
Abstract
The differential distribution of microtubules in osteoclasts in culture was examined by using antibodies against acetylated, tyrosinated, or detyrosinated tubulins. Tyrosinated tubulin was found throughout the cytoplasmic microtubules in all cells examined. An expanding protrusion that contained tyrosinated tubulin but none of the detyrosinated or acetylated form was seen in the immature osteoclasts. Detyrosinated or acetylated tubulin was detectable in the peripheral cytoplasm of the mature osteoclasts displaying the loss of the expanding protrusion. Although most of the microtubules were derived from the centrosome, noncentrosomal microtubules were distributed in the expanding protrusion, which was predominantly positive for tyrosinated tubulin. By tracing single microtubules, the authors found that their growing ends were always rich in tyrosinated tubulin subunits. End binding protein 1 bound preferentially to the microtubule ends. Both acetylated and tyrosinated microtubules were shown to be closely associated with podosomes. Microtubules appeared to grow over or into the podosomes; in addition, the growing ends of single microtubules could be observed to target the podosomes. Moreover, a microtubule-associated histone deacetylase 6 was localized in the podosomes of the osteoclast. On the basis of these results, the authors conclude that posttranslational modifications of microtubules may correlate with characteristic changes in podosome dynamics in osteoclasts.
Osteoclasts are highly polarized cells from morphological or functional points of view. Polarization of the osteoclasts is associated with the cytoskeleton dynamics and cell substratum adhesion. Coordination of cytoskeleton dynamics and cell adhesion mechanisms is critical for efficient cellular activities. The establishment and maintenance of these polarities depend in part on the function of microtubules (Turksen et al. 1988; Mulari et al. 2003; Okumura et al. 2006). Microtubule involvement in the assembly of podosomal adhesions is well documented (Babb et al. 1997; Destaing et al. 2005; Jurdic et al. 2006). Microtubules are requisite for the initial formation of podosomes, but the lack of podosomes themselves does not affect the organization of the microtubule network (Linder et al. 2000). Recent advances have revealed that the posttranslational modification of tubulin subunits, such as acetylation/deacetylation and tyrosination/detyrosination, can regulate microtubule function and organization (Westermann and Weber 2003).
Accumulated data indicate that acetylation or detyrosination of the tubulin subunit results in stable or long-lived microtubules with a low turnover. In contrast, tubulin tyrosination occurs in dynamic microtubules having a high turnover (Witte et al. 2008). These posttranslational modifications have been described to occur in a variety of cell types (Gundersen et al. 1984; Piperno and Fuller 1985; Geuens et al. 1986; Cambray-Deakin and Burgoyne 1987a, 1987b; Gundersen et al. 1987; Piperno et al. 1987; Arregui and Barra 1990; Nagasaki et al. 1992; Gilmer et al. 1999). However, microtubule dynamics and the distribution of posttranslationally modified microtubules in osteoclasts are less well understood. We report here observations on the differential localization of posttranslationally modified microtubules during the podosome patterning in osteoclasts.
As has previously been demonstrated, microtubule plus-end tracking proteins such as end binding protein (EB) 1 can capture growing microtubules at their plus-end site, and their fast-growing ends are usually directed toward the cell periphery (Vaughan 2005; Akhmanova and Hoogenraad 2005). Besides, EB1 interacts with tyrosinated or detyrosinated tubulin (Peris et al. 2006). For a better understanding of the microtubule dynamics during tubulin posttranslational modifications in osteoclasts, we have investigated microtubule polarity. The visualization of the growing end of microtubules with EB1 thus permits us to identify their polarity. In addition, with regard to histone deacetylase (HDAC), its role in reversible acetylation in transcriptional regulation and histone metabolism has been well documented (for review, see Valenzuela-Fernandez et al. 2008). Furthermore, HDAC6 also functions as a microtubule deacetylase that has effects on various cellular functions dependent on microtubule-mediated processes (Hubbert et al. 2002; Matsuyama et al. 2002). Therefore, we also aimed at clarifying immunocytochemically the localization of HDAC6 during podosome clustering and patterning in osteoclasts.
Materials and Methods
Experimental Animals
All experiments were performed in accordance with the principles and procedures of the Animal Ethics Committee, Asahi University School of Dentistry.
Cell Isolation and Cell Culture
Primary osteoclasts derived from 4- to 6-day-old neonatal rabbits were prepared as described previously (Akisaka et al. 2001). Briefly, the cells were mechanically liberated from the long bone fragments into medium 199 by using a dental spoon excavator. The cell suspensions were applied to coverslips and incubated at 37C in medium 199 containing 10% fetal bovine serum and 100 µl Fungizone. All experiments were performed by using cells cultured on coverslips.
