
Abstract
The respiratory epithelium is a primary site for the deposition of microorganisms that are acquired during inspiration. The innate immune system of the respiratory tract eliminates many of these potentially harmful agents preventing their colonization. Collectins and cationic antimicrobial peptides are antimicrobial components of the pulmonary innate immune system produced by respiratory epithelia, which have integral roles in host defense and inflammation in the lung. Synthesis and secretion of these molecules are regulated by the developmental stage, hormones, as well as many growth and immunoregulatory factors. The purpose of this review is to discuss antimicrobial innate immune elements within the respiratory tract of healthy and pneumonic lung with emphasis on hydrophilic surfactant proteins and β-defensins.
Introduction
The respiratory epithelium is a mucosal surface that routinely comes in contact with environmental substances including particulate matter, vapors, aerosols, and microbial pathogens. 32 The innate immune components of the respiratory tract eliminate many of these potentially harmful substances. These components include coughing, the mucociliary escalator, phagocytosis by leukocytes such as pulmonary alveolar macrophages and pulmonary intravascular macrophages, 3, 91 the complement cascade, 66 interferons, 50, 106 neurokinins, 11, 112 and antioxidants. 91 More recently, additional components of the mucosal secretions such as defensin peptides, anionic peptide (AP), and surfactant proteins are increasingly becoming appreciated for their role in preventing microbial colonization/infection and also for triggering the adaptive immune response. 17, 146
Cellular Components of the Respiratory Epithelium
The upper respiratory tract (nasal cavity, sinuses, and trachea) and larger airways of the lower respiratory tract are lined by pseudostratified, ciliated epithelium and submucosal glands (Fig. 1). 32 Other epithelial cells include mucus-secreting goblet cells, small mucous granule cells, and brush cells. 7 The number of basal cells decreases with the decreased diameter of airway branches, and the epithelium becomes simple cuboidal. At the bronchiolo-alveolar junction, nonciliated (Clara) cells are abundant. The alveolar epithelium consists of a continuous layer of 2 principal cell types, thin, elongate type I pneumocytes (ATI cells), which cover 95% of the alveolar surface, and cuboidal to rounded type II pneumocytes (ATII cells). 80 In addition, there are also mucous and serous epithelial cells of the tracheal and bronchial submucosal glands, which together with the pseudostratified epithelium and alveolar epithelial cells serve as an important physical barrier, and also produce and modify airway surface liquid (ASL). 131
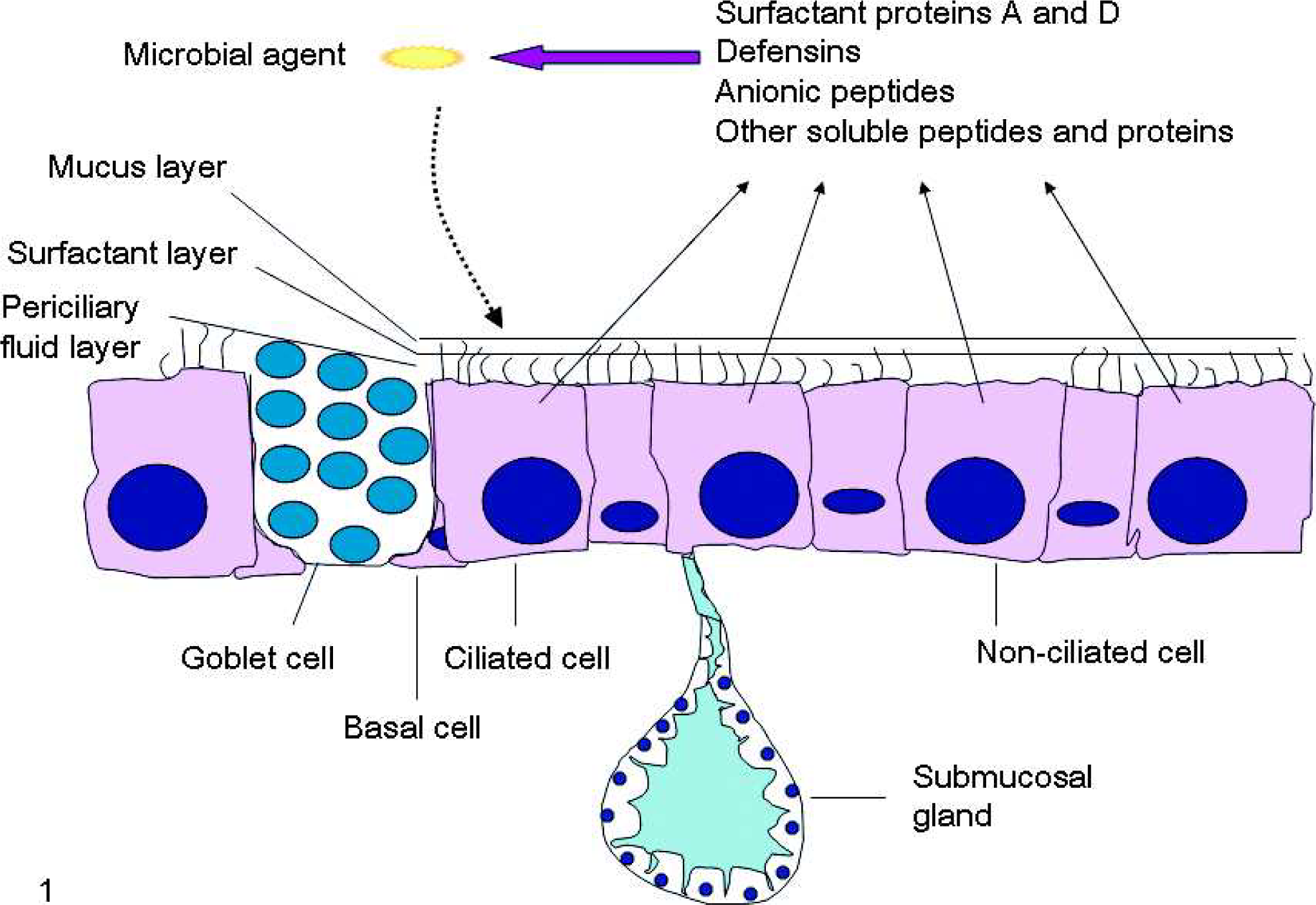
Schematic presentation of the respiratory epithelium and its production of antimicrobial peptides and proteins.
