
Abstract
Postweaning multisystemic wasting syndrome (PMWS) was reproduced in prenatally porcine circovirus 2 (PCV2)-infected pigs by either postnatal infection with porcine parvovirus (PPV) or by immunostimulation. Twenty-four randomly selected piglets from 3 sows, which had been experimentally infected during gestation with PCV2, were randomly divided into 3 groups; group 1 (prenatal PCV2 infection, with postnatal PPV infection), group 2 (prenatal PCV2 infection, with postnatal keyhole limpet hemocyanin, emulsified in incomplete Freund's adjuvant [KLH/ICFA] injection), and group 3 (prenatal PCV2 infection only). Twenty-four randomly selected piglets from 3 uninfected sows were randomly divided into 3 groups; group 4 (no prenatal infection, with postnatal PCV2 and PPV infection), group 5 (no prenatal infection, with postnatal PCV2 infection), and group 6 (negative control pigs). Body weight in negative control pigs (group 6) was increased significantly compared with pigs in groups 1, 2, and 4 at 49, 52, 56, 59, and 63 days of age. The granulomatous inflammatory reaction and lymphoid depletion that are typical lesions in pigs with PMWS were observed in the lymph node of piglets in groups 1, 2, and 4 at 63 days of age. Pigs in group 3 had significantly fewer PCV2-positive cells than those from groups 1,2,4, or 5. When the prenatally PCV2-infected pigs were infected with PPV or injected with immunostimulant in the postnatal period, they developed PMWS. Thus, factors that potentiate the progression of prenatal PCV2 infection to PMWS are postnatal infection with PPV or immune stimulation.
Keywords
Introduction
Porcine circovirus 2 (PCV2) is the primary viral infectious agent of postweaning multisystemic wasting syndrome (PMWS). 1, 5 PMWS is characterized clinically by wasting, decreased weight gain, enlarged lymph nodes, and dyspnea. 4, 6 The characteristic histopathologic lesions in PMWS are lymphoid depletion or granulomatous inflammation (or both) in the lymphoid tissues. 5
Experimental reproduction of PMWS appears to be dependent on immune stimulation or coinfections. PMWS has been reproduced in piglets by inoculation with PCV2 alone or in those coinfected with porcine parvovirus (PPV) or porcine reproductive and respiratory syndrome (PRRS) virus (PRRSV), and also if PCV2-infected piglets are immunostimulated by injections with an immunogen emulsified in an oil-based macrophage-targeted adjuvant. 2, 3, 7, 9, 10, 12, 13, 16– 18
PCV2 infection is also linked to abortion, stillbirths, and mummified fetuses. 15, 21, 22, 25 However, PCV2 typically causes subclinical infections because many if not all farms worldwide are seropositive, while PMWS is manifested in relatively low percentages of pigs in PCV2-infected herds. 19, 24 Recently, it has been reported that PCV2 can infect sows and cross the placenta in late gestation. 11, 22 However, it has not been determined whether prenatally PCV2-infected piglets will develop PMWS in the postnatal period. The objective of the present study was to reproduce PMWS in pigs delivered from sows experimentally infected with PCV2 followed by postnatal infection with PPV or immunostimulation.
Materials and Methods
Experimental design
Six pregnant sows were found to be free of PCV2 by immunofluorescent antibody test, PPV by enzyme-linked immunosorbent assay (ELISA, Svanova, Uppsala, Sweden) and PRRSV-specific antibody by ELISA (IDEXX Laboratories, Westbrook, ME). All were in their first pregnancy. Sows were randomly allocated to an infected (n = 3) or control group (n = 3). Strain SNUVR2003 isolated from the serum of a sow subclinically infected with PCV2 was used in this study. The viral inoculum contained isolate SNUVR2003 at its second passage in PCV-free PK-15 cells. Three pregnant sows were inoculated intranasally with 6 ml of tissue culture fluid containing 1.2 × 105 tissue culture infective doses 50% (TCID50)/ml of SNUVR2003 at 3 weeks before the expected farrowing date. The 3 control pregnant sows were similarly exposed to uninfected cell culture supernatant. Each inoculum was instilled over a period of 4–5 minutes into both nostrils. The sows were housed in different rooms based on infectious status and allowed to farrow naturally, but the farrowings were attended. After delivery, sows were humanely euthanized by electrocution and examined for PCV2 nucleic acid by in situ hybridization. Two newborn piglets from each sow were randomly taken and killed to confirm the congenital infections by in situ hybridization. PCV2 was isolated from superficial inguinal lymph node from all sows and performed sequences previously described. 4, 20
Tissue-culture-propagated PCV2, strain SNUVR000463 (2nd passage), and PPV, strain SNUVR000464 (2nd passage), were used as the sources of viral inocula for postnatal piglets. 12 Twenty-four randomly selected piglets from 3 sows, which had been experimentally infected during gestation with PCV2, were randomly divided into 3 groups of 8 pigs each (groups 1, 2, and 3; Table 1). Twenty-four randomly selected piglets from 3 uninfected sows were randomly divided into 3 groups of 8 pigs each (groups 4, 5, and 6; Table 1). Group 1 piglets were each inoculated intranasally at 28 days of age with 1.3 × 104 tissue culture infective doses 50% (TCID50) of PPV in a 1-ml volume prepared as previously described. 17 In group 2, piglets were injected subcutaneously in the left axilla and left hip with a total of 1.0 mg keyhole limpet hemocyanin, emulsified in incomplete Freund's adjuvant (KLH/ICFA) to a volume of 2.0 ml. The KLH injections were done at 7 days of age and repeated at 21 days of age. In group 3, piglets were not infected any virus and were used as congenital PCV2 controls. In group 4, piglets were each inoculated intranasally at 28 days of age with a 2-ml mixture of 1.2 × 104 TCID50 of PCV2, prepared as previously described, 17 and PPV as described for group 2. In group 5, 8 piglets were each inoculated at 28 days of age with PCV2 as described for group 4, but not with PPV. In group 6, 8 piglets were inoculated intranasally with PCV- and PPV-free PK-15 cell lysates. All 6 groups were housed separately in isolators (2 pigs per isolator) and examined at regular intervals.
