
Abstract
Female dogs are frequently affected by mammary tumors, both carcinomas and sarcomas. The mechanisms behind mammary-tumor formation and the high degree of heterogeneity are not understood. To provide insight into this issue, it is important to determine the properties of the cells forming the different types of tumors. One question is if individual neoplastic cells can give rise to phenotypically distinct tumor types, i.e., show plasticity. We studied 3 different tumors (a spindle-cell tumor, an osteosarcoma, and a carcinoma) and followed the change of lineage marker expression between the primary canine mammary tumors, the clones derived from the corresponding tumors and in tumors generated after inoculation of tumor clones into nude mice (n = 75). Inoculation of clones derived from the spindle-cell tumor gave rise to spindle-cell tumors in nude mice. Several of these contained bone tissue, a sign of plasticity. Clones derived from the osteosarcoma were negative for a panel of lineage markers but, when inoculated into nude mice, they were able to form bone, again a sign of plasticity. In contrast to the primary carcinoma, most of the clones derived thereof lacked keratin expression, but keratin expression was recovered in most of the tumors formed after inoculation of clones into nude mice. Moreover, tumors generated from the carcinoma clones, in contrast to the primary tumor, were positive for smooth-muscle-cell markers. Our results point to plasticity in canine mammary tumors, as shown both by morphologic criteria and by expression patterns for lineage specific markers.
Spontaneous mammary tumors, either of a malignant or benign type often develop in female dogs. Of the malignant mammary tumors, carcinomas are most common, but also sarcomas, such as fibrosarcomas, osteosarcomas, and carcinosarcomas occur. 2, 23, 24 An important question under debate regards the origin of the different canine mammary tumors, in particular, the origin of cartilage and bone in sarcomas and mixed tumors. The expression of lineage markers has been studied to a large extent in primary tumors and has revealed a great diversity of expressed lineage markers in mammary tumors. 6, 12, 13, 32 In dogs, myoepithelial cells have been suggested to be the origin of mammary tumors, as shown by the expression of a myoepithelial-cell marker, maspin in a majority of both benign and malignant canine mammary tumors. 7 There is also an increasing interest for the role of myoepithelial cells in human breast cancer. 20, 21, 27, 29 A key to the understanding of cellular origins of the mammary tumors is to use experimental setups. One approach is to study lineage-characteristic markers and to see how tumor cells express markers before and after they form new experimental tumors.
Conceivably, the phenotype of a tumor, as determined by the expression of lineage markers, could reflect the phenotype of the parent-cell type, i.e., before neoplastic transformation. Alternatively, the parent cell could, during the course of tumor genesis change its phenotype, i.e., exhibiting plasticity. In a previous report, we demonstrated that individual cell lines established from different types of canine mammary tumors could give rise to phenotypically different tumors when inoculated into nude mice, which could be explained by phenotypic plasticity. 13 However, it could not be ruled out that the used mammary-tumor cell lines were heterogeneous, i.e., that tumor cells of different phenotypes were present in the parent-cell line. To investigate, if indeed one single cell can give rise to phenotypically distinct tumors, we here instead used cloned cell lines from 3 different canine mammary tumors: a spindle-cell tumor, a combined osteosarcoma, and a scirrhous carcinoma. Cloned tumor cells were inoculated into nude mice and the phenotypes of the generated tumors were studied. Our results indeed conform to the notion that individual tumor cells can generate phenotypically different tumors in vivo.
Methods
A primary osteosarcoma and a primary carcinoma were collected from a 5-year-old female Papillon during surgical treatment. The dog had 2 mammary tumors on the right side. In the first gland (cranial thoracic), an 8 × 7 × 7-cm ulcerated tumor with central necroses was found. Pleomorphic tumor cells and osteoid formation comprised the tumor. In addition, foci with a chondroid differentiation were also seen. The tumor was diagnosed as a poorly differentiated combined osteosarcoma according to the World Health Organization classification of canine mammary tumors. 24 In the 4th mammary gland (cranial inguinal), a smaller tumor, approximately 2 cm in diameter, adjacent to the nipple was found. Poorly differentiated tumor cells formed a duct-like growth in a desmoplastic stroma, and this tumor was diagnosed as a simple scirrhous carcinoma. The dog was euthanatized 6 months after surgery because of lung metastases and was necropsied. The tumors in the lungs and 1 kidney were combined osteosarcomas, with a poor to a high degree of differentiation, and they were considered metastases from the surgically treated mammary osteosarcoma. There was no sign of any primary osteosarcoma in the dog as seen by an X-ray of the skeleton included in the necropsy. No carcinomatous metastases were found from the postmortem examination.
Establishment of cell lines from the primary osteosarcoma and carcinoma
For tissue culture, pieces of fresh tumors were rinsed in antibiotics (120 µg/ml penicillin and 100 µg/ml streptomycin; National Veterinary Institute, [SVA], Uppsala, Sweden), cut into small pieces, and treated with collagenase overnight. 22 The tumor fragments and established cell lines were cultured in RPMI 1640 medium (Invitrogen, Carlsbad, CA), supplemented with 2 mM L-glutamine (SVA), 10% fetal bovine serum (Invitrogen), and antibiotics as above. The cells were incubated at 37°C in 5% CO2. The cell line generated from the osteosarcoma was named CMT U353 B (combined osteosarcoma cell line) and the cell line from the scirrhous carcinoma was named CMT-U353 H4 (scirrhous carcinoma cell line).
Cloning of cell lines
In addition to the 2 new cell lines CMT-U353 B and CMT-U353 H4 (see above), CMT-U309 (spindle-cell tumor cell line) was used. The latter was established from a spindle-cell tumor as previously described. 13 Single cells with large distance to neighbors were marked and later picked. The cell-culture plates were either not treated, coated with poly-L-ornithine (Sigma-Aldrich, St Louis, MO), or with a combination of poly-L-ornithine and fibronectin (Invitrogen). Colonies from all 3 coatings were picked. Initially, 7 clones of CMT-U353 B, 14 clones of CMT-U353 H4, and 10 clones of CMT-U309 were picked. The procedure of cloning was repeated once for clones A5 and C6 from the CMT-U309 cell line.
Immunocytochemistry
Cytospin slides were prepared from the clones at passages 2–8. Slides were air-dried at room temperature and stored at −70°C. Normal tissue used as positive controls (see below) were sectioned at 5 µm in a cryostat at −25°C. All slides were air-dried at room temperature and frozen at −70°C until used. The slides were fixed in acetone and washed in 0.05 M Tris buffer, pH 7.6. The tumor and control tissues were fixed in 4% phosphate buffered formaldehyde formalin. Bone tissues were decalcified for 30 minutes to 2 hours (Parengys, Bie and Berntsen A-S, Denmark) before embedded in paraffin. The slides were placed in antigen unmasking solution (VECTOR, Burlingame, CA) and boiled in a decloaking chamber (Biocare Medical). The antigen retrieval for the collagen antibodies was done by incubating the slides 30 minutes at 37°C in 0.01 M HCl/34 units/µl pepsin (Pepsin A, Sigma).
Monoclonal antibodies toward smooth-muscle actin (SMA) (clone 1A4), vimentin (clone V9), desmin (clone D33), neurofilaments (clone 2F11), p63 (clone 4A4), keratin 5, 6, 18 (clone LP34), keratin 1–8, 10, 13–16, and 19 (AE1/AE3; pan-keratin) were from Dako Glostrup, Denmark). Antibodies to skeletal myosin (clone MY-32, monoclonal, Sigma), CD31 (polyclonal, Santa Cruz), type I collagen (clone COL-1, monoclonal, Abcam, Cambridge, UK) and type II collagen (clone COLL-II, monoclonal, Chemicon) were from the sources indicated. For every immunohistochemical analysis, positive and negative controls were used. As positive controls, canine spinal cord was used for neurofilaments, canine striated muscle for skeletal myosin, and canine bone for type I and type II collagens. For all other antibodies, canine mammary gland was used as positive control. The corresponding immunoglobulin G isotypes and concentrations were used as negative controls for the monoclonal antibodies. All slides were incubated overnight (4°C) with the primary antibody. The ABC-Elite system was used (Vector Laboratories, CA) and 3-amino 9-etylcarbazole (SVA) was used as chromogen for the cytospin specimens whereas Nova RED or 3,3'-diaminobenzidine (Vector Laboratories) was used as chromogen for the paraffin-embedded tumors. All immunocytochemistry and immunohistochemistry were repeated at least once.