Immunofluorescence for Confocal Microscopy
After 1 to 48 hr of culture on coverslips in medium 199, cells were fixed for 20 min in 4% paraformaldehyde in cytoskeleton buffer containing 10 µg taxol and/or 10 µM phalloidin at room temperature or for 10 min in −30C methanol containing 1% glutaraldehyde. Cells were stained by the double immunofluorescence method. The primary antibodies used were anti-acetylated tubulin at a 1:50 dilution (Sigma-Aldrich, St. Louis, MO), anti-tyrosinated tubulin at 1:500 (Abcam, Cambridge, MA), anti-detyrosinated tubulin (Glu-tubulin) at 1:500 (Abcam), anti–total α- or β-tubulin at 1:100 (Sigma-Aldrich), anti-EB1 at 1:100 (BD Transduction Laboratories, Franklin Lakes, NJ), and anti-HDAC6 at 1:10 (Acris, Herford, Germany). As secondary antibodies, Alexa Fluor 488– or 568–conjugated goat anti-mouse, rabbit, or rat IgG antibodies (all at 1:500; Invitrogen, Carlsbad, CA) were used. All antibodies were incubated with the cells in a humid chamber for 2 hr at room temperature. For F-actin staining, Alexa Fluor 568–labeled phalloidin was used. All confocal images were taken with a Biorad MRC 1024 (Leica, Wetzlar, Germany) and were processed by using Adobe Photoshop (Adobe, San Jose, CA) software. The cells were optically sectioned in x-y planes every 0.3 to 0.5 µm. Micrographs were obtained by superimposition of 2 to 10 optical sections.
Immunogold labeling for electron microscopy
For the ultrastructural visualization of the cytoskeleton, cells on coverslips were treated with a jet stream of buffer solution or with microbubbling generated by sonication with an ultrasonic microprobe, as described previously (Akisaka et al. 2003). By either of these treatments, microtubules and podosome structures were exposed on the protoplasmic surface of the membranes. Briefly, cells on coverslips were washed with cytoskeleton buffer and then fixed for 20 min on ice with a mixture of 4% paraformaldehyde and 1% glutaraldehyde in cytoskeleton buffer containing 10 µg taxol and/or 10 µM phalloidin. Following chemical fixation, double immunochemical procedures were performed with anti-acetylated tubulin and anti-tyrosinated tubulin as described above. Then the cells were incubated with 5- and 10-nm colloidal gold-conjugated antibodies against rabbit or mouse IgG obtained from BBI (Cardiff, UK). After gold labeling, the coverslipped cells were washed with distilled water, quick frozen, and deep-etched or freeze-dried. Finally, they were rotary replicated with 2 nm platinum supported with 20 nm carbon in an Eiko FD-5A freeze-etch device (Eiko Engineering Co., Mito, Tokyo). Stereoview images were taken at 2 different tilt angles (±10 degrees) and converted into digital ones by Adobe Photoshop. Replica images were photographically reversed to aid interpretation; thus, platinum deposits appeared white on a dark background. For three-dimensional (3-D) visualization, we performed reconstruction of 3-D models by using Mex software (Alicona, Grambach, Austria) to create the digitally elevated 3-D model (DEM). These DEM images were rotated freely to obtain highly realistic 3-D images. To obtain clearer visualization of the gold particles in DEM images, we observed images with or without photographic reversal.
Results
Microtubular Organization in Osteoclasts
Osteoclasts that had been freshly plated onto coverslips showed at their leading edge newly formed protrusions that radiated peripherally in all directions. At this time, such newly expanding lamellae or protruding edges were initially devoid of microtubules (Fig. 1A,C). But with time, microtubules appeared; a few microtubules extended into and lay parallel to the expanding edge of the lamella, in which these microtubules seemed to be oriented in the same direction (Fig. 1A). Because of the large number of cytoplasmic structures, including actin filaments or microtubules, individual microtubules were difficult to trace in the central region of the cell. On the contrary, the expanding lamella had just a thin layer of cytoplasm, which allowed us to visualize better the microtubular organization (Fig. 1C,D). Mature quiescent osteoclasts displayed a characteristic system of radially symmetric long microtubules, some straight and some wavy or circular, originating from the centrosomes in the perinuclear region of the cell and extending toward the periphery (Fig. 1B,D). Some osteoclasts contained several scattered podosomes, and others showed clustering of podosomes that subsequently expanded into a ring-like configuration in the cell periphery to form a ring- or belt-like pattern (Fig. 1D). Mature quiescent osteoclasts adhered firmly to the substratum via the podosome rings formed by the clustering podosomes. Multiple widely separated podosomes were connected to each other by the same microtubule. Several microtubules approached from different directions and passed over a given podosome. During the establishment of these podosome rings, the population of microtubules increased gradually and completed the network of microtubules.
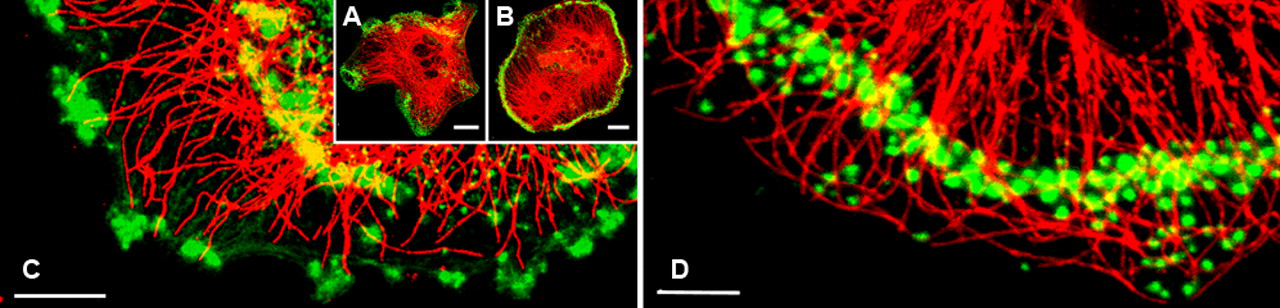
Microtubules and F-actin cytoskeleton of a freshly plated and mature quiescent osteoclasts. (A) In this freshly plated osteoclast, podosome clusters were immunostained with total α-tubulin antibody (shown in red), and F-actin was detected with Alexa Fluor 568–labeled phalloidin (shown in green). F-actin is detected as dot- or belt-like podosomes with Alexa Fluor 568–labeled phalloidin. Note the podosome clusters confined within several protruding edges. Bar = 20 µm. (B) This mature quiescent osteoclast, round in shape, is characterized by the presence of a podosome belt. Most of the microtubules (red) are directed toward the cell periphery where the podosome belt (green) is located. Bar = 20 µm. (C) Note that a few microtubules seem to target several podosomes, which are confined to the newly formed protrusions. Bar = 10 µm. (D) Some microtubules end at the podosome belt, whereas others pass over the belt and intrude into the cell periphery. Individual podosomes also are associated with several approaching microtubules. Bar = 10 µm.