Experimental design for pigs to treat with porcine circovirus 2 (PCV2), porcine parvovirus (PPV) and keyhole limpet hemocyanin emulsified in incomplete Freund's adjuvant (KLH/ICFA) used in this study.
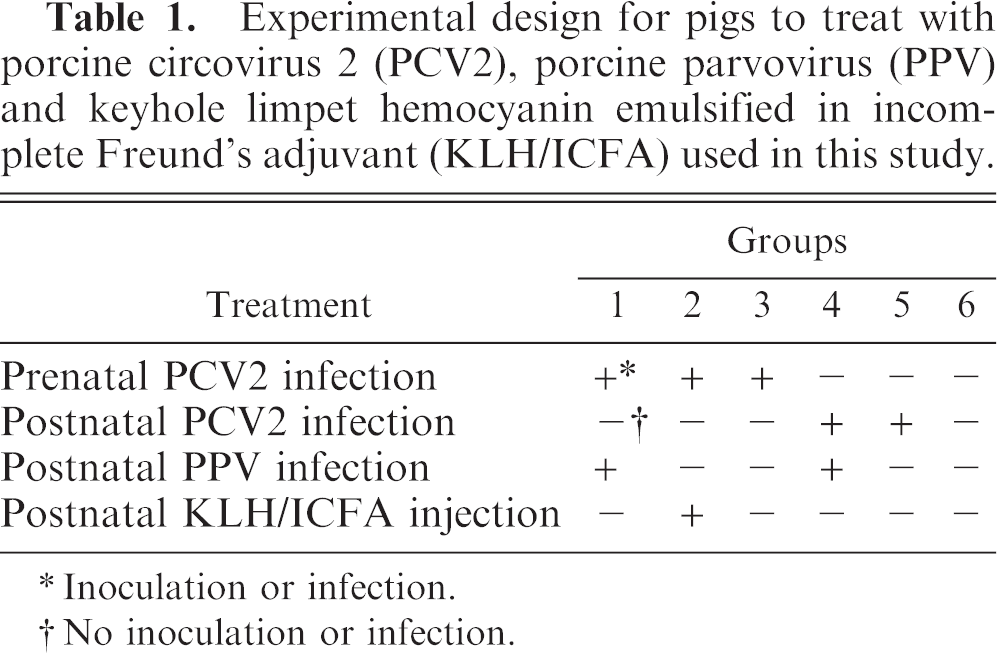
Inoculation or infection.
No inoculation or infection.
Body weight was measured at 21, 24, 28, 31, 35, 38, 42, 45, 49, 52, 56, 59, and 63 days of age. All pigs in each group were euthanized for necropsy at 63 days of age. Superficial inguinal lymph node, lung, liver, kidney, spleen, tonsil, thymus, myocardium, stomach, pancreas, jejunum, and ileum were collected at necropsy and fixed in 10% (v/v) phosphate-buffered formalin for 1–2 days before processing for histopathologic examination. Superficial inguinal lymph nodes were selected for in situ hybridization, as they had previously been found to show consistent and intense labeling for PCV2 and typical granulomatous lesions. 13 The methods were approved by the Seoul National University Institutional Animal Care and Use Committee.
