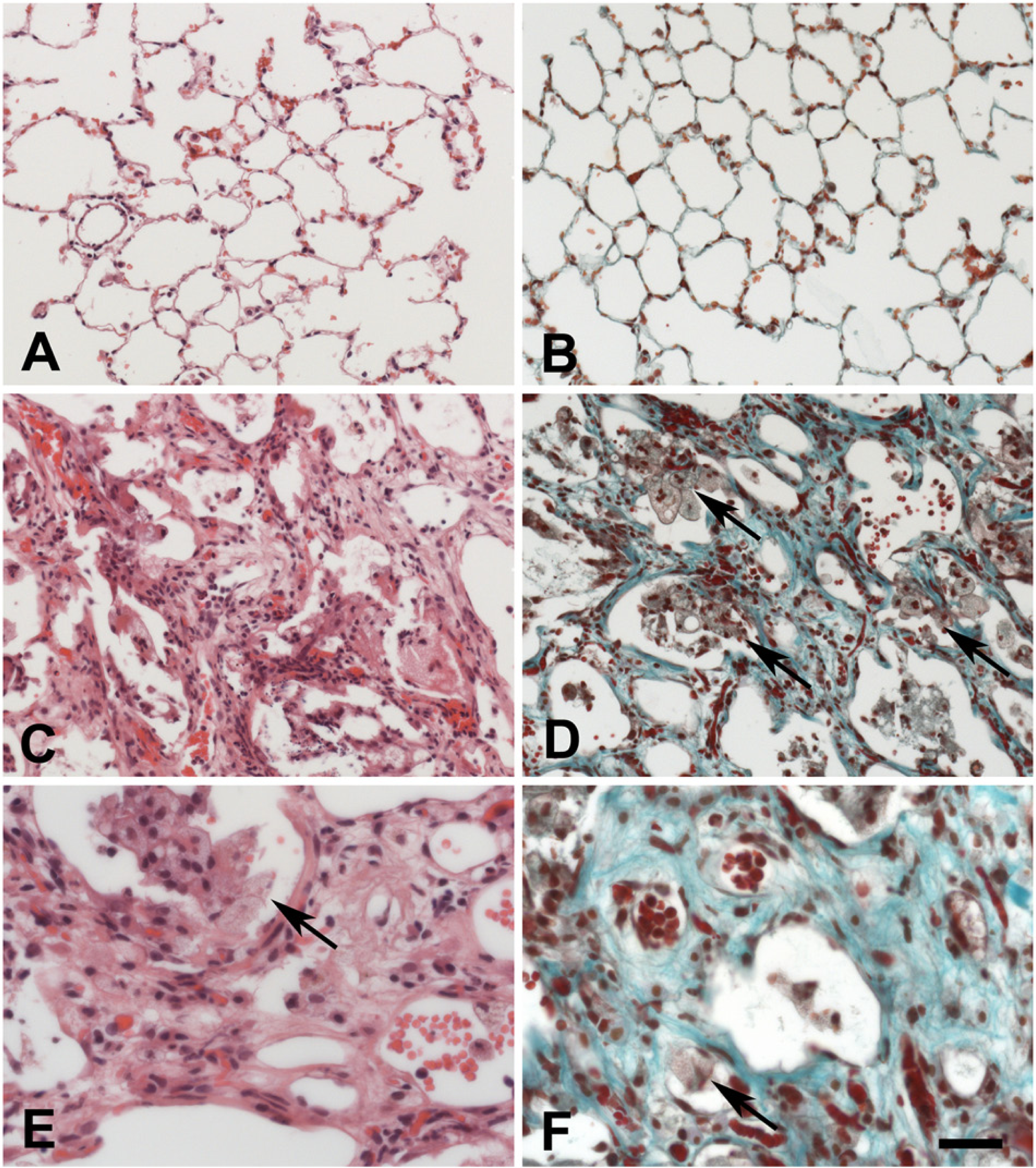
Histology of normal rat lung (A, B) and histopathological changes in the rat lung at 3 months after irradiation (C–F). Sections were stained with H&E (A, C, E) and Trichrome (B, D, F) staining. Pathological changes include the accumulation of foamy macrophages (arrows) in the alveolar spaces as well as an accumulation of extracellular matrix components (turquoise in D and E). The bar corresponds to 50 µm in A, B and 25 µm in C–F.
Expression of HSP90ab1 mRNA in Irradiated Lungs
After irradiation of the right hemithorax of rats with 20 Gy, the major transcriptional changes occur between 4 and 5.5 weeks after IR. Therefore, a subtractive cDNA library was constructed that identified mRNAs that were down-regulated at 5.5 weeks compared with that at 4 weeks after IR of the lung. In this library, the cDNA of HSP90ab1 was identified by DNA sequencing. In order to verify the down-regulation of HSP90ab1 mRNA, quantitative RT-PCR was performed. These experiments demonstrated that the mRNA of HSP90ab1 was down-regulated at 5.5 weeks after IR and remained repressed below that of the control level (Fig. 2A, 2B), with the exception of a peak at 6.5 weeks after IR. One representative figure of three independent experiments is shown. The next step was to examine whether IR also affected protein expression levels.