Western blot analysis
After washing with ice-cold phosphate buffered saline solution (PBS), the cells were lysed by addition of 500 µl boiling 1% sodium dodecyl sulfate (SDS), 10 mM Tris-HCl, pH 7.4. The lysates were then boiled for an additional 5 minutes, passed several times through a 22-gauge (0.7-mm) needle and centrifuged for 5 minutes. Protein concentration was determined with Pierce's protein assay (BCA, Pierce Chemical Co., Rockford, IL). The samples were separated by gradient SDS-polyacrylamide gel electrophoresis (NuPAGE Novex 4–12% Bis-Tris gel, Invitrogen, San Diego, CA) according to the protocol provided by the manufacturer. Proteins were transferred to a nitrocellulose filer (Hybond ECL, Amersham Biosciences, Uppsala, Sweden). The filter was blocked with 5% bovine serum albumin in TBS-T (10 mM Tris-HCl, pH 7.7, 0.15 M NaCl, 2% Tween-20) and incubated with the primary monoclonal antibodies against vimentin (clone V9, Dako) and beta Actin (ab20272, Abcam, Cambridge, UK) at +4°C overnight. The filter was developed by using a substrate solution (chemoluminescence, Super Signal, Pierce Chemical) on X-ray films (Hyperfilm ECL, Amersham Biosciences). Stripping of the filter was done in a solution that contained 110 mMß-mercaptoetanol, 2% SDS, and 62.5 mM Tris-HCl at pH 6.7. The tumor cells were used at passages; 9 for the CMT-U353B clones 1, 2, 3, 6, and 7, and at passages 8 and 10 for the 353H4 clones 5, 6, 10, and 12. As control cells Madin-Darby Canine Kidney Cells (MDCK) at passage 83 and canine dermis at passage 2 were used. The Western blot analysis was repeated 4 times for the CMT-U353B clones and twice for the CMT-U353H4 clones.
Flow cytometry
Cells (passages 4–7) were counted by using a Bürker chamber, frozen in citrate buffer, and stored at −70°C. The staining of the cells was done according to Vindel⊘v et al. 31 Briefly, cells were thawed and 2.5 × 106 cells of each clone were mixed with an internal standard of chicken red blood cells (CRBC). Canine blood cells (CBC) with nucleated leukocytes mixed with CRBC were used as external standard. The cells were treated with trypsin to enable staining of cell DNA with propidium iodide (Sigma). The analysis was done with a FACSort (Becton Dickinson) and 40,000 cells were counted from each analysis. The DNA index (DI) was calculated by dividing the ratio of the clones and the internal CRBC with the ratio of the external standard (CBC and CRBC). Each clone was analyzed at least 3 times, and the DI was calculated as a mean value. The coefficient of variation (CV) was calculated with the program ModFit LT version 3.0 for clones from CMT-U309 and CMT-U353 H4. To test the resolution of the method, the 353 B clones were mixed with normal CBCs and the CV for the clone peak was estimated with the program BD Cell Quest Pro version 4.0.1. The bar was set at half the height of the peak.
Tumorigenicity in nude mice
Five clones from each cell line were subcutaneously inoculated in 5 female Balb/c nu/nu mice (6 weeks old; Bomholtgaard Denmark). From CMT-U309, the clones 1, 2, 4, A5, and C6 were inoculated (n = 25); from CMT-U353 B, the clones 1, 2, 3, 6, and 7 (n = 25); and, from CMT-U353 H4, the clones 5, 6, 9, 10, and 12 (n = 25) were inoculated. The cells (passages 4–9 of the clones) used for inoculation were washed twice in PBS and 5 × 106 cells suspended in 100 µl PBS were inoculated subcutaneously in the flank of the mouse. If no tumor formation was observed after 1 month in any of the mice injected with the same clone, the group received a second injection in the other flank. Mice injected with the same clone lacking signs of tumor growth 1 month after the second injection received a final injection subcutaneously in the lumbar region with 1 × 107 cells in 100 µl PBS. The mice were euthanatized when the tumor reached a diameter close to 1 cm. The tumors were fixed in 4% phosphate buffered formaldehyde and embedded in paraffin. The sections were stained with hematoxylin and eosin (HE). At postmortem, tissue from the brain; all lung lobes, myocardium, liver, spleen, kidney, adrenal gland, uterus, ovary and submandibular lymph nodes; sternal lymph node; accessory axillary lymph nodes; popliteal lymph nodes; and the subiliac lymph nodes were fixed and examined for metastases. No metastases were seen in any of the mice inoculated with the various clones. The animal experiments were performed in accordance with protocols approved by the ethical committee, approval number C 135/3.
Results
Characterization of the spindle-cell tumor (309) and of tumors formed from cloned spindle-cell–tumor cells
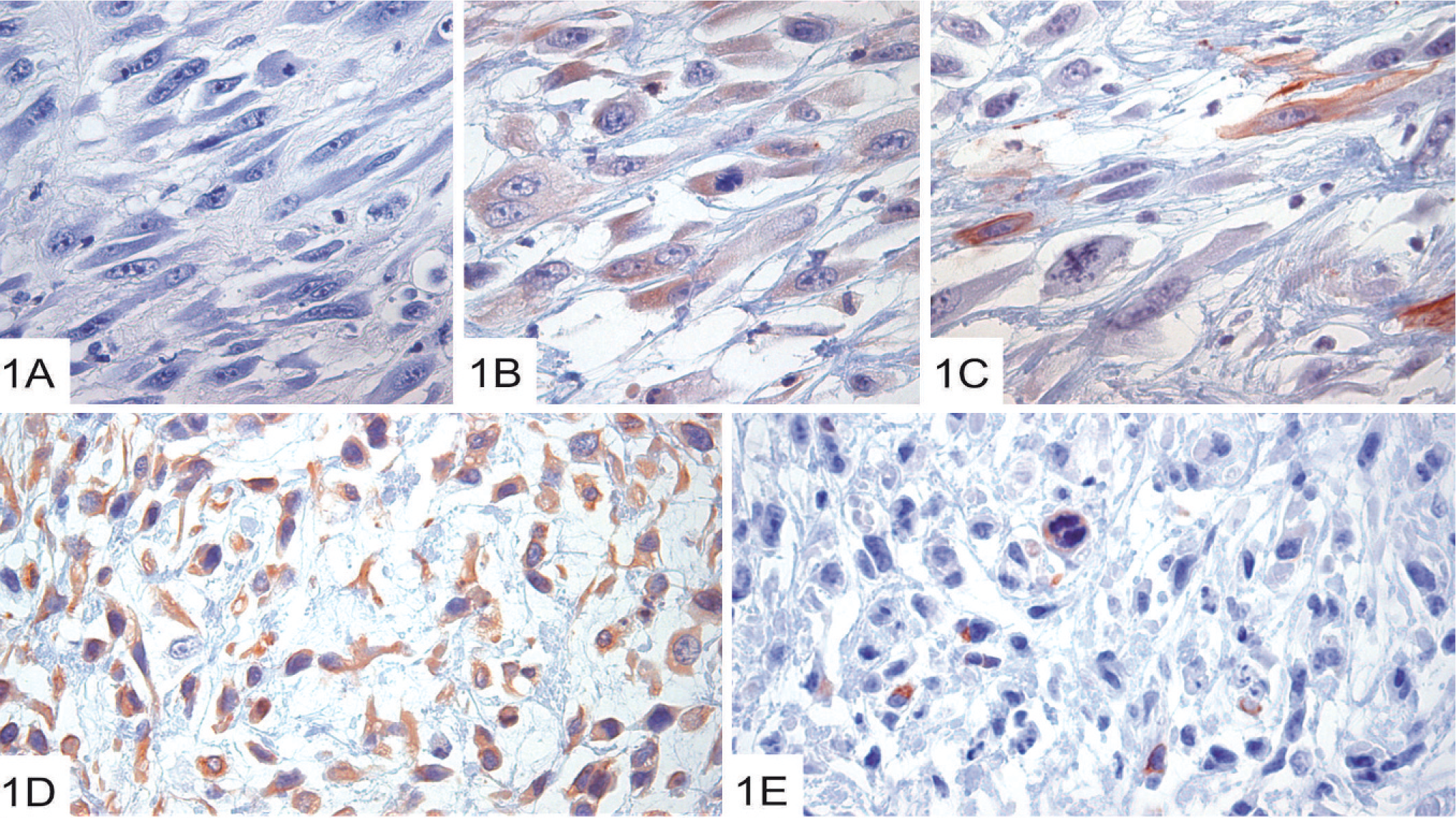
The primary spindle-cell–tumor cells had a uniform morphology, with elongated spindle-shaped cells in a fibrous stroma (Fig. 1). Tumor cells adjacent to central necroses were pleomorphic (Fig. 1A), whereas cells in other parts of the tumor were less pleomorphic (not shown). Vimentin expression was a common feature (Fig. 1D; Table 1), which indicates mesenchymal phenotype. However, keratin (an epithelial marker; Fig. 1B) and SMA (Fig. 1C; Table 1) expression was also seen. In parts of the tumor with less pleomorphic cells, ∼1% of the cells also strongly expressed neurofilaments (Fig. 1E; Table 1).