In situ hybridization
Superficial lymph node, spleen, tonsil, and Peyer's patches from each pig and from each infected and uninfected control sow were examined by in situ hybridization. Polymerase chain reaction (PCR) products were purified with a 30-kD cut-off membrane filter. The nucleotide sequences of the purified PCR products were determined by means of BigDye chemistry with the ABI Prism Sequencer (Applied Biosystems, Foster City, CA). Sequencing was performed on the purified PCR products before they were labeled by random priming with digoxigenin-dUTP (Boehringer Mannheim, Indianapolis, IN) according to the manufacturer's instructions. In situ hybridization for PCV2 and PPV was carried out as previously described. 13
Morphometric analysis
Single sections of formalin-fixed hepatic tissues were taken from each of the virus-infected pigs for morphometric analysis as previously described. 13 To obtain quantitative data, morphometric analysis of in situ hybridization slides was performed with the NIH Image J Program (National Institutes of Health, Bethesda, MA). In each case, 3 fields were randomly selected, the number of positive cells per unit area (0.25 mm2) were counted, and the mean values were calculated.
Statistical analysis
Statistical analysis was performed with the InStat package (GraphPad Softwear, San Diego, CA). One-way analysis of variance (ANOVA)/Tukey was used for group comparisons of body weight. A nonparametric Kruskal-Wallis test and Dunn's multiple comparison test were carried out for comparisons of the mean positive cells per unit area for PCV2 among groups. Statistical significance was accepted as P < .05.
Results
Confirmation of PCV2 infection of sows
The 3 infected sows remained clinically normal, did not farrow prematurely, and delivered 31 liveborn and 1 stillborn piglets. PCV2 was isolated from the inguinal lymph node from 3 infected sows. The isolated PCV2 was confirmed as the same virus for propagation of the challenge stock by sequence analysis. By in situ hybridization, PCV2 DNA was consistently detected in the tonsil, spleen, and lymph node from all 3 infected sows. No histopathologic lesions were observed in any tissue examined from infected and control sows. No clinical signs were observed in uninfected control sows, and no virus was isolated from inguinal lymph node in the uninfected control sows. PCV2 DNA was not detected in the tonsil, spleen, and lymph node from all 3 uninfected control sows by in situ hybridization.
Confirmation of prenatal PCV2 infection of newborn piglets
Prenatal infection with PCV2 was confirmed by the detection of PCV2 in the lymphoid tissues of newborn piglets. No histopathologic lesions were seen in 2 piglets each from the PCV2-infected sows and uninfected control sows. PCV2 DNA was detected by in situ hybridization in lymph node, spleen, tonsil and Peyer's patches of small intestine from all 6 prenatally PCV2-infected piglets from each of the PCV2-infected sows, but not in the 6 piglets from the uninfected control sows. Hybridization signals of PCV2 were seen occasionally in lymphoid tissues in prenatally PCV2-infected piglets.
Gross lesions
Gross lesions of PMWS were observed in all 24 pigs from groups 1, 2, and 4 only. Tracheobronchial, mesenteric, and mediastinal lymph nodes were moderately enlarged and pale in pigs from groups 1, 2, and 4 at 63 days of age. No gross lesions were observed in pigs from groups 3, 5, and 6 at 63 days of age.
Body weight
All 24 pigs from groups 1, 2, and 4 were seen to be thin and unthrifty at 45 days of age (17 days after administration of PPV or KLH/ICFA). Four pigs from group 2 and 3 pigs from group 4 became anorectic at 52 days of age (24 days postinoculation). These pigs were recovered slightly without treatment until necropsy at 63 days of age. Sixteen pigs from group 3 and 5 were seen to be slightly thin at 49 days of age (20 days postinoculation). Significant differences in body weight were not detected among the 6 groups at 28 days of age. Body weight in control pigs from uninfected control sows (group 6) were increased significantly compared with pigs in groups 1 and 2 (prenatal PCV2 infection, with postnatal PPV infection or KLH/ICFA injection, respectively) and in group 4 (no prenatal infection, with postnatal PCV2 and PPV infection) at 49, 52, 56, 59, and 63 days of age. Body weight in pigs in group 3 (prenatal PCV2 infection only) was also increased significantly compared with pigs in groups 2 and 4 at 59 and 63 days of age (Fig. 1).