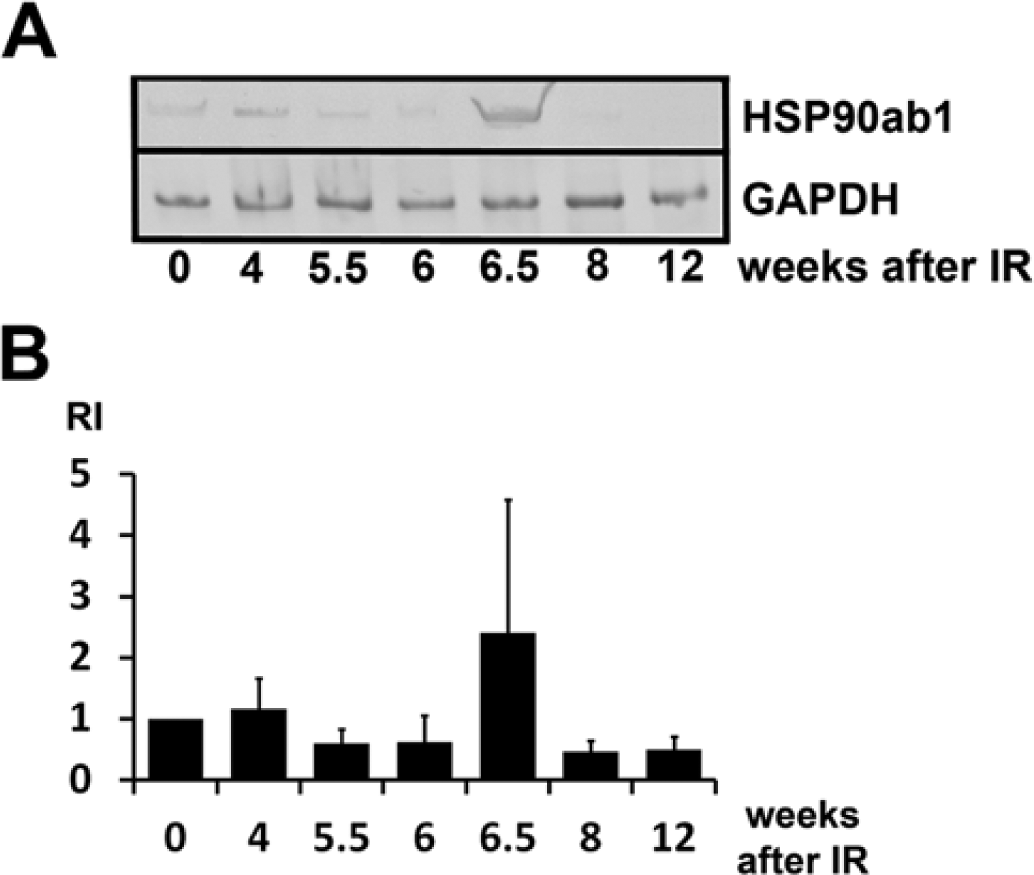
Quantification of HSP90ab1 (HSP90beta) mRNA by RT-PCR in total RNA extracts from irradiated rat lungs represented as a gel photo from one out of three representative experiments (A) and as a diagram representing the values of three independent experiments (B). Relative intensities (RI) of western blotting bands were corrected against the loading control (GAPDH). Error bars represent the SD. HSP90ab1 mRNA was decreased to about 60% of the control level at 5.5 weeks after irradiation (IR).
Protein Expression of HSP70 and HSP90ab1 in Irradiated Rat Lungs
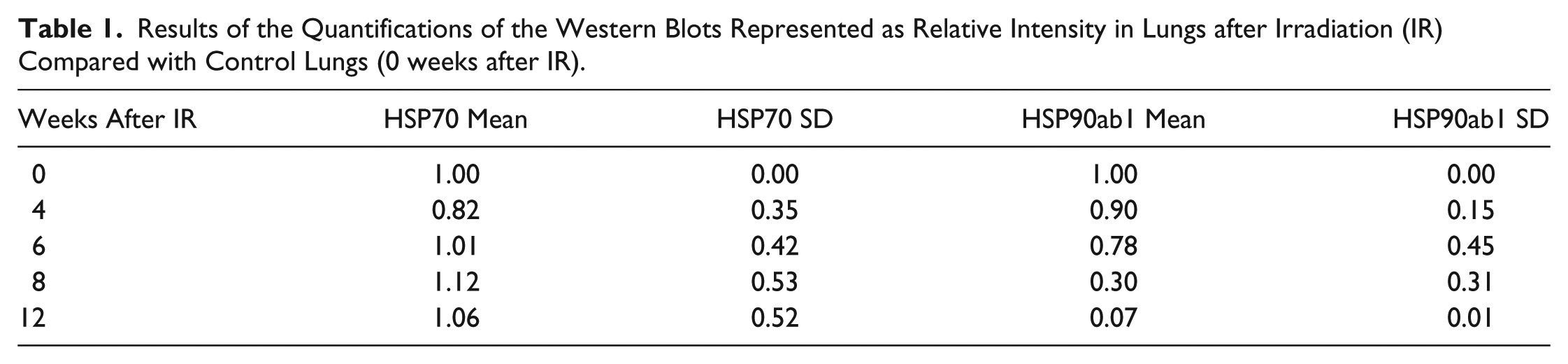
To determine quantitative HSP90ab1 protein expression, cytoplasmic protein extracts from lungs were prepared at several time points after IR and subjected to western blotting. For comparison, another major HSP, HSP 70, was included in the study. The HSP70 protein level remained nearly constant after irradiation (Fig. 3A, 3B; Table 1). At the same time, HSP90ab1 protein was down-regulated. Quantification of total protein using relative intensity (RI) showed that total HSP90ab1 was reduced to 0.07 (7%) in the rat lung at 12 weeks after irradiation as compared with protein levels at 0 weeks after IR (taken as 1.0) (Fig. 3A, 3C; Table 1). One representative figure of three (HSP90AB1) and four (HSP70) independent experiments is shown in Figure 3. In order determine the cell types that were affected, subsequent IH experiments were performed.