Distribution Pattern of Posttranslationally Modified Forms of Tubulin
To examine the involvement of tyrosination, detyrosination, and acetylation as posttranslational modifications, we compared the localization of tyrosinated, detyrosinated, and acetylated tubulins in the osteoclasts. Tyrosinated microtubules were predominant in the expanding lamella of immature osteoclasts, which reflects their dynamic state required for steering and extension (Fig. 2A). Most of the microtubules all over the cell, even in the expanding lamella, were brightly stained with tyrosinated tubulin antibody, whereas detyrosinated or acetylated tubulin microtubules were restricted to the central part of the cell, where both antibodies labeled the tubulins to the same degree (Fig. 2B). After the formation of marginal cytoplasmic networks of microtubules, the percentage of detyrosinated or acetylated tubulin throughout the mature osteoclast increased. Both detyrosinated and acetylated microtubules showed a very similar distribution pattern (Fig. 2A,C).
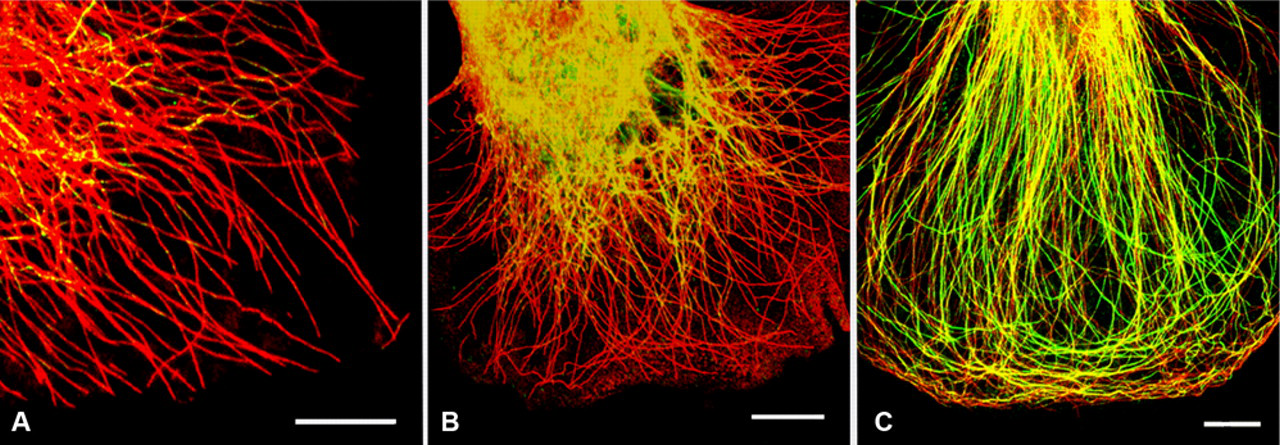
(A) Differential distribution of tyrosinated and acetylated microtubules in an immature osteoclast. The cell was immunostained for tyrosinated tubulin (red) and acetylated tubulin (green). Co-localization of both types is indicated by a variable brilliant yellow color. In comparison with the mature osteoclast (C), tyrosinated tubulins containing microtubules extend into the cellular extrusion, in which the acetylated microtubules are absent. The acetylated microtubules are enriched in the perinuclear region. Bar = 10 µm. (B) The immunofluorescence image shows the differential distributions of tyrosinated and detyrosinated tubulins in an immature osteoclast. Tyrosinated tubulins (red) are predominantly stained throughout the cell, whereas detyrosinated tubulins (green) are absent in the extension area where the tyrosinated microtubules are present. A variable brilliant yellow color indicates co-localization. Bar = 10 µm. (C) Confocal merged image showing differential distribution of tyrosinated (red) and acetylated (green) microtubules in a mature osteoclast. The variable yellow color in the microtubules indicates co-localization. In contrast to the cell in B, the marginal regions where podosome cluster or belts are located contain acetylated microtubules. Bar = 10 µm.