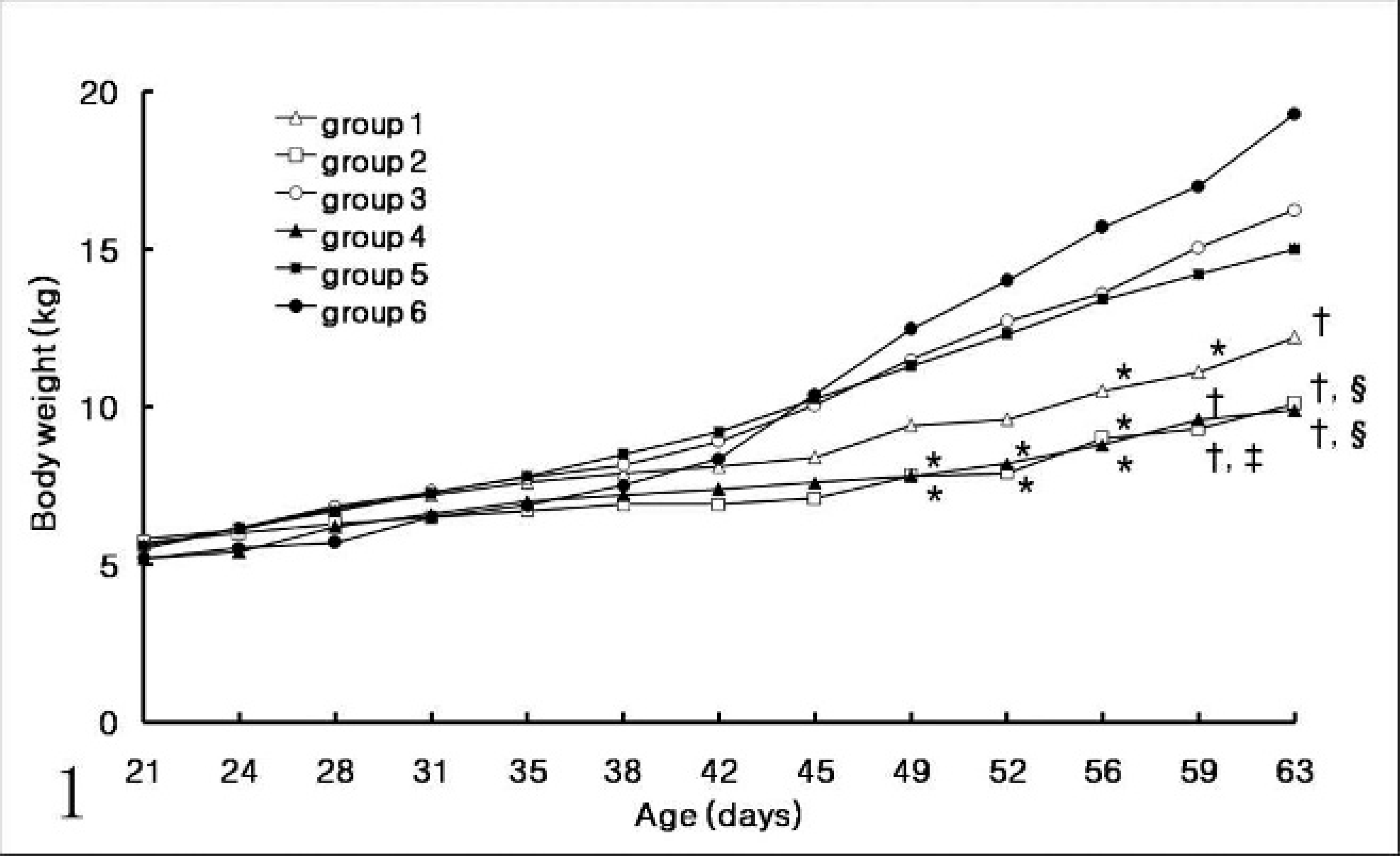
Body weight in pigs infected prenatally with porcine circovirus 2 (PVC2) (groups 1–3) with postnatal porcine parvovirus (PPV) infection (group 1), immunostimulation with KLH/ ICFA (group 2), or no additional stimulation (group 3); and in pigs from the uninfected control sows (groups 1–3) with postnatal PCV2 and PPV coinfection (group 4), postnatal PCV2 infection alone (group 5), and uninfected control pigs (group 6). Body weight in group 1 was increased significantly compared to other groups (∗P < .05; †P < .001). Body weight in group 3 was increased significantly compared to other groups (‡P < .05; §P < .001).
Histopathology
The granulomatous inflammatory reaction and lymphoid depletion that are typical lesions in pigs with PMWS (Fig. 2) were observed in the lymph node of piglets in groups 1 and 2 (prenatal PCV2 infection, with postnatal PPV infection or KLH/ICFA injection, respectively) and in group 4 (no prenatal infection, with postnatal PCV2 and PPV infection) (Table 2). Lymph nodes were depleted of mature lymphocytes and contained pyknotic basophilic nuclear material and adjacent karyorrhectic debris; germinal centers were reduced or absent. Multifocal granulomatous inflammation was characterized by the presence of epithelioid macrophages and multinucleated giant cells. Lymphoid depletion was seen in these lymph nodes. Reactive lymphoid hyperplasia and germinal center formation were present. Lymph nodes contained pyknotic basophilic nuclear material and adjacent karyorrhectic debris. Intracytoplasmic inclusion bodies were often seen in the lymph node in pigs from group 4. No intracytoplasmic inclusion bodies were observed in the lymph node in all pigs from any other groups. No histopathologic changes were seen in piglets from groups 3, 5, and 6.