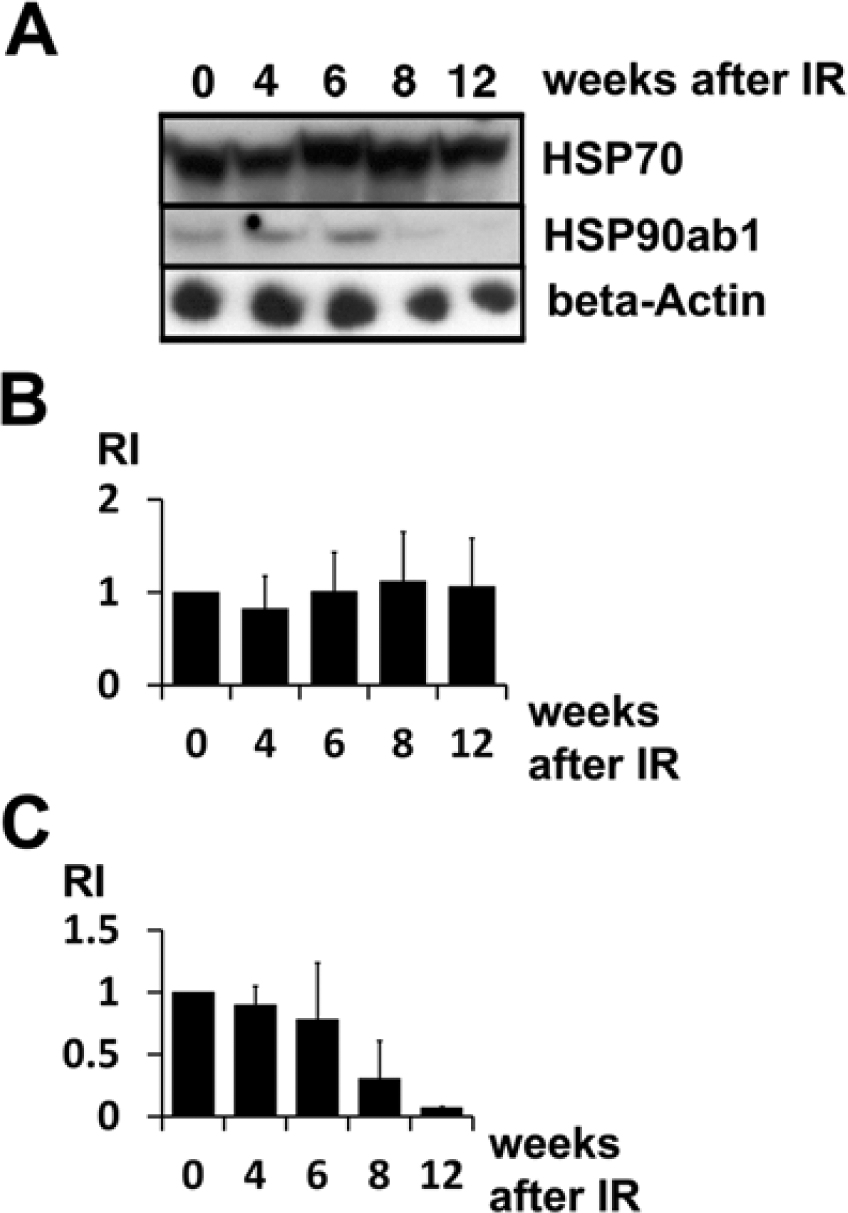
Protein levels of HSP70 and HSP90ab1 in irradiated rat lungs determined by western blotting using cytoplasmic protein extracts of total lungs. (A) Blots for protein samples harvested over time for HSP70 and HSP90ab1 as compared with loading control (β-actin). (B-C) Relative intensities (RI) of bands after β-actin correction for HSP70 (B) and HSP90AB1 (C) proteins as calculated from western blots. The values of control lungs (0 weeks after IR) were arbitrarily set to 1.0. HSP70 is strongly detectable in control lungs at all time points after IR, with a slight increase of the absolute value at 8 and 12 weeks after IR. HSP90ab1 protein was reduced at 8 weeks after IR, and finally at 12 weeks to less than 10% of the control level. Three independent experiments were performed. Values at 12 weeks after IR—after induction of fibrosing alveolitis—ranged from 0.06 to 0.08, with a mean value of 0.07 as compared with the control level. Error bars represent SD.
Results of the Quantifications of the Western Blots Represented as Relative Intensity in Lungs after Irradiation (IR) Compared with Control Lungs (0 weeks after IR).
Cellular Distribution of HSP70 and HSP90ab1 in Normal and Irradiated Lungs
In normal rat lungs, both HSP70 and HSP90ab1 proteins were ubiquitously expressed (Fig. 4A–4D), with strong expression in type II pneumocytes (red arrows), type I pneumocytes (black arrows), alveolar macrophages (most cells in the alveolar lumen), bronchial epithelial cells (arrowhead with +) and in endothelial cells (arrowhead with asterisk). A moderate to low expression level was observed in smooth muscle cells of bronchiolar walls (Fig. 4B) and blood vessels (Fig. 4B).