By tracing the growing end of single microtubules, we always detected tyrosinated tubulin subunits, but not detyrosinated or acetylated forms of tubulin, in the microtubules in motile, highly dynamic regions of the osteoclasts (Fig. 3A). Under close examination of the differential distribution of modified tubulins along the individual microtubules, the double labeling with acetylated and tyrosinated tubulin antibodies showed that the distribution of tyrosinated and acetylated tubulin was similar but that some microtubules contained tyrosinated tubulin but none of the acetylated type (Fig. 3B,C). Especially, the long microtubules derived from centrosomes exhibited differential localization of tyrosinated and acetylated tubulins—that is, their growing tips were labeled with tyrosinated tubulin (Fig. 3B). Even tyrosinated tubulin was always detectable in the free microtubules, whereas in some cases, no or only a few microtubules were stained by antibody against acetylated tubulin (Fig. 3C). Non-centrosomal, free microtubules could be seen clearly near the cell margin, including the expanding lamella and in the vicinity of podosomes. These free microtubules were located among the array of long microtubules of variable length and direction in the thin lamella (Fig. 3A).

A confocal merged image showing the differential distribution of tyrosinated (red) and acetylated (green) tubulins. (A) Many long microtubules show variable co-localization of both types of modified tubulins (yellow). Their growing ends are rich in tyrosinated tubulin (red). Non-centrosomal, free microtubules are observed in the extrusion area of the cell periphery (arrows). Free microtubules are stained predominantly with antibody against tyrosinated tubulin. (B) A single long individual microtubule. The growing ends of both types of modified microtubules are usually directed toward the cell periphery and predominantly stained with anti-tyrosinated tubulin. (C) The growing end of this single free microtubule is also stained with anti-tyrosinated tubulin. Bars: A = 10 µm; B, C = 5 µm.
To determine whether tyrosinated or acetylated tubulin was associated with F-actin dots corresponding to podosomal adhesion structures, we stained osteoclasts simultaneously for F-actin and tyrosinated or acetylated tubulin forms (Fig. 4A). An overlay of the immunofluorescence images revealed that tyrosinated tubulin was co-localized with F-actin in the protrusion of the expanding lamella. Acetylated microtubules forming an extensive network were predominant in the osteoclasts that contained a well-developed podosome ring (Fig. 4B).
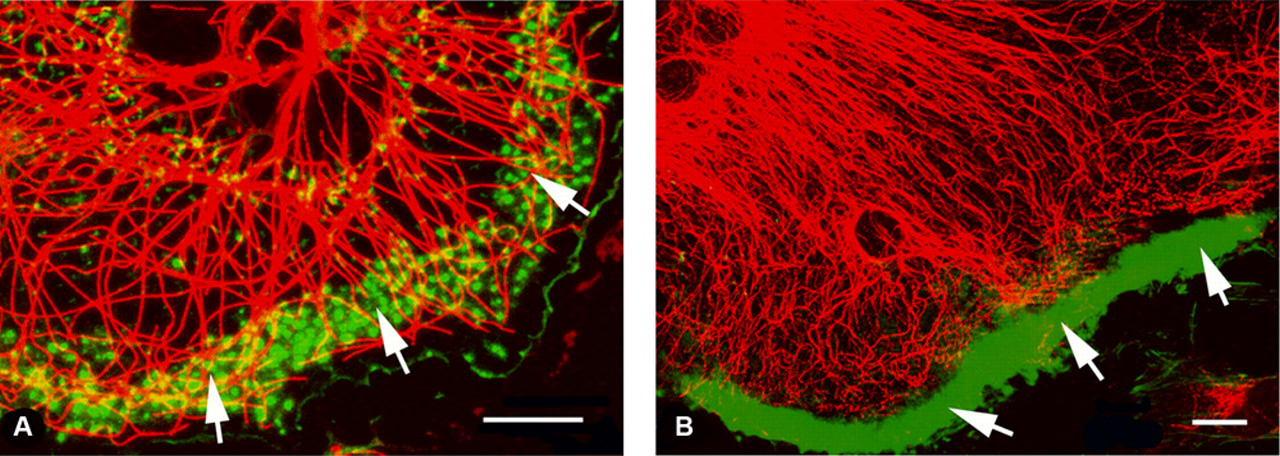
Co-localization of F-actin and modified tubulins (tyrosinated or acetylated tubulins). (A) Tyrosinated microtubules (red) are predominantly found in the podosome clusters (green, arrows). (B) After the formation of the podosome belt (green, arrows) at the cell periphery, stable acetylated microtubules (red) are significantly increased in number. Bar = 10 µm.
To examine the distribution of tyrosinated and acetylated tubulin on individual microtubules, we used sheared-open cells for double labeling at the electron microscopic level. After shearing with a jet streaming buffer or ultrasonication, the attachment membranes and associated cytoskeletal structures were left behind. In sheared-open replicas, we could clearly identify the podosomes, which were composed of an actin cytoskeleton, from 3-D reconstructed images. Judging from deep-etched replicas, the podosomes had two different cytoskeletal organizations, characterized by a tightly packed network of F-actin core. This core was surrounded by sparse F-actin radiating from the core region, which is referred to as an actin cloud (Fig. 5A,B). By tracing the excursions of microtubule ends as they approached and became positioned over the podosomes, in some cases we observed that the growing end of microtubules seemed to directly end at or to make contact with podosomes (Fig. 5A). The use of the immunogold labeling and frozen replica methods confirmed the results obtained from the immunofluorescence localization of modified α-tubulin. Co-localization of tyrosinated and acetylated tubulins was thus demonstrated. Heterogeneous localization patterns of the gold particles may reflect the immunofluorescence labeling. No specific localization of modified tubulins was observed in individual microtubules.