Results of granulomatous inflammation and porcine circovirus 2 (PCV2) detection by in situ hybridization in 8 pigs from each group.
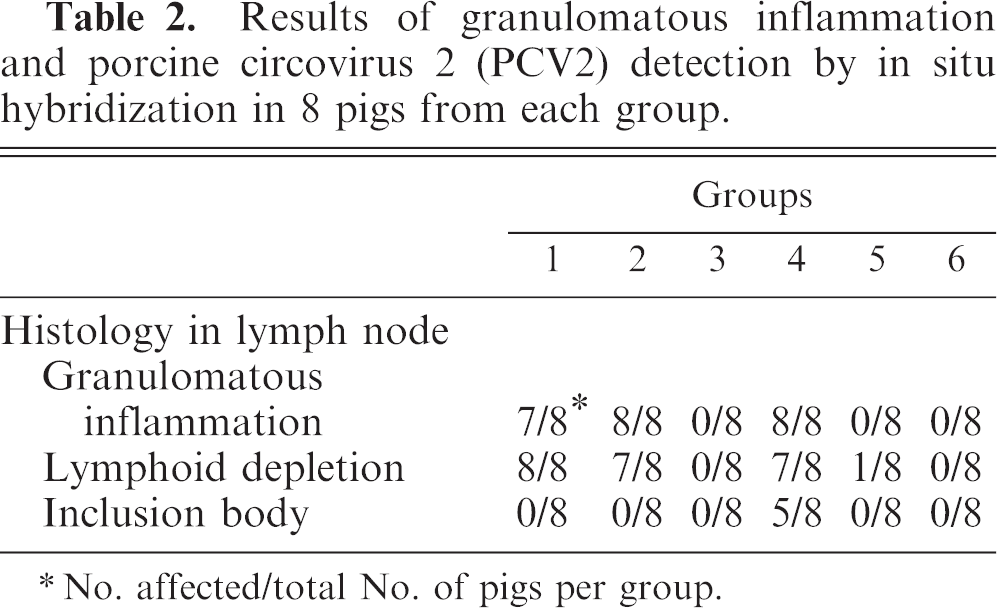
No. affected/total No. of pigs per group.
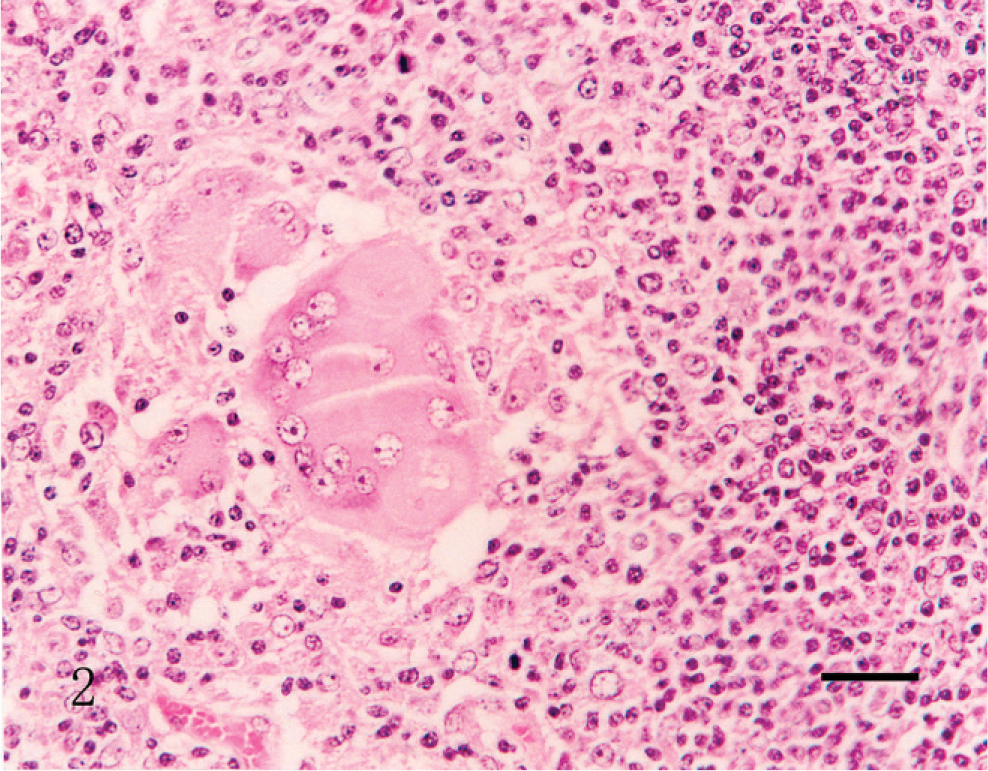
The pig infected prenatally with porcine circovirus 2 with postnatal porcine parvovirus infection (group 1), and euthanized at 63 days of age. A cluster of histiocytes and multinucleated giant cells are seen in lymph node. HE. Bar = 55 µm.