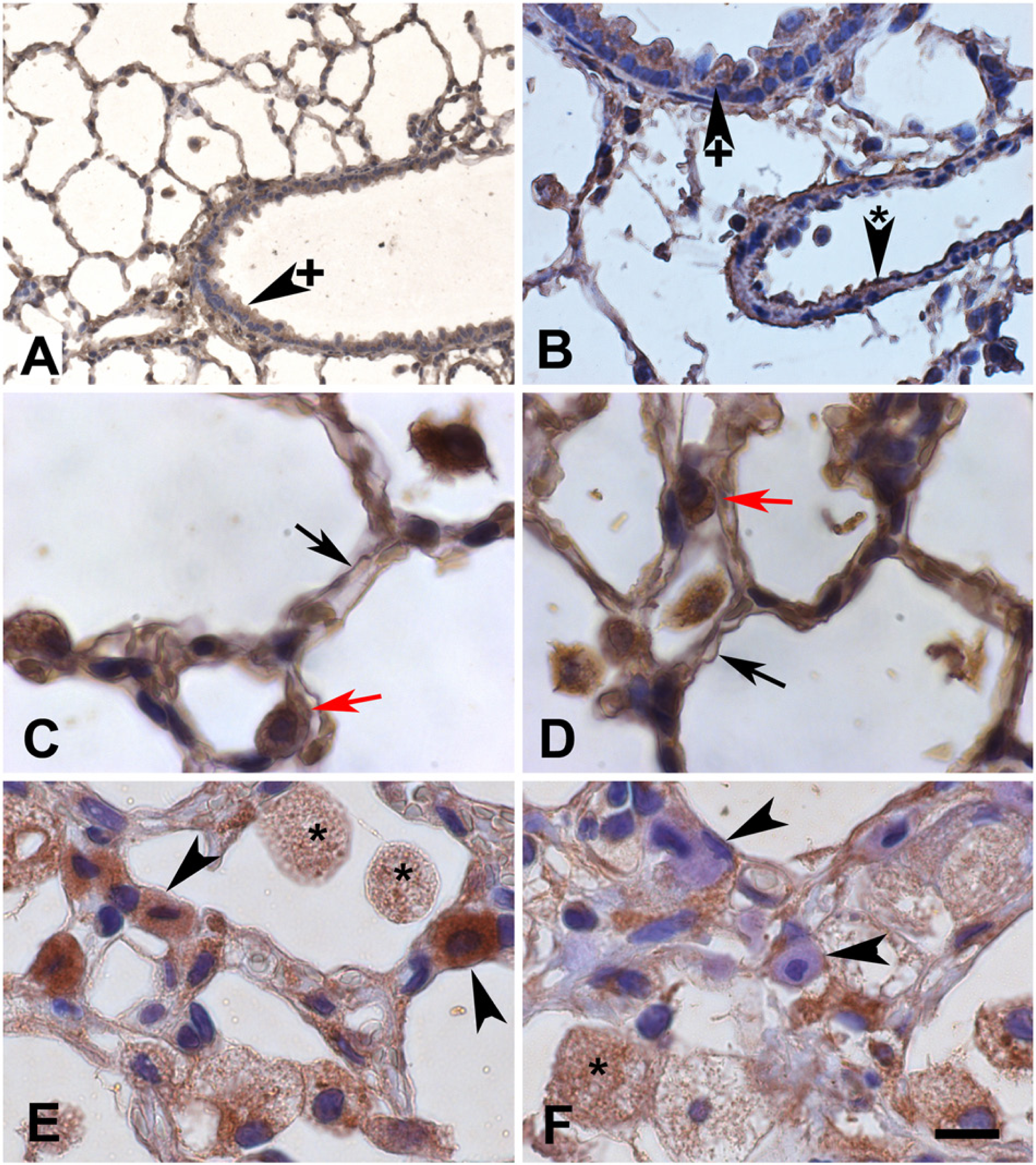
Immunohistochemistry (IH) demonstrated positive HSP70 (A, C, E) and HSP90ab1 (B, D, F) protein expression (brown) in normal lungs (A–D) and lungs with fibrosing alveolitis (FA) at three months after irradiation (E, F). Most cell types were positive for both proteins including bronchial epithelial cells (arrowheads with +), endothelial cells (arrowheads with asterisk), type I (black arrows) and type II (red arrows) pneumocytes, as well as foamy alveolar macrophages (asterisks). Mast cells in irradiated lungs (arrowheads) were positive for HSP70 (E) but negative for HSP90ab1 (F). The bar corresponds to 50 µm in A, 16 µm in B, 10 µm in C–F.
In irradiated rat lungs three months after irradiation (Fig. 4E, 4F), the same distribution pattern was observed for both proteins. However, the HSP90ab1 staining intensity in cells of the alveolar walls, such as type I pneumocytes, endothelial cells (not indicated in Fig. 4) and in alveolar macrophages seemed to be reduced. However, alveolar macrophages in the irradiated lungs were enlarged, formed aggregates (Fig. 1 D–1F; arrows) and had a foamy cytoplasm with a granular distribution of both HSP70 and HSP90ab1 (Fig. 4E, 4F; asterisks). In contrast to the normal lung, the irradiated lung contained numerous mast cells that were strongly positive for HSP70 protein (Fig. 4E, arrowheads). Surprisingly, many mast cells contained no HSP90ab1, as judged by IH (Fig. 4F, arrowheads). In order to verify that these cells really represented mast cells, we performed IF double labelling with the mast cell-specific enzyme, mast cell tryptase (MCT, Fig. 5). In the normal rat lung, only single mast cells could be detected (see also next paragraph). These mast cells contained no detectable or low amounts of HSP70. An example of a subpleural, MCT-positive mast cell of the normal lung (Fig. 5A–5C) contained very low amounts of HSP70 (Fig. 5B, 5C). All MCT-positive mast cells detected in the normal rat lung contained very low or no HSP90ab1 (Fig. 5D–5F). In lungs with FA, mast cells (Fig. 5G, 5I, 5K, 5M) were strongly positive for HSP70 (Figure 5H, 5I) but were often negative for HSP90ab1 (Fig. 5L, 5M). IF double labelling with the type I pneumocyte marker aquaporin 5 (aqua 5) also confirmed the positivity of type I cells for HSP90ab1 (Fig. 6A–6C), as did double labelling with the type II pneumocyte marker surfactant protein D (SFTPD) (Fig. 6D–6I). These double labelling experiments performed with the HSP70 antibody gave the same results (data not shown).