Immunohistochemical characterization of canine primary mammary tumors, of clones derived from the respective tumors and of tumors derived from inoculated clones.∗
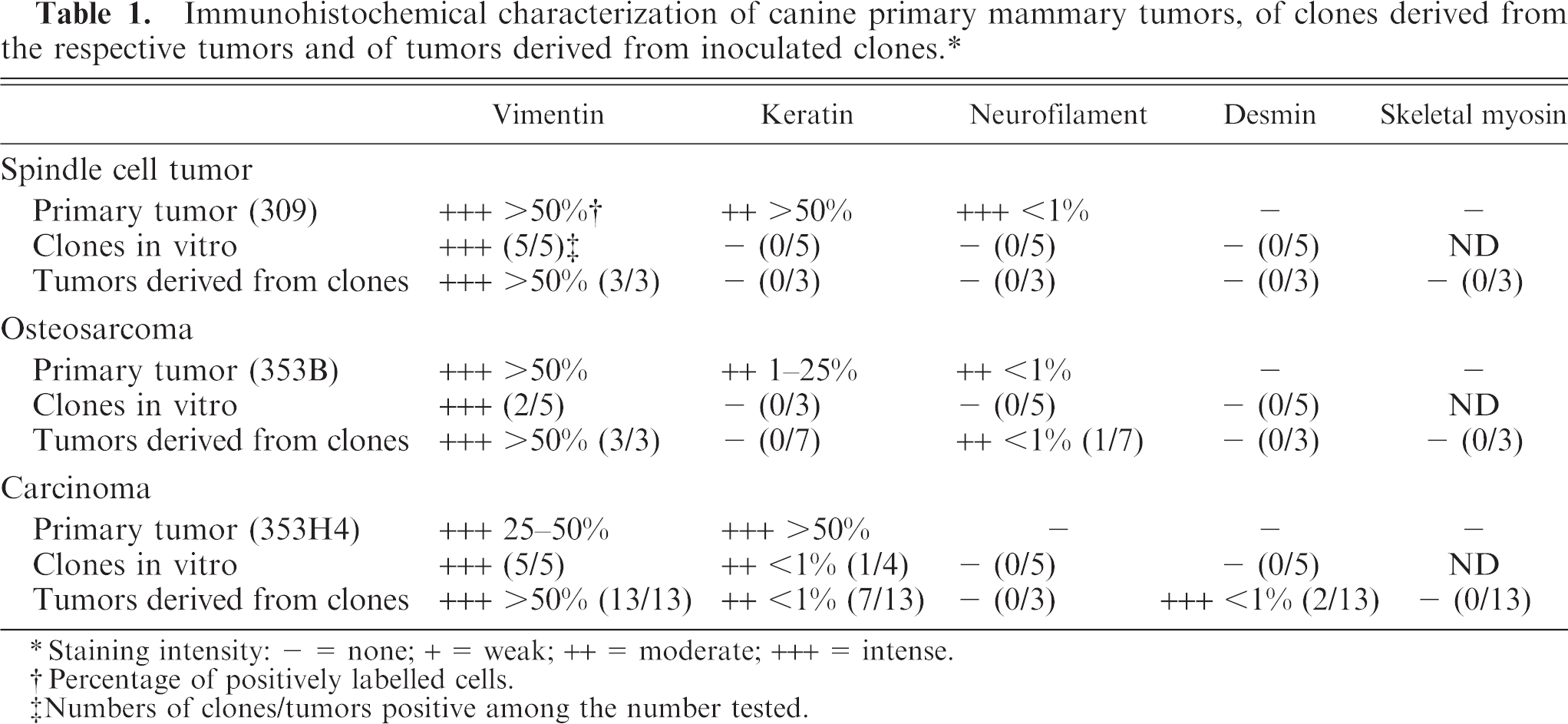
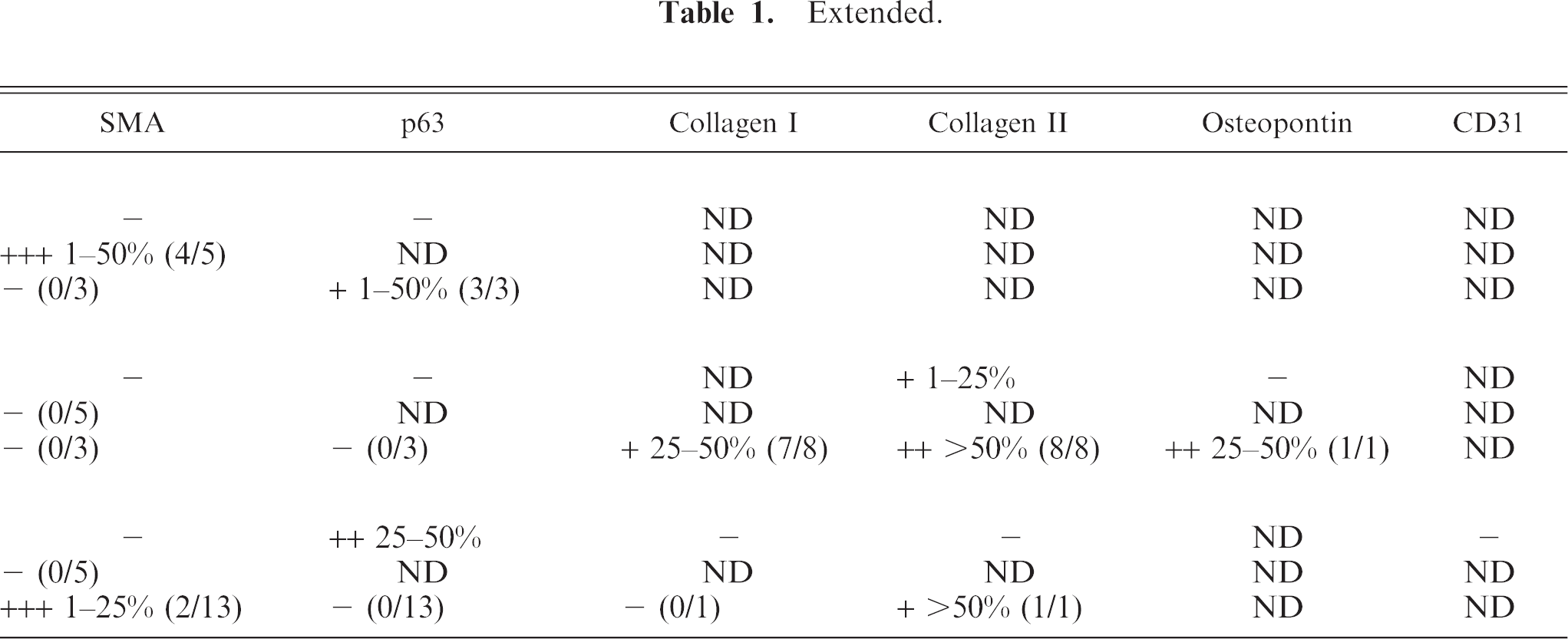
Staining intensity: - = none; + = weak; ++ = moderate; +++ = intense.
Percentage of positively labelled cells.
Numbers of clones/tumors positive among the number tested.
Characterization of the primary mammary spindle-cell tumor (309).
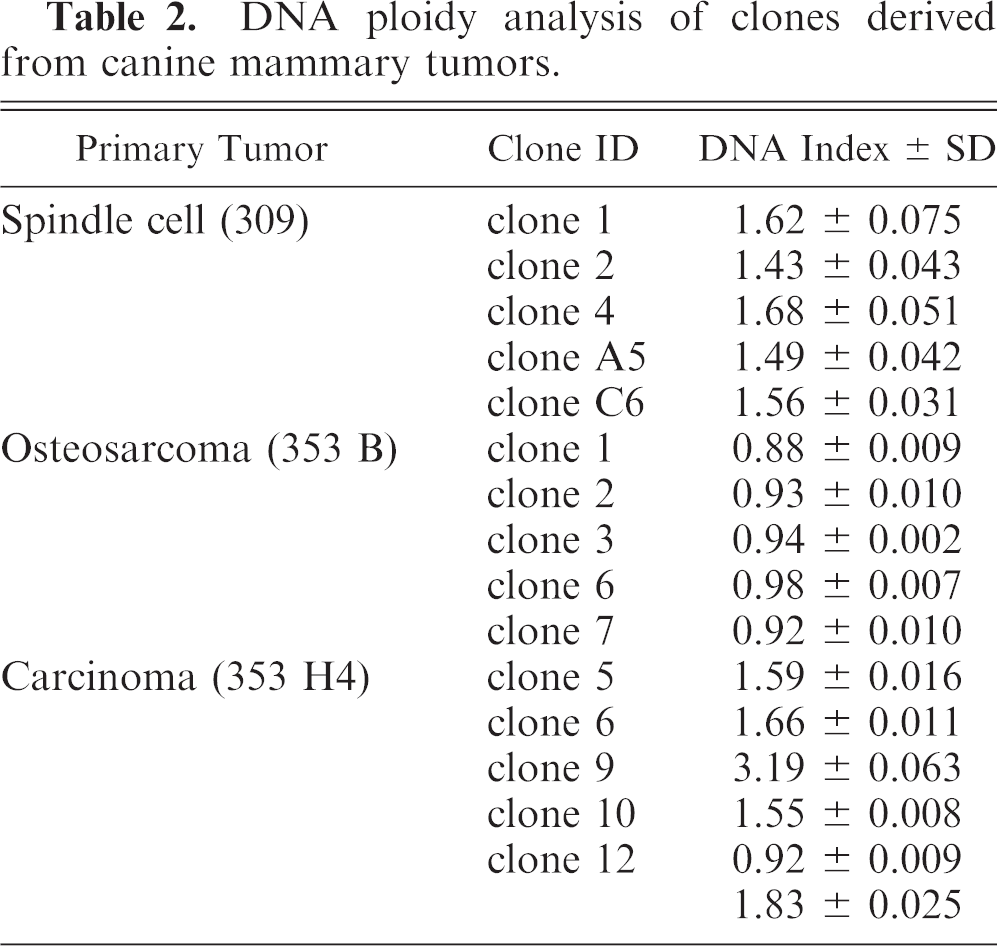
As described in Materials and Methods, clones were established from the spindle-cell tumor. Characterization of these showed that all of the clones were positive for vimentin (Table 1) and that 4 clones of 5 were partly positive (up to 50% of the cells) for SMA (Table 1). All of the clones were negative for expression of desmin (a muscle-cell marker), neurofilaments, pan-keratin, CD31 (an endothelial cell marker), and keratin 5, 6, 18 (Table 1). The 5 clones had all different DIs, although the variation was low (from 1.43 to 1.68; Table 2), and the histograms revealed single peaks for each of the clones, the latter indicated that the established cell lines indeed represent distinct clones.