Porcine circovirus 2 distribution by in situ hybridization
PCV2 DNA was detected in all lymph node, spleen, tonsil and Peyer's patches of small intestine from all pigs infected prenatally with PCV2 (groups 1–3) with postnatal PPV infection (group 1), immunostimulation with KLH/ICFA (group 2) or no additional stimulation (group 3) and in pigs from uninfected control sows (groups 4 and 5) with postnatal PCV2 and PPV coinfection (group 4) and postnatal PCV2 infection alone (group 5). The morphology of host cells was preserved despite the relatively high temperature required in parts of the incubation procedure. The signal intensity varied within and between histologic structures in any one section and between pigs. Positive cells typically exhibited a dark brown labeling, mainly in the cytoplasm but occasionally in the nucleus, without background staining.
A strong hybridization signal for PCV2 was detected in the cytoplasm of epithelioid macrophages and multinucleated giant cells in the inguinal lymph nodes from pigs in groups 1, 2, and 4 (Fig. 3). Scattered macrophages exhibited a less intense and dispersed hybridization signal in the cytoplasm in the inguinal lymph nodes from pigs in groups 3 and 5. No hybridization signal was seen in tissue sections pretreated with DNase A. Sections from the 8 control piglets from uninfected control sows (group 6) showed no hybridization signal for PCV2.
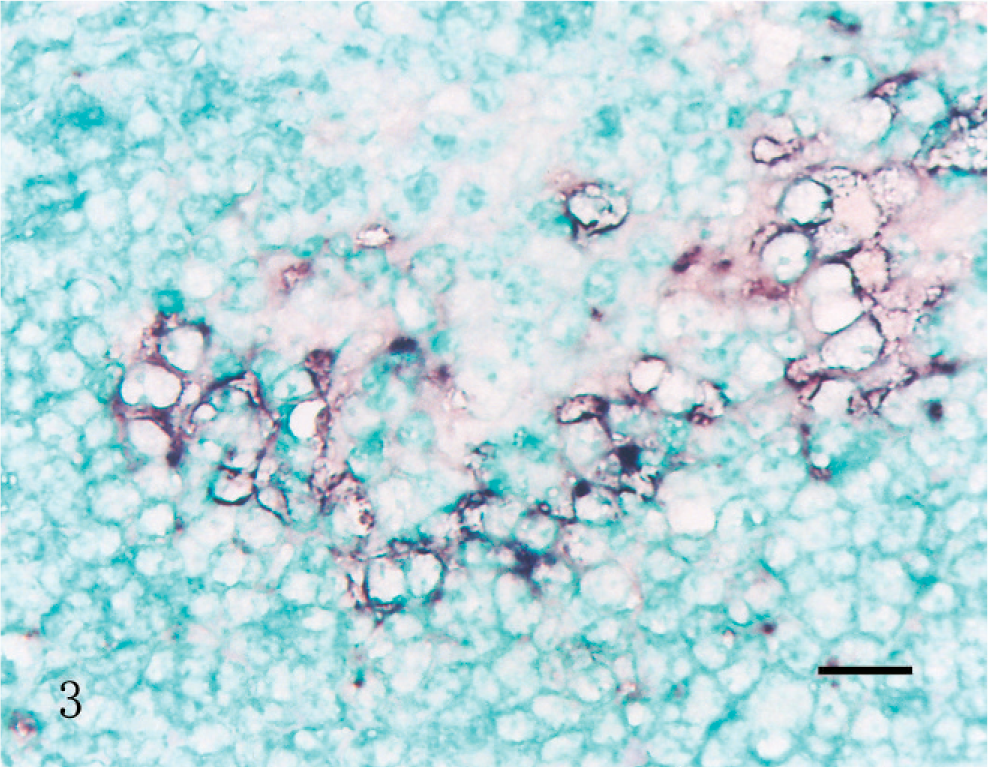
The pigs infected prenatally with porcine circovirus 2 with postnatal immunostimulation with KLH/ICFA (group 2), and euthanized at 63 days of age. PCV2 DNA (indicated by the blue–black labeling) was detected in macrophages in the lymph node. In situ hybridization; DNA probe; nitroblue tetrazolium/5-bromocresyl-3-indolylphosphate, methyl green counterstain. Bar = 30 µm.