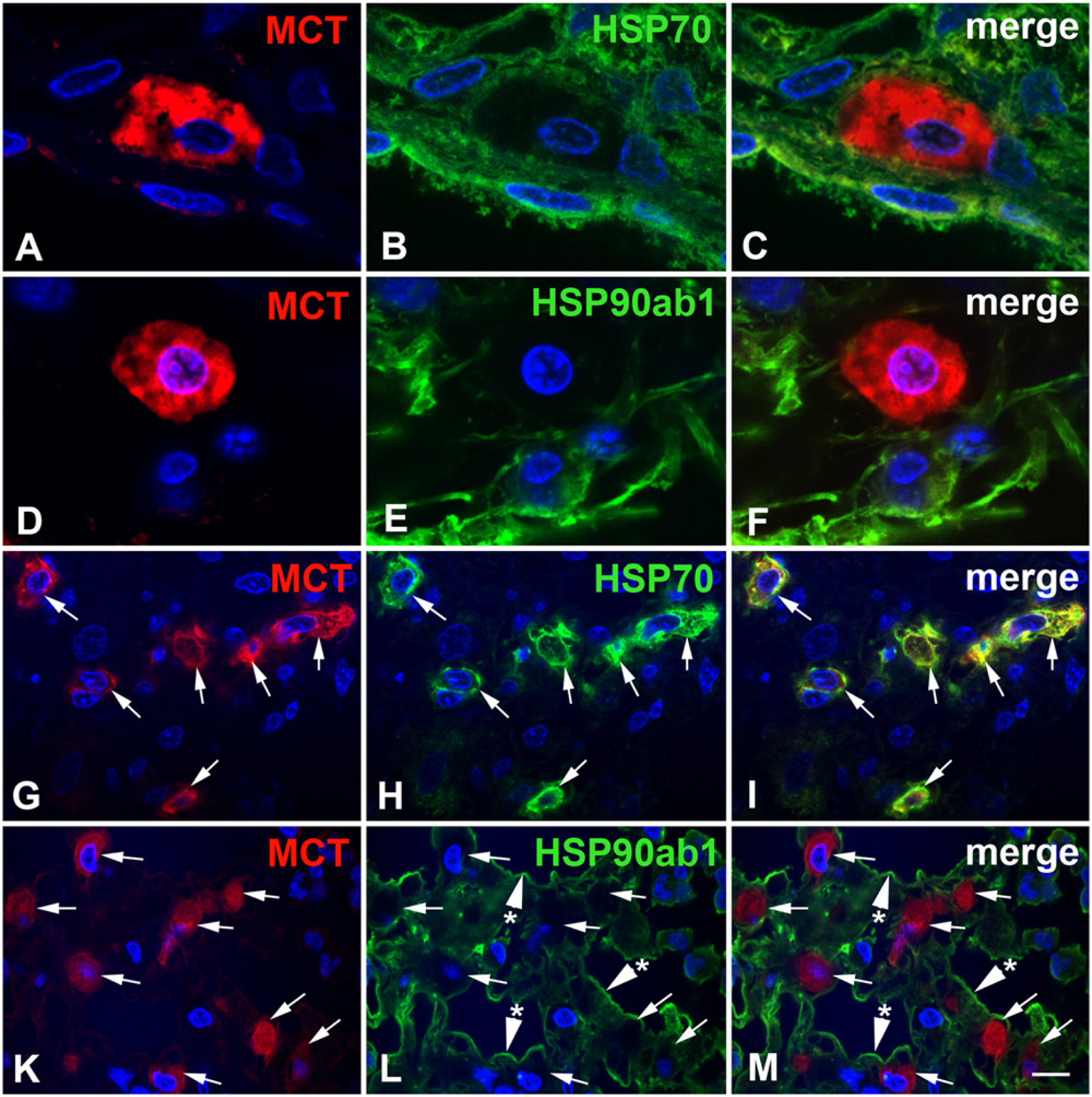
Immunofluorescence (IF) double staining for HSP70 or HSP90ab1 expression (green) and mast cell tryptase (MCT, red) in normal rat lungs (A–F) and rat lungs with fibrosing alveolitis (FA) at three months after irradiation (G–M). (A–C) In the normal rat lung, a subpleural mast cell was specifically labeled with mast cell tryptase (MCT; red), but contained very low amounts of HSP70 (green). (D–F) MCT-positive mast cell of the normal lung was negative for HSP90ab1 (green), except for some very weak, speckled cytoplasmic staining. (G–M) In rat lungs with fibrosing alveolitis, MCT-positive mast cells (arrows) contained high amounts of HSP70 (H) but were often negative for HSP90ab1 (L; arrows). Type I pneumocytes (L, M; arrowheads with asterisk), however, were positive for HSP90ab1. Nuclei were counterstained with DAPI. The bar corresponds to 4.5 µm in A–F and 10 µm in G–M.