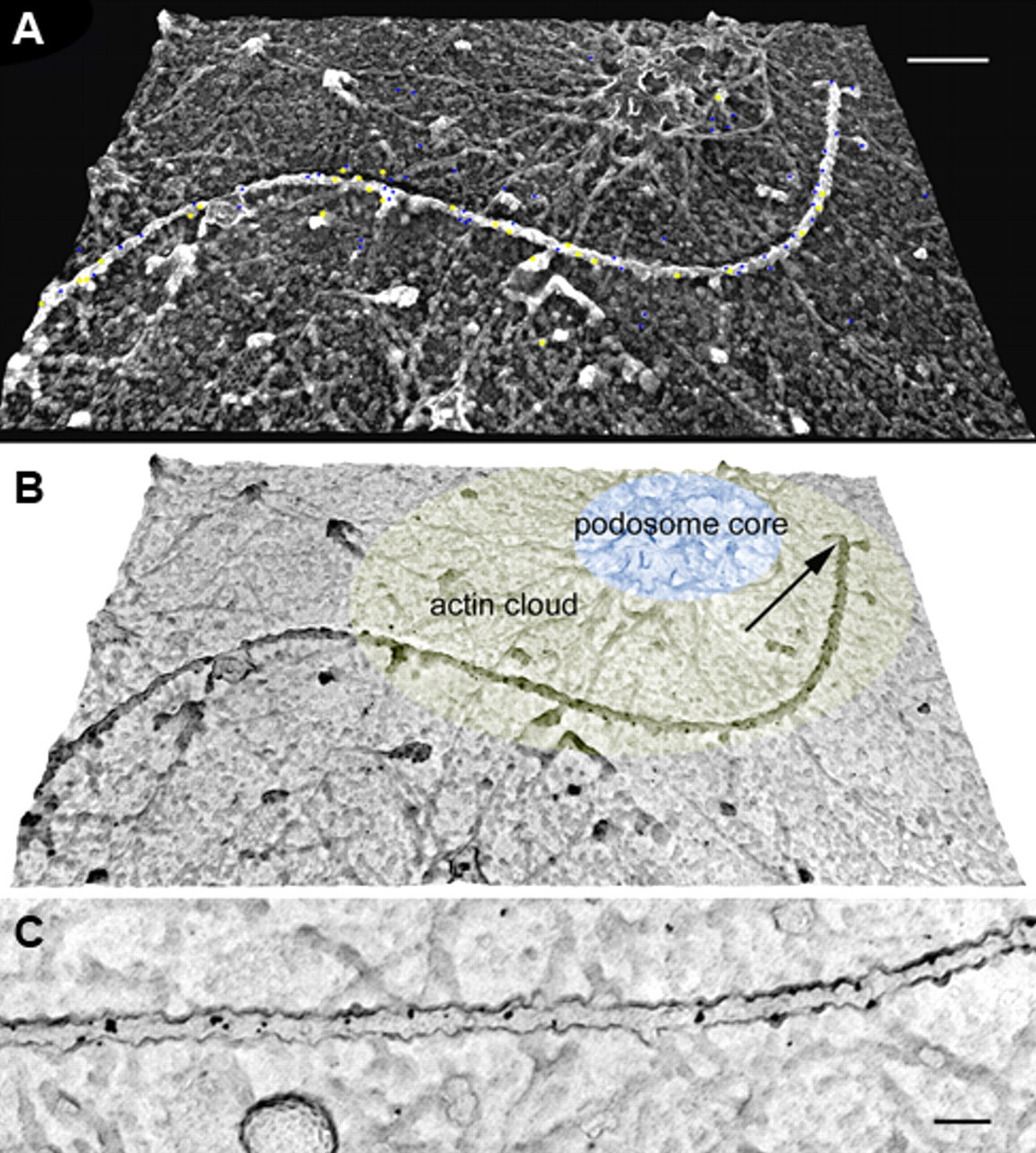
DEM (digital elevation models) of podosome and microtubule labeled with anti-tyrosinated (5-nm gold particles) and anti-acetylated (10-nm gold particles) α-tubulin antibodies. After cell shearing, the inner side of the surface membrane with associated undercoat structures could be visualized. (A) This micrograph was photographically reversed for a more realistic three-dimensional image. The gold particles are pseudocolored in blue (5-nm particles) and yellow (10-nm particles). (B) This image was obtained without photographic reversal, for an easy visualization of the gold particles, which appear in black on a white substrate. The growing tip of the microtubule appears to be closely associated with a podosome (arrow). The single podosome structure is composed of F-actin radiating from the podosome core (pseudocolored in light blue) surrounded by an actin cloud (pseudocolored in light yellow) consisting of a lesser amount of F-actin. Labeling by both types of antibodies is detected along the microtubules. (C) High magnification of a double-labeled microtubule using the two different sizes of colloidal gold particles. Bars: A = 200 nm; C = 100 nm.