Morphometric analysis
Statistical analysis of the mean number of PCV2-positive cells per unit area for inguinal lymph node showed significant differences among groups. Pigs in group 3 (prenatal PCV2 infection only) had significantly fewer PCV2-positive cells than those from groups 1, 2, 4, or 5. Pigs infected prenatally with PCV2 only (group 3) had significantly less PCV2-positive cells than those in pigs infected prenatally with PCV2 with postnatal PPV infection (group 1), immunostimulation with KLH/ICFA (group 2) and in pigs from uninfected control sows (groups 4 and 5) with postnatal PCV2 and PPV coinfection (group 4) and postnatal PCV2 infection alone (group 5) (Fig. 4). There was no difference for PCV2-positive cells from piglets between groups 1 and 4 or between groups 2 and 5.
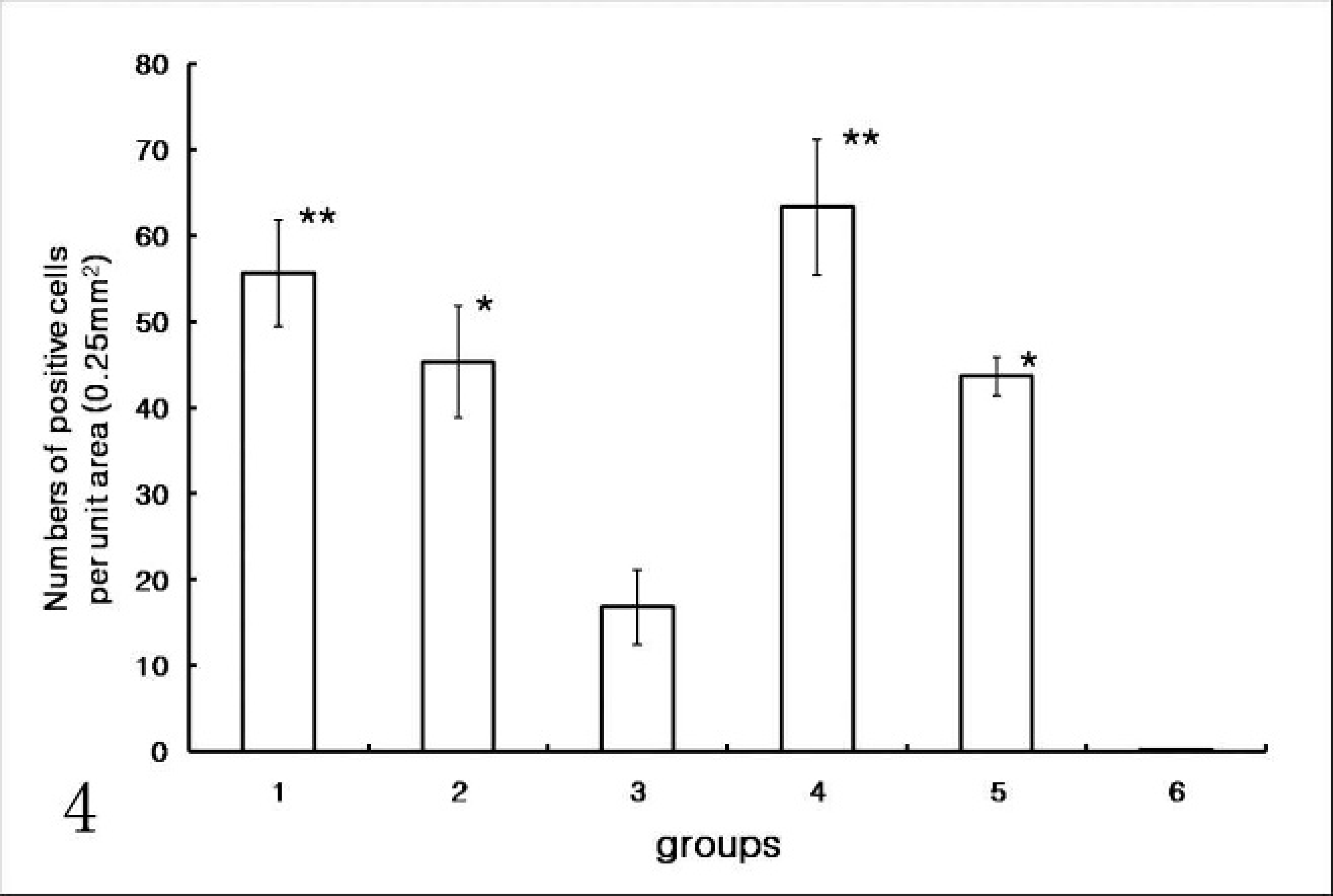
Numbers of porcine circovirus 2 (PCV2)-positive cells in inguinal lymph node in in pigs infected prenatally with porcine circovirus 2 (PVC2) (groups 1–3) with postnatal porcine parvovirus (PPV) infection (group 1), immunostimulation with KLH/ICFA (group 2), or no additional stimulation (group 3); and in pigs from the uninfected control sows (groups 4–6) with postnatal PCV2 and PPV coinfection (group 4), and postnatal PCV2 infection alone (group 5), and uninfected control pigs (group 6). Pigs infected prenatally PCV2 alone (group 3) had significantly less PCV2-positive cells than those from groups 1, 2, 4, 5, or 6 (P < .001∗).