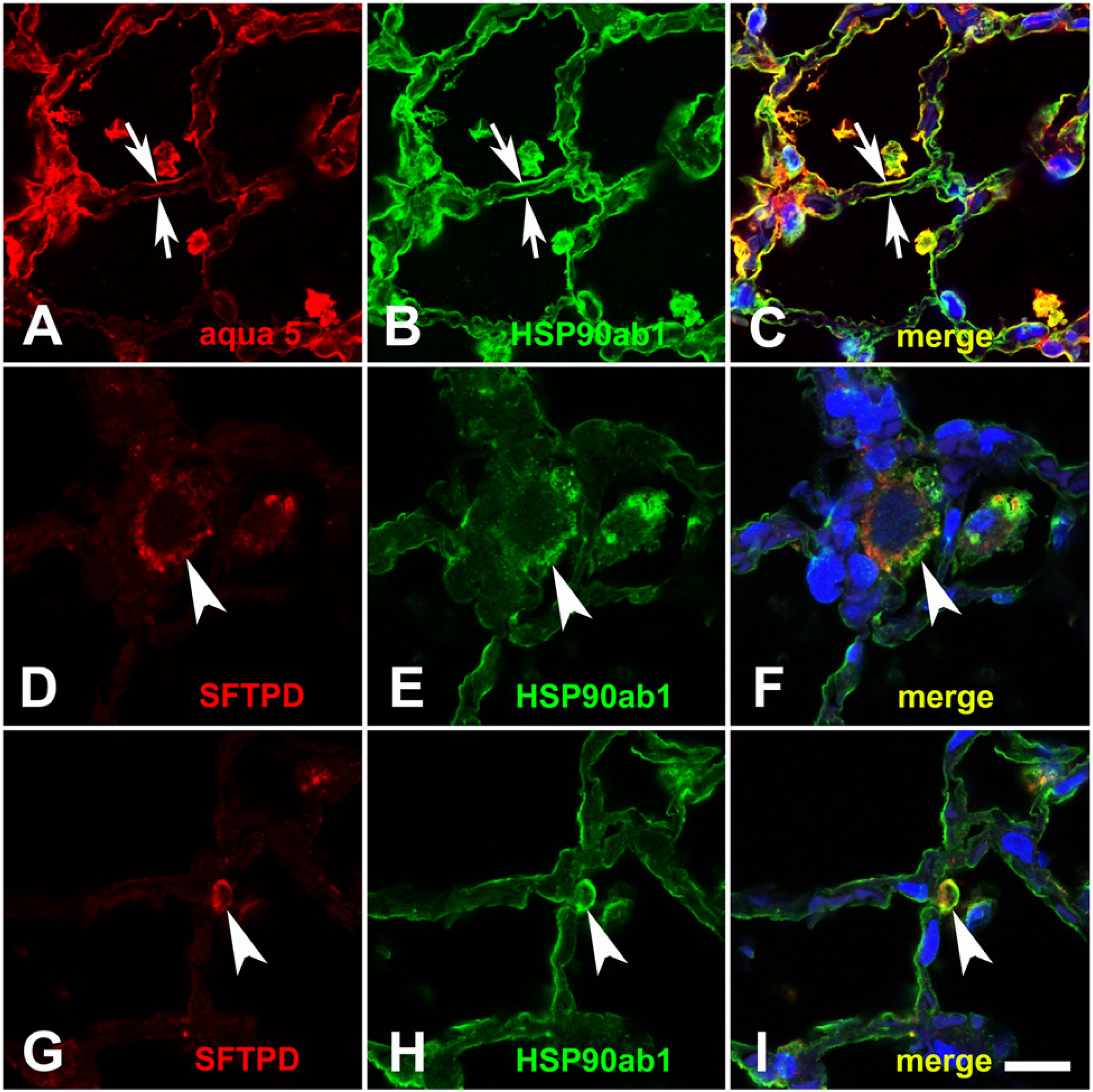
IF double staining of normal rat lung for type I and type II pneumocytes. Aquaporin 5 (aqua 5, red in A, C) and HSP90ab1 (green in B and C) double staining demonstrated that type I pneumocytes were positive for HSP90ab1 (arrows). Double staining of type II pneumocyte marker surfactant protein D (SFTPD; red in D, F, G, I) and HSP90ab1 (green in E, F, H, I) demonstrated that type II pneumocytes were also positive for HSP90ab1 (arrowheads). Nuclei were counterstained with DAPI. The bar corresponds to 14 µm.
Quantification of Mast Cell Protein Expression
Mast cell quantification of the staining for the protein markers is given in Table 2. Normal lungs contained only very few mast cells, ~8 cells/cm2, in histological sections. However, from about 6 weeks after irradiation, a massive increase in mast cell number was observed. Irradiated lungs at 3 months after irradiation contained about 11,400 mast cells/cm2, which corresponds to an approximate 1400-fold increase as compared with mast cell counts in control lungs. In the normal lungs, mast cells were almost exclusively found in the peribronchiolar (peribronchial) or subpleural regions. However, in the irradiated lungs, mast cells were distributed across all regions of the lung including the alveolar walls. At three months after irradiation, mast cells represented more than one third (37%) of the cells in the rat lung and covered 8.5% of the lung tissue.
Quantification of Western Blotting Combined with Histochemistry in Lungs at 12 Weeks After Irradiation (IR) Versus Control Lungs (0 Weeks).
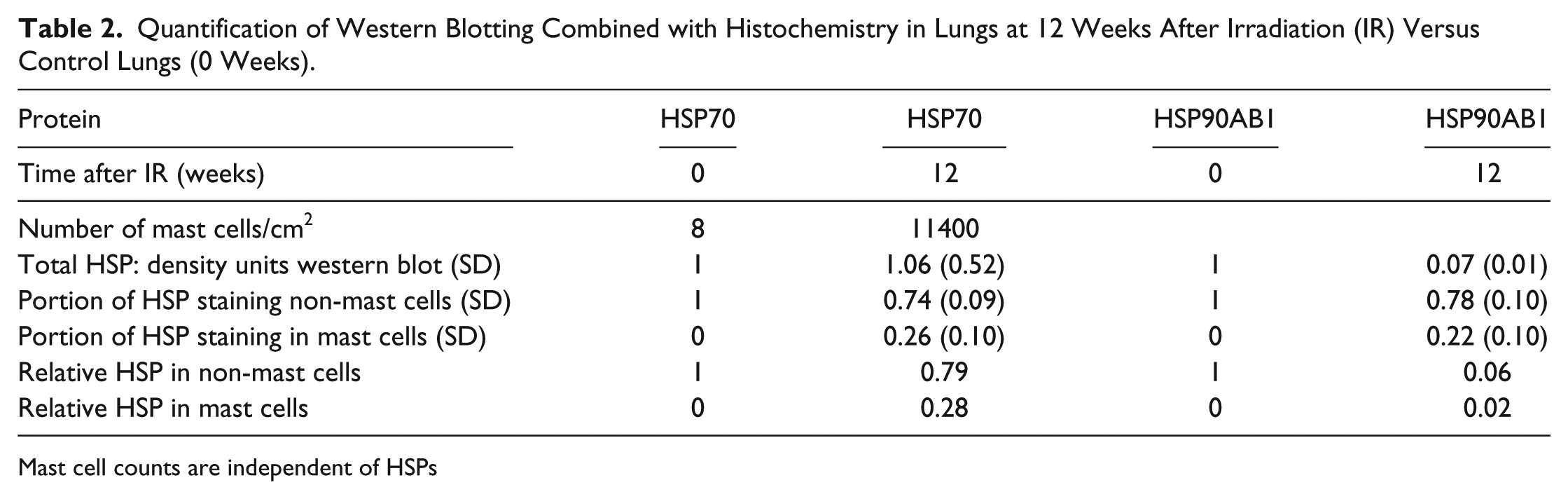
Mast cell counts are independent of HSPs
In order to determine the relative contribution of mast cells to the total amount of HSPs, HSP protein content in the total lung was related to the HSP content in mast cells (Table 2). The proportion of mast cell HSP70 was 0.26 (26%) and that of mast cell HSP90ab1, 0.22 (22%). the amount of HSP70 in non-mast cells was reduced to 0.79 (79%) (Table 2). This reveals that the almost constant HSP70 levels in the lung at three months after irradiation was caused by the emergence of mast cells. Instead, the reduction of HSP90ab1 to 0.07 determined in western blotting was slightly lowered to 0.06 by the mast cells that were positive for HSP90ab1. Taken together, HSP70 protein was slightly reduced whereas HSP90ab1 protein expression was more than 10-fold reduced in lung cells other than mast cells in radiation-induced FA (Fig. 7, Table 2).