Function of the Respiratory Epithelium
Ciliated epithelial cells participate in mucociliary clearance, form a physical barrier, and secrete a broad spectrum of antimicrobial factors. 32, 131 Clara cells are the major progenitors of ciliated cells after bronchiolar injury, 28, 64 and are involved in the detoxification of xenobiotic compounds through cytochrome P-450 monooxygenase system activity. 91 Clara cells produce surfactant proteins and Clara cell secretory 10-kd protein (CCSP/CC10), which are natural regulators of airway inflammation and cytodifferentiation markers. 28, 107 ATII cells are responsible for pulmonary surfactant synthesis, storage in lamellar bodies, secretion, and recycling. 80, 124 They also act as stem cells by repopulating the lung with ATI and ATII cells during normal tissue turnover and during the repair of alveolar epithelium after injury. Recently, it has been shown that the dendritic cell-lysosomal associated membrane protein (DC-LAMP/CD208) is constitutively expressed by mouse, sheep, and human ATII cells and can be used as a marker for normal as well as transformed (adenocarcinoma) ATII cells. 124 ATII cells also express cathepsins, major histocompatibility complex class II molecules, intercellular adhesion molecule-1 (CD54), and costimulatory molecules such as CD80 and CD86. 124 For ATI (also known as membranous pneumocytes), one of the most useful differentiation markers is T1α because it is not expressed by ATII cells or endothelial cells. 145 The main function of ATI cells is to facilitate gaseous exchange. There is limited data on ATI cells' metabolic and immune functions; however, these cells may play a role in the maintenance of normal alveolar homeostasis and protection from injury, lung development and remodeling, host defense, tumor/growth suppression, surfactant metabolism, and other activities. 30
Airway Surface Liquid
The airway surface liquid represents a first line of defense against inhaled pathogens. 94 It consists of the superficial gel or mucus layer and the layer of periciliary fluid, which are separated by a thin layer of surfactant (Fig. 1). 119 The mucus layer is 93–95% water and 5–7% solid material principally composed of mucins (large glycoproteins) admixed with phospholipids, proteoglycans, cellular debris, and various proteins some of which possess antimicrobial, antiprotease, and antioxidant activity. 85 The mucus traps inhaled particles and microbial pathogens, neutralizes soluble gases, and is cleared by an interaction between airflow and cilia activity. 91 The periciliary fluid layer is a watery layer that surrounds the cilia on the apical surface of the ciliated cells. 94 This layer is necessary for efficient ciliary beating and mucin hydration. Surfactant is 80% phospholipids (phosphatidylcholine being the most abundant), 12% neutral lipids (cholesterol being the most abundant), and 8% surfactant proteins (SP) including hydrophilic SP-A and SP-D, and hydrophobic SP-B and SP-C. 108 The primary function of surfactant is to regulate the surface tension at the air–liquid interface, preventing alveolar collapse at low lung volumes and overinflation at high lung volumes. Surfactant also facilitates effective spreading of mucus, preservation of gel and periciliary fluid integrity, and beating of cilia without entanglement.
Antimicrobial Factors of Airway Surface Liquid
Airway surface liquid contains several proteins and peptides with antimicrobial activity (Table 1). 22, 32, 34, 75, 123, 125, 126, 131, 143, 144 Most of these factors act cooperatively in their microbicidal activity and the degree of their activity depends on the ionic strength of the solution. 131 With increased ionic strength of the ASL, the antimicrobial activity of ASL components decreases dramatically.
Antimicrobial factors or substances in airway surface liquid.

∗ Note: These proteins have highest production levels in salivary glands. SP-A = surfactant protein A; SP-D = surfactant protein D.