DNA ploidy analysis of clones derived from canine mammary tumors.
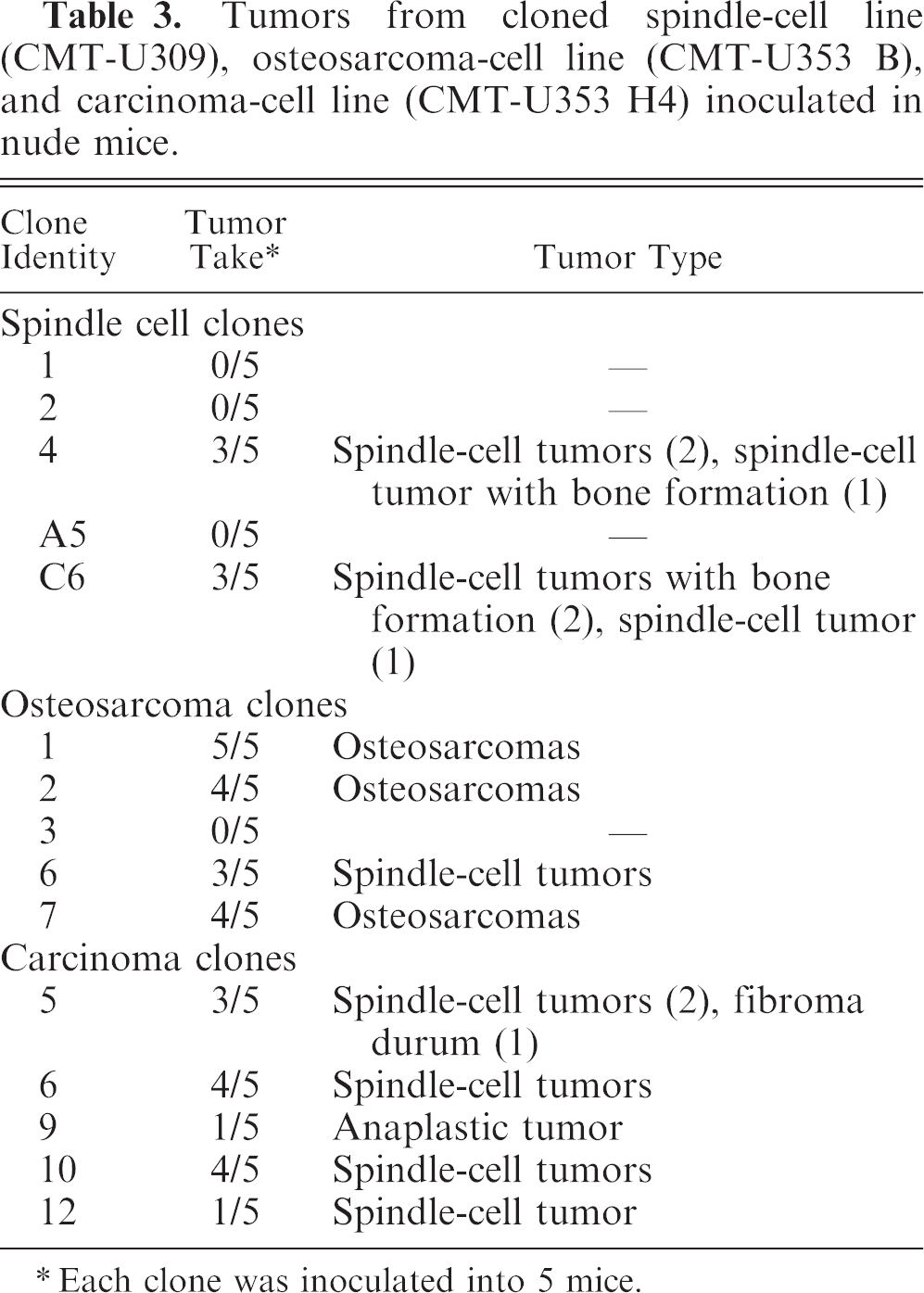
Each of the individual clones was inoculated into nude mice, followed by characterization of the generated tumor. The clones derived from the spindle-cell tumor gave rise to tumors that developed slowly, with only 2 of the 5 clones (clones 4 and C6) being able to induce tumors in nude mice (Table 3). In one of the tumors formed from clone 4, a focus of calcified bone was formed in a spindle-cell tumor (Fig. 3A), whereas spindle-cell tumors without bone formation were seen in the other tumors formed from clone 4 (Table 3; Fig. 3C). Also, in 2 of the tumors generated from clone C6, spindle-cell morphology and calcified bone was observed (Fig. 3B). Tumors formed from clones that did not give bone formation all displayed typical spindle-cell morphology (not shown). Immunohistochemical analysis revealed that all tested tumors generated from the spindle-cell clones expressed vimentin (Fig. 3B; Table 1). Furthermore, 25–50% of the cells in the tumor formed from clone 4, and 1–25% of the cells in the tumors generated from clone C6 also expressed the myoepithelial marker p63 (Fig. 3D; Table 1).
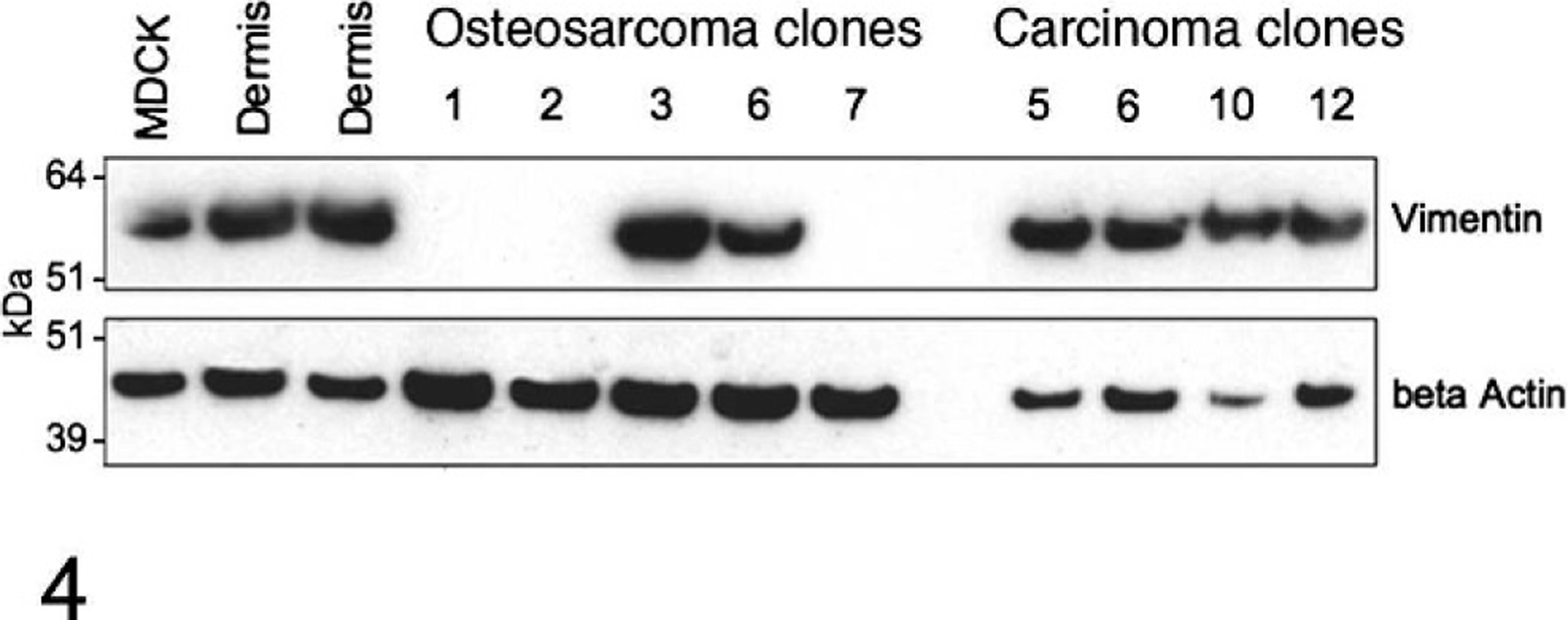
Tumors from cloned spindle-cell line (CMT-U309), osteosarcoma-cell line (CMT-U353 B), and carcinoma-cell line (CMT-U353 H4) inoculated in nude mice.