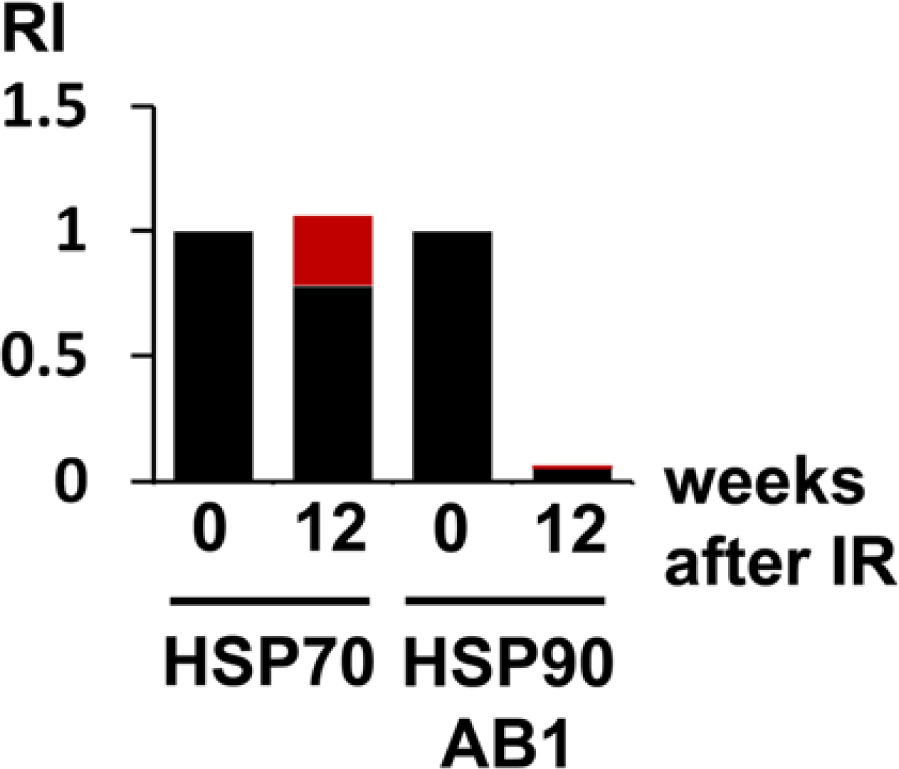
Relative protein amounts of HSP70 and HSP90ab1 in lung cells other than mast cells (black) versus mast cells (red) of control lungs (0) and lungs at 12 weeks (12) after irradiation (IR). Relative intensity (RI) values were obtained after a combined calculation of total protein amounts (western blotting), and relative staining intensities of the cell types. In lung cells other than mast cells, HSP70 was only slightly reduced whereas HSP90ab1 was reduced to about 6% (0.06) of the control level. Control levels were set arbitrarily to 1.
Discussion
In this model of IR-induced FA, previous studies have noted major changes in transcription factor activities between 4 and 5.5 weeks after irradiation. In particular, the activities of the transcription factors Sp1 and AP-1 are down-regulated during this time (Haase et al. 2000; Haase et al. 2008). This implies that the transcription of Sp1- and AP-1-dependent genes would also be down-regulated at that time. Thus, we searched for such genes in a subtractive cDNA library and found HSP90ab1. Indeed, the mRNA of HSP90ab1 is down-regulated between 4 and 5.5 weeks after IR. We also tested the expression changes of a member of another big family of HPSs, HSP70, as a control. Both HSP genes are possible target genes for Sp1 and AP-1 since the HSP70 promoter in various species, including mouse and man, contains binding sites for these two transcription factors (Morgan 1989; Williams et al. 1989). The murine HSP90ab1 promoter also contains potential binding sites for Sp1 and AP-1 (Dale et al. 1996). HSP90ab1 mRNA remains low during the time of observation, which corresponds to the activities of both Sp1 and AP-1 in this model (Haase et al. 2000; Haase et al. 2008). One exception to the down-regulation of HSP90ab1 mRNA occurred at 6.5 weeks after irradiation. We have no explanation for this short-term increase, which occurred in two of three independent experiments and was not accompanied by changes in AP-1 activity (Haase et al. 2008). The down-regulation of HSP90ab1 protein occurs at time points between 6 and 8 weeks after IR, most likely after the late mRNA peak at 6.5 weeks. The down-regulation of HSP90ab1 was unexpected since radiation damage induces cellular stress, including stress caused by radiation-induced mutations. HSPs function to stabilize mutated proteins (Rutherford & Lindquist 1998; Yahara 1999); therefore, one would expect an adaptive increase of all HSPs after irradiation. But despite the increased cellular stress, we observed the opposite for HSP90ab1. The down-regulation of HSP90ab1 suggests that, in radiation-damaged tissue, the stability of altered proteins might be even lower compared with that of control tissue. It is suspected that the down-regulation of HSP90ab1 is likely to aggravate the cellular injuries in radiation-damaged tissue.