Localization of EB-1
The EB1 immunofluorescence was localized to the growing plus ends of microtubules, where it appeared with a comet-like pattern of red front tips with green rear tails along the majority of free and centrosomal microtubules (Fig. 6A,B). EB1 was also localized at the perinuclear region, which is the site of the microtubule-organizing center, the so-called centrosome. The growing ends of cytoplasmic microtubules revealed an intimate relationship with actin filament networks. EB1-positive growing ends of microtubules grew close to the cell margin where the podosomes were located (Fig. 6B). We investigated whether EB1 would bind preferentially to microtubules containing tyrosinated or acetylated tubulin during posttranslational modifications. EB1 was localized predominantly at the growing ends of microtubules containing tyrosinated tubulin (Fig. 6C). The present study clearly identified the localization of EB1 at not only the growing end of centrosomal microtubules but also non-centrosomal, free microtubules (Fig. 6C,D). The polarity of free microtubules showed various orientations with some of them pointing toward the cell periphery and some toward the centrosome.
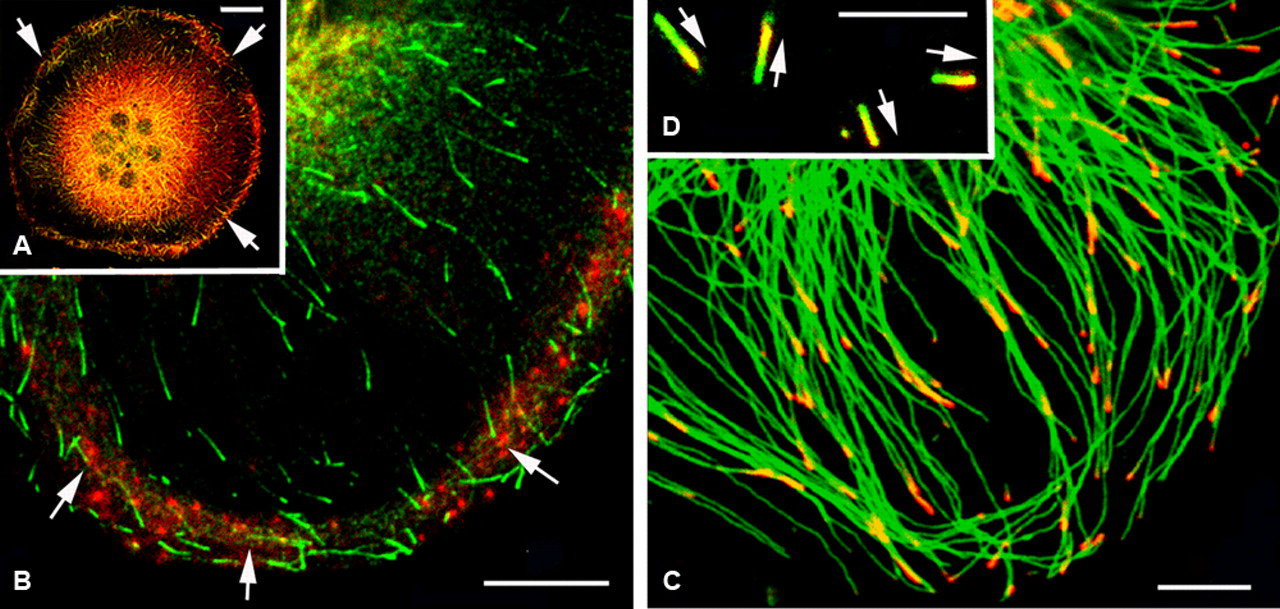
An osteoclast co-immunostained for F-actin of podosomes (red) and for end binding protein 1 (EB1; green) to reveal the growing end of microtubules (green). (A) Fluorescence indicating EB1 was detected in the peripheral podosomal region and also in the perinuclear cytoplasm where the centrosome is located (arrows). (B) EB1 was localized at the growing tips of microtubules, where it appeared as having a short comet-like pattern of brightly fluorescent fronts with dark rear ends along the microtubules. EB1-positive growing tips of microtubules were concentrated at the podosomal regions (arrows). (C, D) Double immunostaining of the cytoplasm of an osteoclast for tyrosinated tubulin (green) and EB1 (red). EB1 was localized at the growing ends of long, centrosomal microtubules (C). Most of their growing ends were directed toward the cell periphery. In free, non-centrosomal microtubules, EB1 was also detectable at their tips, which exhibited various directions (D). Each arrow indicates a direction of growing end of individual free microtubule. Bars: A = 20 µm; B = 10 µm; C = 5 µm; D = 2 µm.
Localization of HDAC6
Actin-rich podosomes were immunopositive for HDAC6, as judged from the results of double labeling with fluorescence-labeled phalloidin and anti-HDAC6 antibody (Fig. 7). Unlike the more widespread distribution of HDAC6 in the perinuclear region, there was a punctate localization concentrated around the cell periphery where the podosomes were located. However, most parts of the cytoplasm, including the leading edges and extending lamella, were negative for HDAC6.