Each clone was inoculated into 5 mice.
Characterization of the primary combined mammary osteosarcoma (353 B).
Characterization of the primary osteosarcoma (353B) and of tumors formed from cloned osteosarcoma cells
The primary osteosarcoma was diagnosed as a poorly differentiated combined mammary osteosarcoma dominated by osteoblasts forming an osteoid matrix but also with areas with chondroid differentiation (Fig. 3A). A major portion of the tumor cells, both in the osteoid (not shown) and chondroid (Fig. 3B) areas, expressed vimentin (Table 1). Up to 25% of the tumor cells expressed keratins (Table 1), with the keratin-positive cells being evenly distributed both in osteoid (not shown) and chondroid (Fig. 3C) areas. Strikingly, strong expression of neurofilaments was seen in some tumor cells (Fig. 3D; Table 1). Further, 1–25% of the tumor cells expressed type II collagen, but the tumor was negative for desmin, skeletal myosin, SMA, p63, and type I collagen (Table 1).
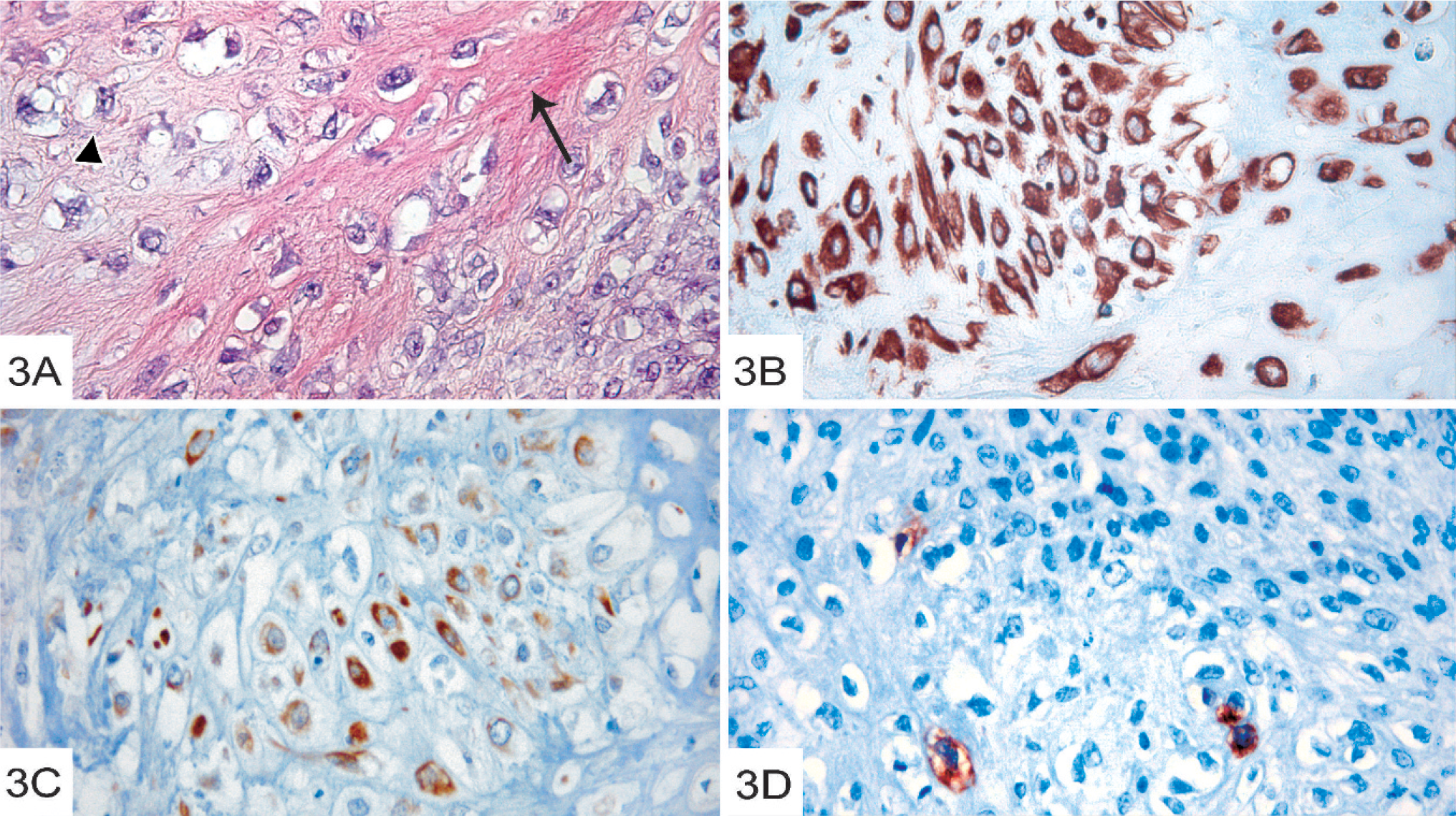
Individual clones were established from the primary osteosarcoma (see Materials and Methods). Characterization of these showed that 3 of the 5 established clones (clones 1, 2, and 7) were negative for a panel of lineage markers (vimentin, pan-keratin, keratin 5, 6, and 18; SMA; desmin and neurofilaments; Table 1). The lack of vimentin expression was confirmed by Western blot analysis (Fig. 4). In contrast, almost 100% of the cells in clones 3 and 6 strongly expressed vimentin but were negative for pan-keratin; keratin 5, 6, and 18; SMA; desmin; and neurofilaments (Table 1). The DIs of all osteosarcoma clones were below 1.00, indicating hypoploidy (Table 2), and the histograms revealed single, well-defined peaks, which indicated distinct clonal cell populations.
Western blot analysis demonstrating lack of vimentin expression in the osteosarcoma clones 1, 2, and 7, and presence in clones 3 and 6, as well as in the carcinoma clones. MDCK and canine dermal control cells showed expression of vimentin.
Clones from the osteosarcoma were inoculated into nude mice, followed by characterization of the generated tumors. All mice inoculated with clone 1 developed tumors, and clones 2 and 7 gave rise to tumors in 4 of 5 mice (Table 3), with tumors being formed more rapidly (within 1 month) than tumors generated after inoculation of the spindle-cell clones (7–9 months). All tumors from clones 1, 2, and 7 were poorly to highly differentiated osteosarcomas (Table 3). The poorly differentiated osteosarcomas formed from clones 1, 2, and 7 were composed by pleomorphic osteoblasts and little osteoid (not shown) whereas the highly differentiated osteosarcomas formed a calcified bone matrix (Fig. 5A).