Porcine parvovirus distribution by in situ hybridization
PPV DNA was detected by in situ hybridization in lymph node, spleen, tonsil, and Peyer's patches of small intestine from all pigs infected prenatally PCV2 with postnatal PPV infection (group 1) and all pigs from the uninfected control sows with postnatal postnatally PCV2 and PPV coinfection (group 4). Scattered macrophages exhibited a strong intense and dispersed hybridization signal in the cytoplasm in the inguinal lymph nodes from pigs in groups 1 and 4. No PPV DNA was detected in any tissues from other groups (group 2, 3, 5, and 6) because PPV was not inoculated in pigs from these groups.
Discussion
The results of present study demonstrate that prenatal infection with PCV2 alone does not induce PMWS in the postnatal period. However, stimulation of prenatally PCV2-infected piglets by either KLH immunostimulant or infection with PPV induced the typical PMWS-associated microscopic lesions of granulomatous inflammation and lymphocyte depletion in lymph nodes. Under conventional husbandry conditions, coinfections and immunostimulation are common and may account for the overt expression of PMWS in the postnatal period, in piglets that were infected in utero with PCV2. Transplacental PCV2 infection, as demonstrated in this and other studies, 11, 22 may not only cause reproductive failure in the prenatal period, but also may lead to the development of multisystemic disease in later life. Since subclinically infected sows are highly prevalent, there is high possibility of transplacental infection of fetuses with PCV2 in late gestation. Therefore, transplacental PCV2 infection may play an important role in development of clinical PMWS in the postnatal period.
The diagnosis of PMWS requires the presence of three criteria: 1) compatible clinical signs, 2) characteristic microscopic lesions, and 3) PCV2 within the lesions. 1, 5 PCV2-associated microscopic lesions consistent with PMWS were observed in piglets from PCV2-infected sows (groups 1 and 2) only if they were postnatally challenged with PPV or KLH. Although the body weight from piglets which were prenatally infected with PCV2 but not otherwise stimulated (group 3) was decreased significantly compared with that from uninfected piglets (group 6), these piglets were not considered as having PMWS because characteristic microscopic lesions were not seen. However, these pigs (group 3) may develop PMWS once they are infected with PPV or immunostimulated in the postnatal period.
PPV has been used as a model coinfecting agent with PCV2 to induce PMWS in experimental studies. Pigs experimentally infected with PCV2 displayed clinical signs and lesions typically associated with this virus, and coinfection of pigs with PCV2 and PPV appeared to result in wasting disease. 2, 9, 17 PPV was demonstrated in 25.6% of the cases of PMWS examined in the Republic of Korea 14 and in about 18% of the cases of PMWS in Canada. 8 Since PPV is endemic in the swine population, many weaned pigs, including those that develop PMWS, may be infected with PPV at a time when maternal antibodies to PPV are in decline. 23 Therefore, it is possible that postweaning infection with PPV in prenatally PCV2-infected pigs may induce the PMWS in the field.
Besides PPV infection, immune stimulation is a key component of the pathogenesis of PCV2-associated PMWS in swine. Severe clinical PMWS has been reproduced in gnotobiotic pigs inoculated oronasally with PCV2 at 1 day of age and then parenterally stimulated by KLH emulsified in ICFA. 16 In the present study, prenatally PCV2-infected piglets developed PMWS when prenatally PCV2-infected piglets were stimulated by KLH and ICFA. Although the degree of immune stimulation needed to convert PCV2-infection from a limited, self-contained, and subclinical infection into clinical PMWS in weanling piglets is not known, frequent immune stimulation by vaccination may promote development of clinical PMWS in piglets that were previously subclinically infected with PCV2.
The effect of the sow should be built into the model for statistical analysis. The effect of the sows of origin on growth rates of the pigs could well be an important factor because the assignment of pigs to groups was truly random. Analysis of a more general model using sow as an effect may partially account for this effect. However, a limitation of the method is that it will be impossible to distinguish whether low growth rates in prenatally PCV2-infected piglets is the result of this infection or an effect of the sow of origin.
In conclusion, prenatal or postnatal PCV2 infection did not by themselves induce lesions of PMWS, but PPV infection or immune stimulation were found to stimulate the progression of prenatal PCV2 infection to PMWS in postnatal period. PMWS is a disease complex influenced by many factors, and piglets infected with PCV2 in either the prenatal or the postnatal period may be susceptible to development of PMWS in response to immune stimulation and PPV infection in the postnatal period.
Footnotes
Acknowledgements
The research was supported by contract research funds of the Research Institute for Veterinary Science (RIVS) from the College of Veterinary Medicine and by Brain Korea 21 Program for Veterinary Science in Republic of Korea.