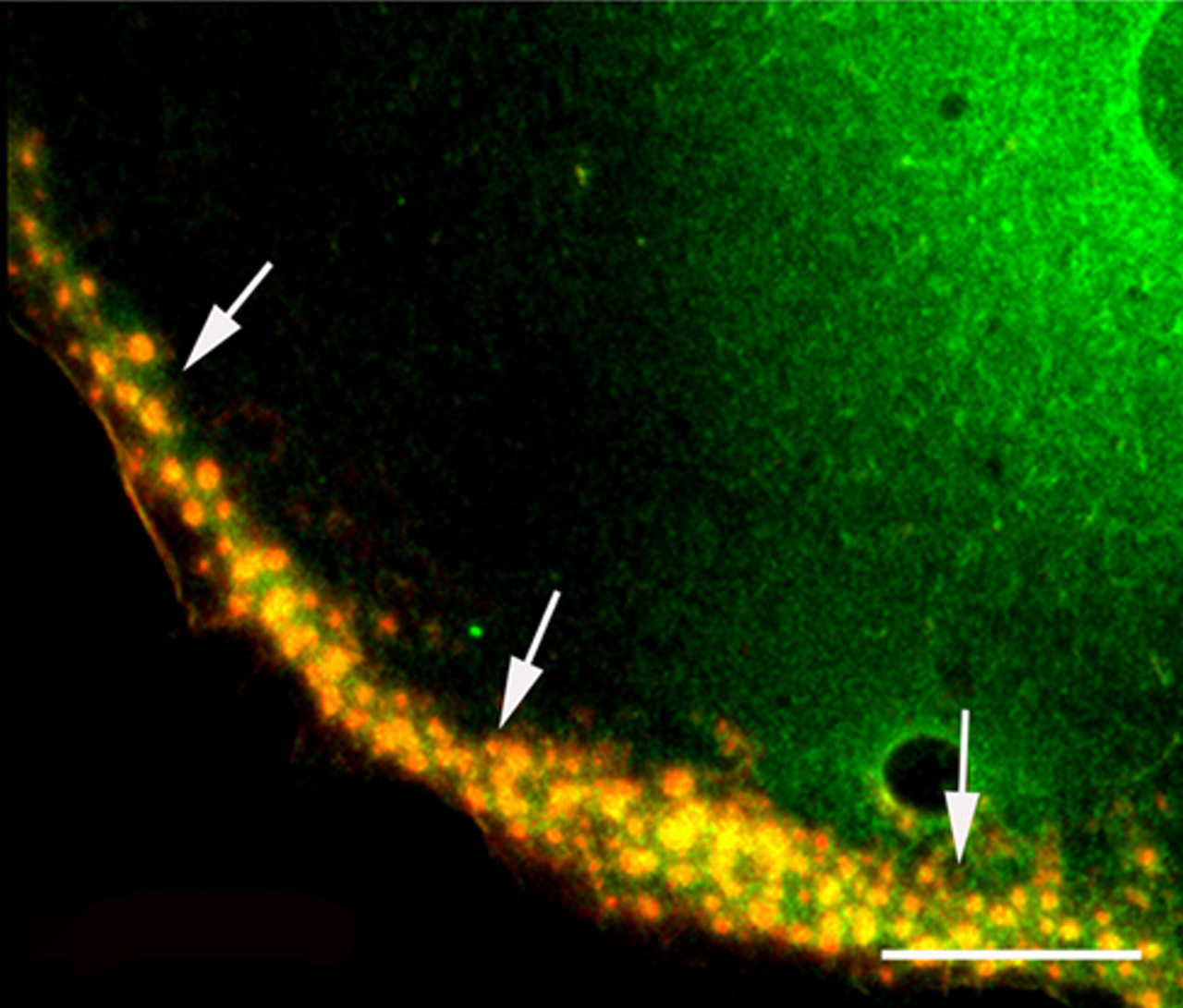
Co-localization of histone deacetylase 6 (HDAC6) and scattered podosomes at the cell periphery. HDAC6, stained with anti-HDAC6 antibody, appears green, and F-actin, stained with phalloidin, is red. The merged image shows a brilliant yellow color, indicating co-localization of HDAC6 and podosomes (arrows). Bar = 10 µm.
Discussion
The morphological and molecular organizations of the podosomes are principally the same as seen under both in vivo and in vitro conditions (Teti et al. 1991; Linder and Aepfelbacher 2003; Jurdic et al. 2006; Luxenburg et al. 2007). When confocal microscopy is used, an individual microtubule can be visualized only in cells widely spread on coverslips because rounded-up cells grown on calcified matrices have many microtubules in their thick cytoplasm.
The functional roles of posttranslationally modified microtubules are proposed to be the regulation of the interaction between microtubules and motor proteins, signal transduction, and organization of microtubules into supramolecular structures. Most microtubules undergo reversible posttranslational modifications such as acetylation/deacetylation or tyrosination/detyrosination. The positive correlation between microtubule stability and acetylation of microtubules in various cells is well documented (Cambray-Deakin and Burgoyne 1987a; Gilmer et al. 1999; Robson and Burgoyne 1989). Acetylated microtubules were first observed in long-lived, stable microtubule populations and seem to be markers for more stable microtubular forms (LeDizet and Piperno 1986; Gundersen et al. 1987; Witte et al. 2008). Microtubules with co-localized acetylated and detyrosinated tubulins have been demonstrated in various types of cells (Cambray-Deakin and Burgoyne 1987a, 1987b; Arregui and Barra 1990; Gilmer et al. 1999; Bulinski and Gundersen 1991). Microtubules are not stabilized directly by detyrosination, but it can be used as a marker for how long a microtubule has been assembled (Webster et al. 1990). Consequently, acetylated and detyrosinated microtubules represent a more stable and/or long-lived microtubule population with a low turnover rate. In contrast, tyrosinated tubulins reflect dynamic microtubules with a high turnover rate (Piperno and Fuller 1985; LeDizet and Piperno 1986; Gundersen et al. 1987; Robson and Burgoyne 1989; Rochlin et al. 1990; Nagasaki et al. 1992).