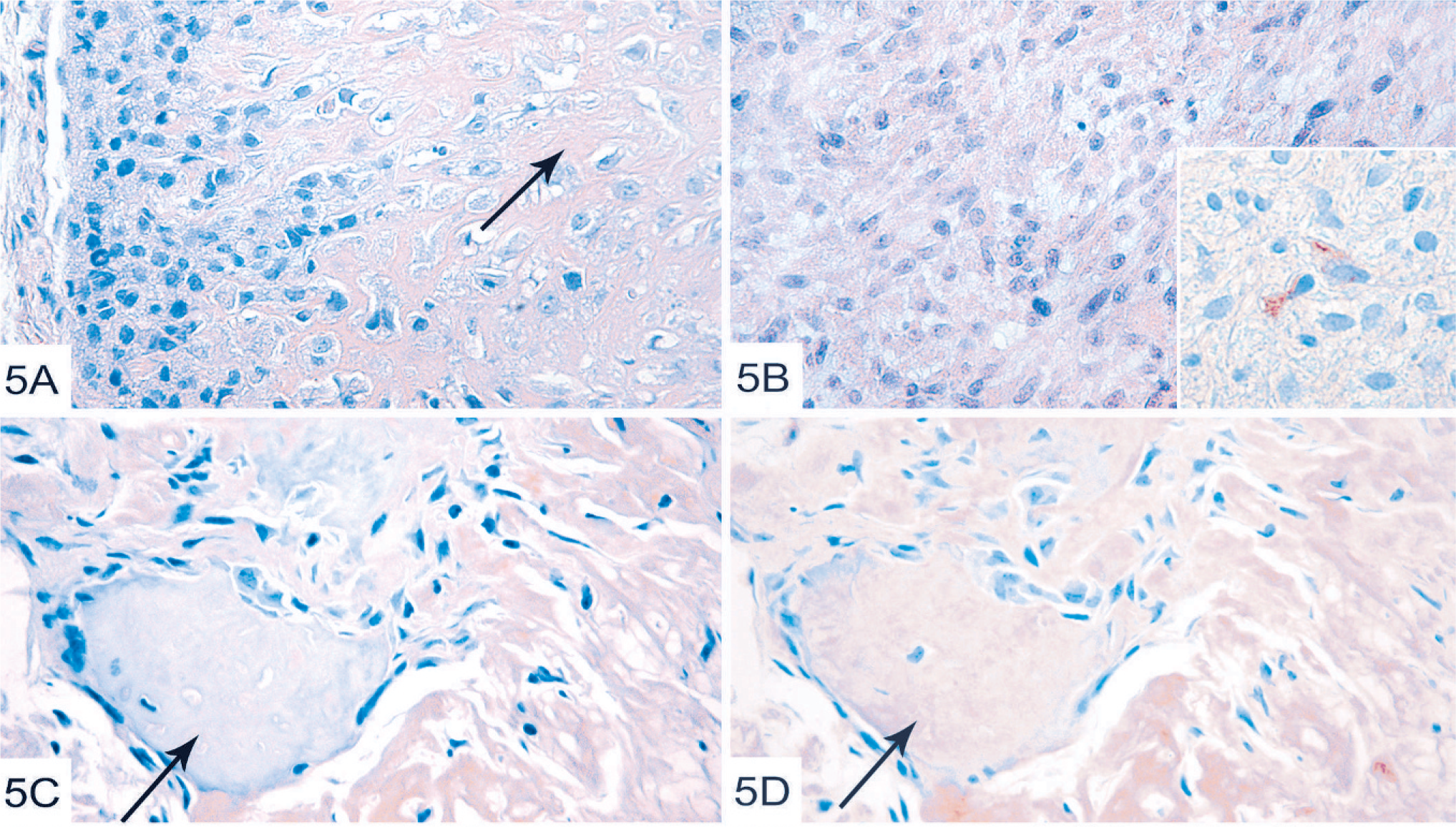
Tumors formed after inoculation of osteosarcoma clones in nude mice.
Three of the 5 mice inoculated with clone 6 developed tumors, and all of these exhibited spindle-cell characteristics (Fig. 5B; Table 3). These tumors developed slower (∼2 months) than tumors from clones 1, 2, and 7. Immunohistochemical analysis revealed strong expression of vimentin in near 100% of the tumor cells (Table 1). Interestingly, some cells in the tumor formed from one of the clones (clone 6) expressed neurofilaments (Fig. 5B, insert; Table 1), i.e., in contrast to the corresponding cell clones maintained in vitro but reflecting the expression of neurofilaments in the corresponding primary tumor (see Fig. 3D; Table 1). Furthermore, in contrast to the primary tumor, the spindle-cell tumor formed from clone 6 was negative for pan-keratin, SMA, desmin, and p63 (Table 1).
Because the morphologic examination of the primary osteosarcoma revealed both osteoid and chondroid areas (see Fig. 3A), we wanted to test if the tumors generated from the corresponding clones were positive for osteoid and chondroid markers, respectively. As a marker for bone, we used collagen I, whereas collagen II was used as a chondroid marker. As shown in Fig. 5C, the tumor formed from clone 1 was weakly positive for collagen I but, unexpectedly, only in noncalcified areas (Fig. 5C). In contrast, strong expression of collagen II, i.e., the chondroid marker, was seen both in calcified (arrow) and noncalcified areas. Tumors formed from clones 1 and 2 were negative for neurofilament expression and were also negative for pan-keratin (Table 1).
Characterization of the primary mammary carcinoma (353H4) and of tumors formed from cloned mammary carcinoma cells
The primary carcinoma was diagnosed as a simple scirrhous carcinoma, with highly pleomorphic tumor cells forming ductlike structures surrounded by a desmoplastic stroma (Fig. 6A). All neoplastic epithelial cells were strongly positive for the pan-keratin antibody, whereas the stroma as expected was negative (Fig. 6B). The neoplastic epithelial cells were also positive for vimentin, and also the stromal cells were positive for vimentin (Fig. 6C). Further, 25–50% of the neoplastic epithelial cells were positive for the myoepithelial cell marker p63 (Fig. 6D). Some stromal cells (25–50%) showed strong positivity for SMA and none of the tumor cells (epithelial or stromal) expressed CD31, an endothelial cell marker (Table 1).
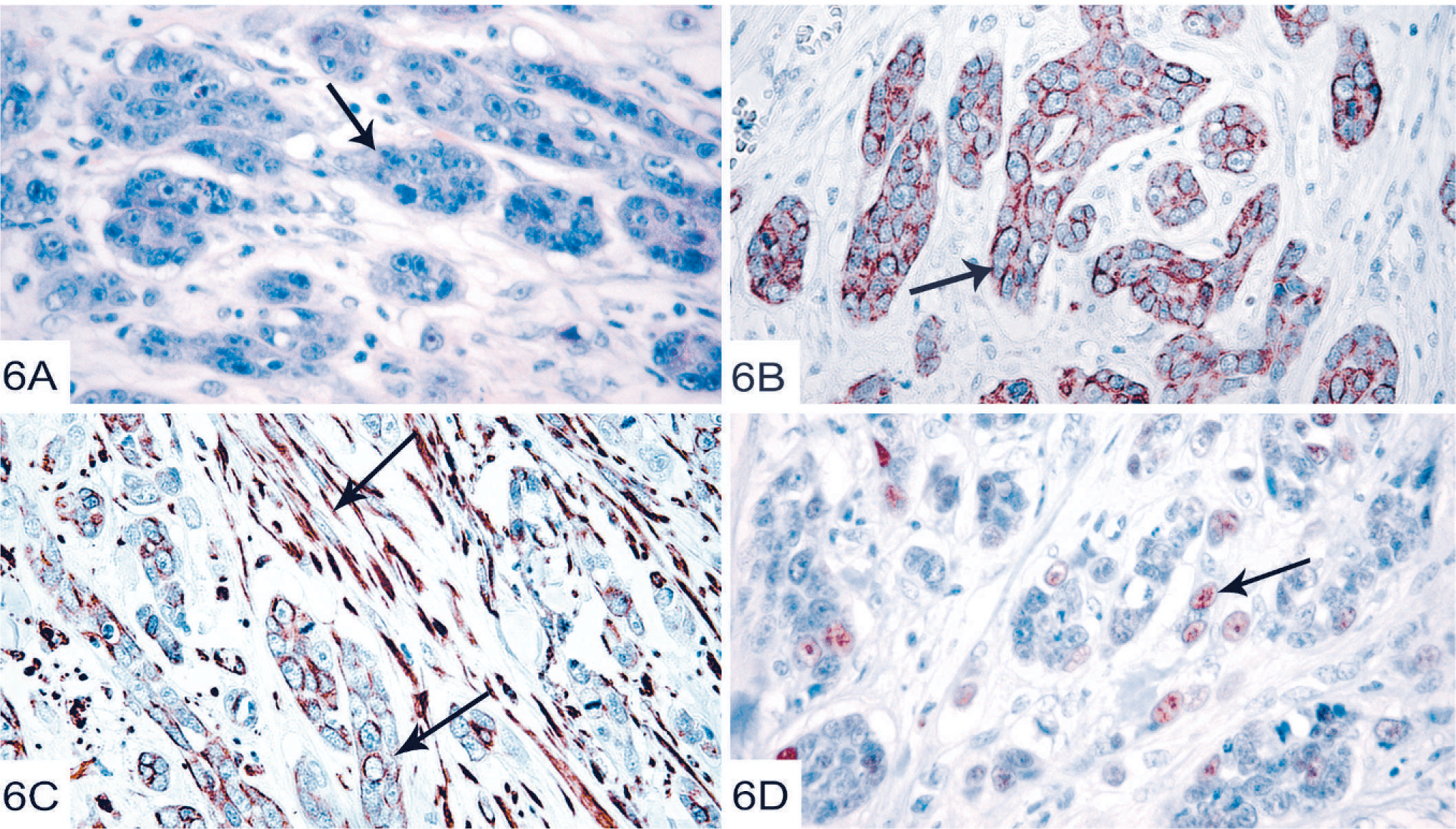
Characterization of the primary mammary carcinoma (353 H4).