In contrast to their down-regulation in the long-term radiation response, HSPs have been described to be activated in response to short-term IR and UV radiation (Trautinger et al. 1999; Park et al. 2000; Kang et al. 2002). The transcription factor Ap-1, like many acute-phase response genes, is also up-regulated shortly after irradiation (Hallahan et al. 1995; Prasad et al. 1995; Hallahan et al. 1996; Usenius et al. 1996; Martin et al. 1997; Shaulian & Karin 1999). This is another indication that the level of HSP expression is likely to be regulated by AP-1. These data also demonstrate that the short- and long-term responses to ionizing radiation are fundamentally different at the molecular level concerning their regulation by transcription factors and HSPs. In line with its protective role during cellular stress, it has been demonstrated that induction of HSP70 protects against bleomycin-induced pulmonary fibrosis (Fujibayashi et al. 2009). However, there is no uniform mechanism for all types of fibrosis. In a model of ureteral obstruction-mediated renal fibrosis, the HSP90 inhibitor 17-allylamino-17-demethoxygeldanamycin (17AAG) inhibited renal fibrosis that occurred 2 weeks after the injury (Noh et al. 2012). Both the time course of the changes (2 weeks versus 8 weeks) and the likely different role of HSP90 suggest the involvement of other regulatory molecules in the pathogenesis of fibrosis induced by mechanical damage versus radiation-mediated damage.
In our library, we did not detect Hsp47, which has been described elsewhere to be overexpressed in various fibrosing disorders such as bleomycin-induced pulmonary fibrosis (Razzaque et al. 1998b; Ishii et al. 2003; Kakugawaet al. 2004) and idiopathic fibrotic lung diseases (Razzaque et al. 1998a; Iwashita et al. 2000; Kakugawa et al. 2005). One possible reason is the fact that we did not construct a library that compared normal lungs to lungs with full-blown fibrosis. Instead, our library was aimed at detecting proteins that were down-regulated at 5.5 weeks after irradiation as compared with that at 4 weeks after irradiation, since major regulatory changes occur around this time period (Haase et al. 2008).
Using IH and IF, we demonstrated that, in normal rat lungs, some mast cells contain low amounts HSP70. In addition, the level of HSP90ab1 is very low to not detectable in mast cells of both normal lungs and in a portion of mast cells in radiation damaged lungs. This was unexpected as not merely the presence but the maintenance of high levels of molecular chaperones has been described to be necessary for cell survival in both prokaryotic and eukaryotic cells (Borkovich et al. 1989; Sreedhar et al. 2003). Mast cells especially were expected to contain a high amount of HSPs because they are rich in proteases and signaling proteins that are possibly required to be stabilized by chaperones (Caughey 1994; Haase et al. 2004). In the lung specimens examined, no other cell type was found to be negative for HSP70 and HSP90ab1. As a compensation for the low amount of expression, part of the function of HSP90ab1 might be taken over by other isoforms such as HSP90aa1 or, in the irradiated lung, by members of the HSP70 family. Yet, HSP70 proteins cannot completely fulfill the functions of HSP90 proteins (Wegele et al. 2004; Young et al. 2004). In any case, the lack of HSP90ab1 and the partial lack of HSP70 is a phenomenon that has not been described for any other cell type. Further studies should clarify whether the lack of specific HSPs is restricted to mast cells in this model and focus on the consequences of HSP90ab1 (and, in part, HSP70) deficiency for the function of specific proteins.
Two subpopulations of mast cells have been described: mucosal mast cells (MMCs) and connective tissue mast cells (CTMCs) (Wilkes et al. 1992). Since we found mast cells in the normal lung that were negative as well as moderately positive for HSP70, these mast cells may represent different subtypes of mast cells or different functional states of one subtype. In our model, most of the mast cells in the normal lungs were observed in the deep peribronchiolar region or in the subpleural tissue. Therefore, the CTMCs seem to be the dominant type. This finding is in favor of the view that the different content of HSP70 represents different functional states. In contrast to normal lungs, all mast cells (>99%, as judged by the co-localization of MCT-positive and HSP70-positive cells) from radiation-damaged lungs were strongly positive for HSP70 protein. This may be the result of three different processes or a combination thereof: (I) HSP70 is up-regulated in all mast cells of the lung following radiation damage; (II) there is a selective expansion of HSP70-positive pulmonary mast cells and an additional up-regulation of HSP70 in these cells; or (III) there is a selective accumulation of HSP70-positive bone marrow-derived mast cells in the radiation-damaged lung. Further studies are required to clarify this question.
In general, mast cells are thought to play an important role in pulmonary fibrosis (Broide et al. 1990; Schildge et al. 2003). In addition to their release of pro-inflammatory cytokines, the mitogenic activity of MCT may be one mediator (Ruoss et al. 1991). Therefore, knowledge of the specific regulatory mechanisms in mast cells should help to gain a better understanding of the pathogenesis of pulmonary fibrosis. In this study, we described some properties of mast cells that discriminate them from surrounding cells and these findings may help to design targeted therapies for pulmonary fibrosis.
Considering the necessity to maintain high levels of molecular chaperones HSP70 and HSP90ab1 for cellular survival, their decrease in radiation-induced lung damage suggests a role for these proteins in the development of FA. Mast cells are the only cell type described that can survive with low levels of HSP70 and HSP90ab1, which raises the question as to which regulatory mechanisms may have evolved in these cells to compensate for the low level of chaperones.
Footnotes
Acknowledgements
The authors thank Anke Klawitter for excellent technical assistance.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by the Deutsche Forschungsgemeinschaft (DFG, MU 1299/1-1) and the BMBF 01ZZ9604, as well as a grant from the Faculty of Medicine of the Dresden University of Technology to M.G.H.