In osteoclasts at any stage, most microtubules exhibited strong immunoreactivity with antibody against tyrosinated tubulin. In quiescent osteoclasts, microtubules immunoreactive with acetylated tubulin antibody became apparent behind podosome clusters and the podosome ring of the cells. As revealed by the present study, microtubules comprised a continuum of different compositions of tyrosinated and acetylated tubulins in a single microtubule. One possible explanation of these findings is that the co-localization of acetylated and tyrosinated tubulins reflects the rapid turnover of microtubules in the expanding or extrusion region of moving osteoclasts. Long-lived, stable acetylated microtubules increased in number after the establishment of the podosome clustering and ring formation, indicating the possible role of these microtubules for maintenance or support of the podosome rings of quiescent osteoclasts. Furthermore, tyrosinated tubulins are found in highly dynamic microtubules with a high turnover rate (Destaing et al. 2003; Witte et al. 2008). Such dynamic podosomes may coordinate with unstable microtubules to maintain protrusion and support directional migration of the osteoclast. Podosome rings or belts are dependent on microtubules for their stability (Babb et al. 1997; Turksen et al. 1988; Evans et al. 2003; Destaing et al. 2005). In addition, a recent study has shown that microtubule acetylation promotes kinesin-1 motor binding and cargo transport by microtubules (Reed et al. 2006). So, tubulin acetylation would be expected to enhance the intracellular transport of proteins.
Most microtubules are derived from the centrosomes, which are the main microtubule-organizing center in osteoclasts and monocyte-derived giant cells (Moudjou et al. 1989; Mulari et al. 2003). The growth of non-centrosomal, free microtubules in fibroblasts is centered on the peripheral termini of stress fibers corresponding to focal adhesion sites (Kaverina et al. 1998). In the present study, free microtubules were distributed in the protrusion or expanding lamella where podosomes were located. The cytoplasmic origin of free microtubules has been well known to exist in a wide variety of cells (for review, see Bartolini and Gundersen 2006). Most free microtubules were reactive with anti-tyrosinated tubulin antibody, and some of them also reacted with antibody against acetylated tubulin. The question then arises as to whether free microtubules can grow. EB1 binds preferentially to a population containing tyrosinated tubulin, suggesting that EB1 may participate in the generation of new polymeric tubulin and act as a marker of dynamic microtubules (Tirnauer and Bierer 2000). In addition, the localization of EB1 at the microtubule organizing center (i.e., centrosome, which is located in the perinuclear region) suggests its functional role in microtubule nucleation, polymerization, and minus-end anchoring (Berrueta et al. 1998; Louie et al. 2004). EB1 is a member of the family of microtubule plus-end-tracking proteins, which become localized at the growing microtubule plus-end. Possible cross-talk between these proteins and actin-organizing proteins would be suggested in the guidance of microtubules into adhesion foci (Krylyshkina et al. 2003). A pathway in which a crosslinking protein directly links actin to microtubules through interaction with EB and is associated with mDia (Kodama et al. 2003; Slep et al. 2005), which participates in the Rho-mDia2-HDAC6 signaling pathway, controls podosome patterning through microtubule acetylation in osteoclasts as mentioned above. Our observation that EB1 was demonstrated at the growing tips of tyrosinated microtubules indicates that centrosomal and free microtubules maintain their ability for growth and influence microtubule dynamics in osteoclasts. EB1 is a potential candidate to contribute to microtubule stability during podosome clustering and patterning, but its exact functional role remains to be defined.
To explain the relationships among tubulin modification, HDAC6, and podosome formation in osteoclasts, we confirmed that tubulin modifications occurred in various stages of the osteoclast and that HDAC6 was localized in their podosomes. Microtubules affect podosomal adhesion and the direction of cell movement by modulating the stability of cell adhesions and other actin-rich structures through Rho GTPase (Destaing et al. 2003; Destaing 2005). All of these observations suggest that microtubule stabilization can affect podosome formation and distribution during osteoclast maturation. HDAC6 can induce microtubule deacetylation, which regulates cytoskeletal structures, including microtubules. The Rho effector mDia has been implicated in the regulation of posttranslationally modified microtubules (Palazzo et al. 2001). Destaing et al. (2003) revealed that microtubule acetylation for podosome ring or belt stabilization is controlled by the Rho-mDia2-HADC6 pathway in osteoclasts. The present study showed that HDAC6 was localized in the podosomes as well as in the perinuclear region. In accordance with studies showing the localization of HADC6 in F-actin spots of membrane ruffle-forming cells (Serrador et al. 2004) and in such spots in leading lamellipodia (Cabrero et al. 2006), all of these structures are highly dynamic ones in which actin remodeling occurs.
Footnotes
The author(s) declared no potential conflicts of interest with respect to the authorship and/or publication of this article.
The author(s) disclosed receipt of the following financial support for the research and/or authorship of this article: This work was supported in part by the following grants to T.A.: Grants-in-Aid for Scientific Research (C) (no. 21592350) from the Japan Society for the Promotion of Science, the Ministry of Education, Culture, Sports, Science, and Technology.