As described in Materials and Methods, individual clones were generated from the primary scirrhous carcinoma, and were characterized. All characterized clones (13 clones) were positive for vimentin (Table 1), in agreement with the strong vimentin expression seen in the primary tumor (see Fig. 6C). However, in contrast to the strong expression of keratin in the primary tumor (see Fig. 6B), most clones were negative for the keratin, with only one of the characterized clones (clone 6) being positive (Table 1). Furthermore, all clones were negative for SMA, desmin, and neurofilaments (Table 1). The DIs varied from 1.55 to 3.19 (Table 2), which indicated substantial chromosomal rearrangements, the single peaks again indicated distinct clonal-cell populations with one exception; in clone 12, one peak at DI 0.92 could be seen, with an additional aneuploid peak at DI 1.83 (Table 2).
The experimental tumors generated from the carcinoma clones grew slowly, taking 4 to 5.5 months to form, except for one of the tumors generated from clone 6, which developed within 2 months. All of the clones generated tumors in nude mice, although clones 5, 6, and 10 appeared to be more efficient (Table 3). The tumor generated from clone 9 was anaplastic and composed of pleomorphic cells in a myxoid stroma. The tumor formed from clone 5 was a small fibroma durum. All remaining tumors formed from the carcinoma clones had spindle-cell–like morphology (Table 3). No presence of ductular growth was seen in any of the tumors (Fig. 7A).
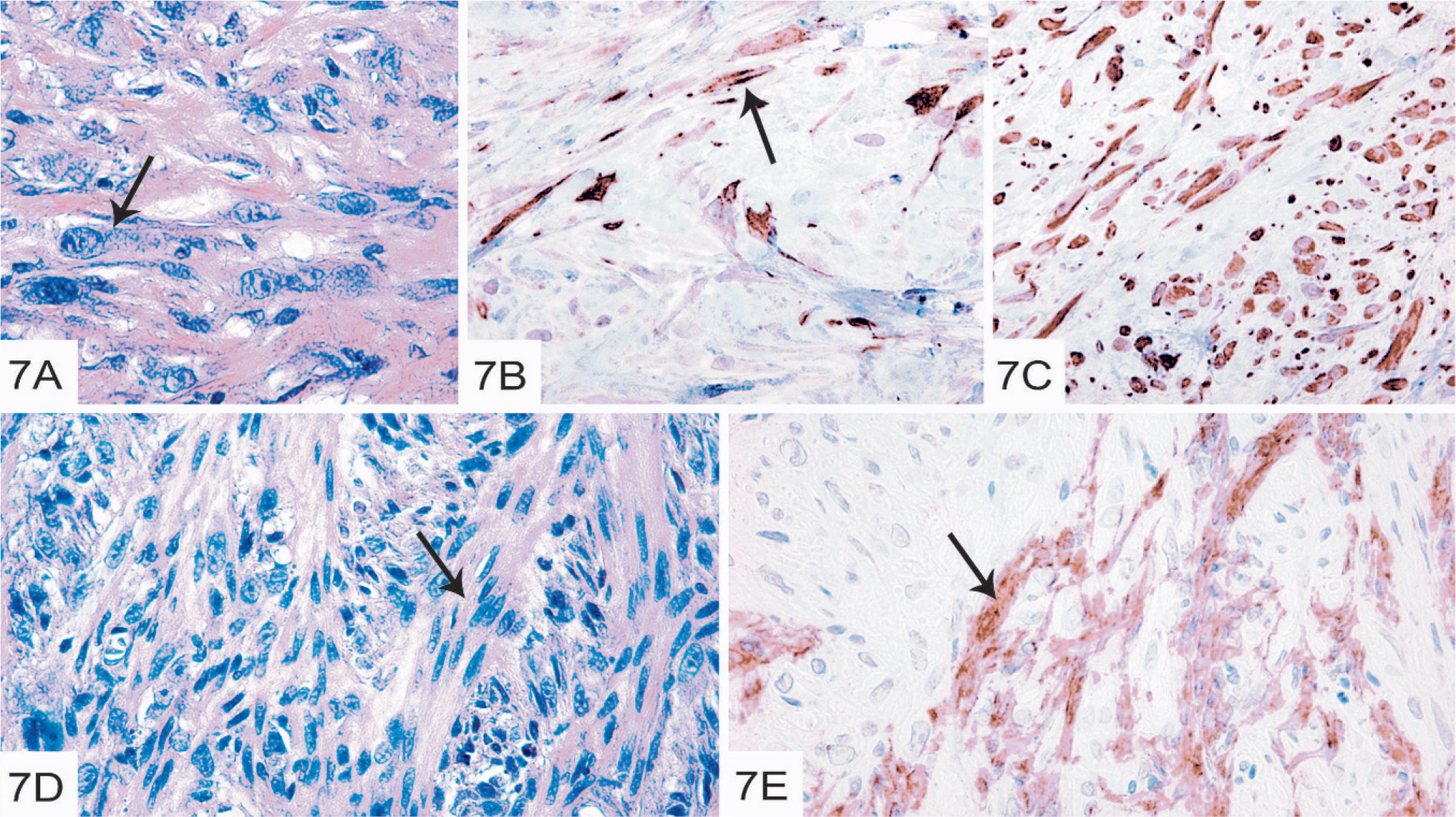
Tumors formed after inoculation of carcinoma clones in nude mice.
Immunohistochemical analysis showed that most of the tumors (7 of 13) were positive for keratin (Fig. 7B), although most of the used clones were negative for keratin in vitro (Table 1). However, keratin expression was only seen in a limited number of the tumor cells (up to 1%). In all of the tumors formed by the carcinoma clones, the majority of the tumor cells were positive for vimentin (Fig. 7C; Table 1), in agreement with the strong expression of vimentin in the isolated carcinoma clones. In 2 of the tumors (from clone 10), SMA and desmin expression was seen in one distinct part of the tumor, whereas other parts were negative (Fig. 7E; Table 1). In the same tumor, HE staining revealed bundles of smooth-muscle–like cells, interspersed by pleomorphic tumor cells (Fig. 7D). Thus, the SMA and desmin positivity of this tumor is in agreement with the smooth-muscle-cell–like morphology. In contrast to the primary tumor, none of the tumors generated from the carcinoma clones showed expression of p63 (Table 1).
Discussion
This study was undertaken to investigate if individual, cloned mammary tumor cells were capable of generating tumors of different phenotype in vivo, i.e., displaying phenotypic plasticity. Indeed, we herein presented multiple lines of evidence that supported such a notion. One important example is our demonstration of bone formation in tumors derived from cloned spindle-cell–tumor cells (Fig. 2A and B). To the best of our knowledge, this has not been demonstrated previously in nonosteosarcoma spindle-cell tumors. Another example along the same line is the formation of bone in tumors formed from osteosarcoma clones 1, 2, and 7. These clones were strikingly negative for a panel of lineage markers in vitro, including keratin and vimentin. Such an absence of both keratin and vimentin has previously only been shown in preimplantation embryos and in blastocysts. 15, 25 Yet, they generated tumors with calcified bone when inoculated into nude mice, and, in addition to vimentin, the tumors were positive for both collagen I and collagen II. Another example of phenotypic change is constituted by the clone from the osteosarcoma that expressed only vimentin in vitro but was positive for neurofilaments, as well as vimentin when inoculated in mice. It is thus clear that these clones have undergone dramatic changes in their intermediate filament expression profile during the course of tumor formation, i.e., after inoculation of the individual clones. Unexpectedly, collagen II, a chondroid marker, was found in both calcified and noncalcified areas, and collagen I, usually regarded as a marker for bone, was absent in highly differentiated, calcified bone areas of tumors generated from osteosarcoma clones. In many cases, collagen I and II showed coexpression. These findings are also in line with previous reports that showed coexpression of collagen I and II, for example, during chondroid differentiation, and that the collagen expression profile may undergo changes during the course of differentiation. 11, 14, 26, 28, 34
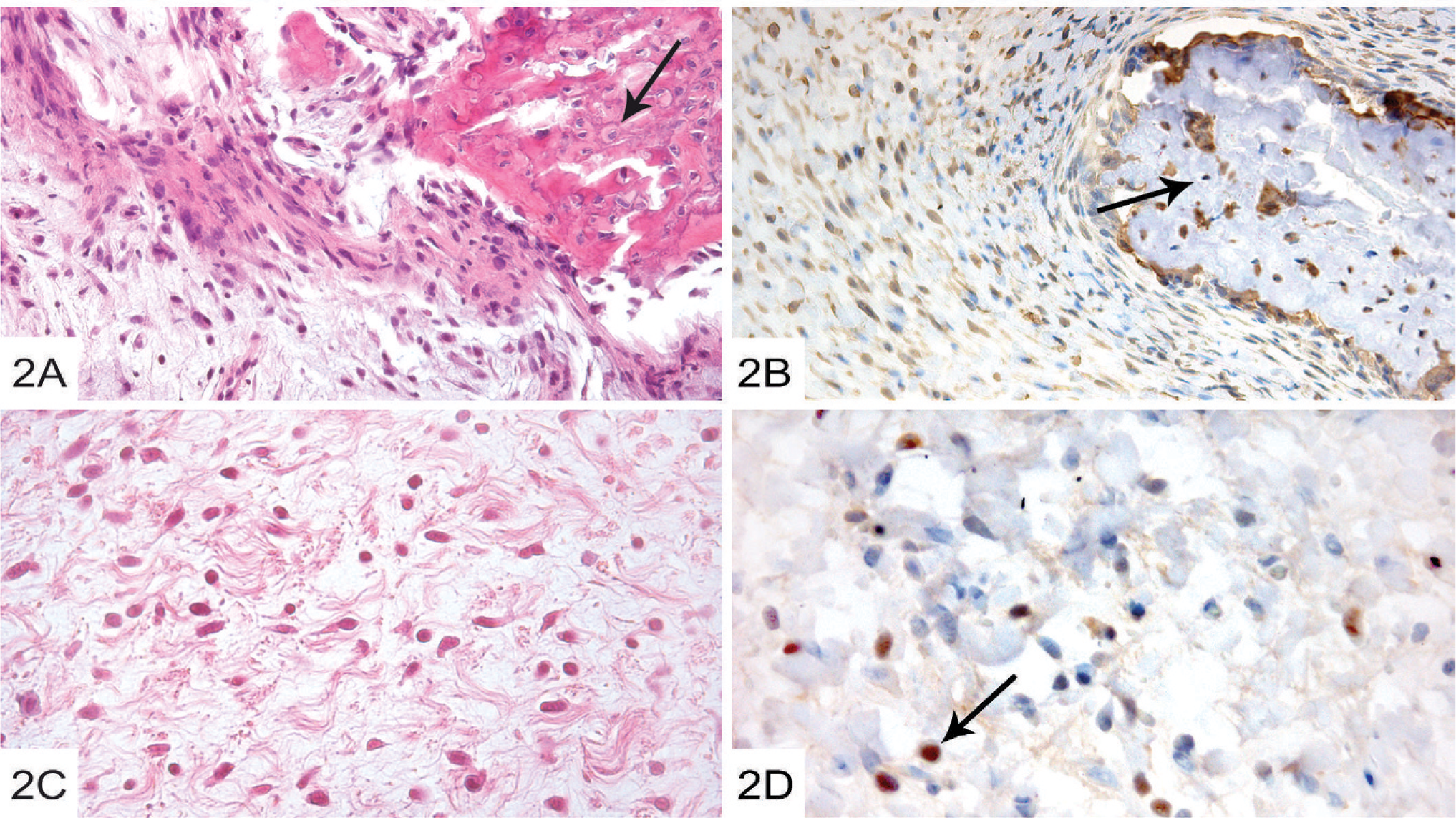
Tumors formed after inoculation of spindle-cell clones in nude mice.
Neurofilament expression was seen in the primary osteosarcoma (Fig. 3D), whereas the corresponding clones were negative in cell culture. However, the neurofilament expression was regained after inoculation of one of the clones into nude mice, thus providing yet another example of phenotypic changes during tumor formation. The expression of neurofilaments in the canine mammary osteosarcoma is also in line with previous reports that show expression of neurofilaments in different canine mammary tumors, 12 and there are also examples of neurofilament expression in human breast carcinoma. 10 Based on morphologic criteria, the neurofilaments were expressed by tumor cells, i.e., the most likely explanation is that the neurofilament expressing cells were of canine origin. However, we cannot completely rule out the possibility that the stained cells represent host neural structures infiltrating the mass. Somewhat unexpectedly, keratin expression was seen in the canine primary osteosarcoma, although similar findings were previously reported. 12 The occurrence of chondroid foci expressing keratins has also been reported for human metaplastic carcinoma of the breast. 4, 17, 19 In human pathology, a breast tumor of pure sarcomatoid morphology that expresses keratin is by definition a metaplastic carcinoma. However, there are studies that indicate that sarcomatoid metaplastic carcinomas have a biology similar to sarcomas. 5, 9 The keratin expression in the osteosarcoma in this study was lost during the cloning procedure, and no keratin expression was seen in any of the tumors generated from the inoculated osteosarcoma clones.
Another interesting finding was that the primary carcinoma was positive for vimentin, a mesenchymal marker. However, vimentin expression was previously reported in the basal cells of a canine mammary carcinoma, and, interestingly, vimentin expression in human breast carcinoma has been correlated with invasiveness and resistance to chemotherapy. 18 As expected, the primary carcinoma was positive for keratin expression, but, unexpectedly, most of the derived carcinoma clones were negative for keratin. However, a majority of the tumors generated from the carcinoma clones regained their expression of keratin after inoculation in nude mice and formation of tumors in vivo. In addition, whereas all of the carcinoma clones were negative for myoepithelial markers, SMA and desmin, occasional tumors generated after inoculation of these clones into nude mice were positive for SMA and desmin, and even showed smooth-muscle–like morphology. Thus, these findings provide other examples of phenotypic changes during the process of tumor generation.
P63, another myoepithelial marker, has recently been suggested to be a marker for canine mammary tumors, with a recent study that showed that a majority of neoplastic canine mammary tissues were p63 positive, which indicated myoepithelial origin. 8 In our study, p63 expression was indeed seen in the primary carcinoma, thus supporting a myoepithelial origin of the tumor. However, p63 expression was not seen in the primary osteosarcoma or in the spindle-cell tumor. Furthermore, we were unable to detect p63 protein expression in the experimental tumors with the exception of the tumors derived from inoculated spindle-cell–tumor clones. Thus, we were not able to confirm in our experimental system that p63 can be used a general marker for canine mammary tumors.
Our findings may have a bearing on 2 central issues of tumor genesis: epithelial to mesenchymal transition (EMT) 1 and involvement of cancer stem cells. 30 EMT may, for example, explain the lack of keratin expression in clones derived from the mammary carcinoma. However, several of our findings are more compatible with stem-cell involvement. First, the formation of bone in tumors derived from spindle-cell clones indicates that these cells have a differentiation potential, and this, in turn, could be explained by stem-cell phenotype of the bone-generating spindle-cell–tumor clones. Second, the immature appearance of 3 of the osteosarcoma clones in vitro, but formation of highly differentiated bone tissue after their inoculation into nude mice can be explained by stem-cell characteristics. Also the expression of neurofilaments in a tumor derived from an osteosarcoma clone that was neurofilament negative in vitro can be explained by stem-cell characteristics. In a recent study, the stem-cell marker Oct4 was identified in cells from a number of canine neoplasms, including a mammary adenocarcinoma, thus possibly identifying a cancer stem-cell population in the tumors. 33 Although the significance of these findings remains to be proven, it implicates stem cells in tumorigenesis. Another possible explanation to our results could be clonal heterogeneity or clonal evolution. According to such a model, the acquirement of mutations during cell culture could generate heterogeneity in our experimental tumors. There is substantial evidence for both clonal evolution and the cancer-stem-cell model and, importantly, there are indications that the models are not necessarily mutually exclusive (reviewed by Campbell and Polyak 3 ). However, in our experimental setup, we favor the possibility that plasticity has a more important role than clonal evolution. For example, to minimize the influence of in vitro clonal evolution, we consistently used cells at low passage numbers. Furthermore, bone formation is the result of complicated genetic programs and less likely to be the result of mutations caused by clonal evolution. It is also important to note that 5 different clones (3 osteosarcoma clones and 2 spindle-cell clones) formed bone in the experimental tumors, and it is unlikely that all of these clones had undergone similar clonal evolution processes, i.e., which resulted in induction of bone-generating programs. Plasticity is also supported by the findings that several osteosarcoma clones have a downregulated expression of vimentin and probably the bone-forming genetic program in cell culture, as also shown in the mammary osteosarcoma cell-line MCO-Y4, 16 but regained vimentin expression and formed bone when inoculated in mice. In conclusion, the fact that genetic programs can be turned on/off in a controlled way and that multiple clones performed in the same way favor plasticity before in vitro clonal evolution.
Footnotes
Acknowledgements
This work was financed by a Strategic funding from the Swedish University of Agricultural Sciences (SLU), Uppsala, Sweden (to EH). We thank Karin Agehall, Ulrika Larsson, and Ewa Westergren for skillful technical assistance. We also thank Nicolette Hillbertz and Kumar Juneja for control samples, Juana Martin de las Mulas and Wim Misdorp for interesting discussions on histopathology, Ivana Juko for Western blot analyses, and Nils-Erik Heldin for helpful discussions and comments on the manuscript.