Lysozyme and lactoferrin are the most abundant proteins in ASL and are present at 0.1–1 mg/ml concentration. 43 Lysozyme was first recognized by Alexander Fleming in 1922. It is a 14-kd enzyme produced by neutrophils, monocytes, macrophages, and epithelial cells. 43, 131 It enzymatically cleaves glycosidic bonds of the bacterial membrane peptidoglycans or kills bacteria by a nonenzymatic mechanism. Lactoferrin is an 80-kd cationic iron-binding protein and is also produced by neutrophils and epithelial cells. This protein inhibits growth of iron-requiring bacteria and can also be directly microbicidal through its N-terminal cationic fragment.
Antiproteinases are also important constituents of ASL that are involved in the acute phase of inflammation. 104, 125 Their molecular characteristics include low-molecular weight, positive charge, and numerous disulfide bonds. They are present at the mucosal surfaces and protect against noxious effects of proteolytic enzymes released by phagocytic cells thereby controlling excessive inflammation and septic shock. An important member of antiproteinases is secretory leukoprotease inhibitor (SLPI), a low-molecular-weight cationic proteinase inhibitor produced by epithelial cells and macrophages. This factor acts as an effective inhibitor of neutrophil elastase via the C-terminal domain. Secretory type II phospholipase A2 (PLA2) is another antiproteinase that belongs to a growing family of enzymes produced by epithelia and blood-derived cells. Such enzymes are involved in cytokine-mediated inflammation, surfactant degradation, and bactericidal activity especially against Gram-positive bacteria. 5, 47, 88 Other antiproteinases include locally synthesized and secreted elastase-specific inhibitor (elafin), and systemic antiproteinases produced in the liver and secreted in the circulation such as α1-antichymotrypsin and α1-proteinase inhibitor.
Antimicrobial peptides and proteins from the ASL that have received the least attention are those produced by respiratory epithelia, including CC10, APs, cationic antimicrobial peptides (AMP), and collectins. CC10 protein is induced by corticosteroids and is a potent anti-inflammatory cytokine produced by Clara cells. Although tumor necrosis factor-α (TNF-α) released during lung inflammation increases the expression of both CC10 and PLA2, CC10 inhibits the activity of PLA2. 71, 88, 152 Additional features of CC10 include a dose-dependent suppressive effect on TH2 cytokine expression and a potential protective role in lung tumorigenesis. 71, 150
Anionic peptides are small, hydrophilic peptides that contain homopolymeric regions (e.g., 5–7 residues) of aspartic acid, which are responsible for their negative charge. 16, 41 They have been detected in the lungs, bronchoalveolar lavage (BAL) fluid, and pulmonary surfactant of humans, sheep, and cattle. For maximal bactericidal activity, APs require zinc as a cofactor, which may allow the peptide to overcome the net negative charge once on the microbial surface. 17
Collectins
Collectins produced by vertebrates are a family of proteins that include mannan-binding lectin (MBL/mannose-binding protein), collectin liver 1 (CL-L1), collectin placenta 1 (CL-P1), SP-A, SP-D, conglutinin, 43-kd collectin (CL-43), and 46-kd collectin (CL-46), the last three being found only in the bovidae.
57, 62, 68
These proteins share a common structure made of a C-terminal located C-type
Lung Collectins
Functional Roles
A large body of evidence has been collected for at least four potential functional roles of lung surfactant proteins based on in vitro and in vivo studies. 29 These roles include antimicrobial, anti-inflammatory, immunomodulatory, and surfactant regulatory activities.
Lung collectins bind, aggregate, and opsonize different microorganisms including Gram-positive and Gram-negative bacteria, enveloped (influenza A virus [IAV], respiratory syncytial virus [RSV]) and nonenveloped viruses (Rotavirus), and fungi to enhance phagocytosis, killing, and clearance of microorganisms from the lung (Table 2). 68 Initially, SP-A and SP-D have an antimicrobial role that is often followed by a proinflammatory response to destroy the pathogen and prevent further spread. In vitro, SP-D enhances phagocytosis of RSV by alveolar macrophages and neutrophils, and suppresses the inhibitory effect of RSV on oxygen radical production by these cells. 82 SP-D also inhibits the infectivity and hemagglutination activity of IAV in vitro by blocking pathogen-target cell attachment sites. 29 Next, SP-D binds, agglutinates, and/or kills pathogens, resulting in enhanced physical or cellular clearance. 25 SP-D agglutinates Mycobacterium tuberculosis but inhibits its internalization; 29 a more recent study shows that both SP-A and SP-D may enhance mannose receptor-mediated phagocytosis of Mycobacterium avium by macrophages. 79 SP-A can also cause direct killing of microorganisms by inducing the formation of reactive oxygen species (ROS). 53 In addition, both SP-A and SP-D can inhibit the proliferation of pathogens such as Gram-negative bacteria by increasing the permeability and disrupting the microbial cell membrane. 147
Interaction of surfactant protein A(SP-A) and surfactant protein D (SP-D) with pathogens.

According to in vivo studies, the antimicrobial role of SP-A and SP-D appears to depend on the type of pathogen and may be species-specific. In wild-type mice, SP-D levels increase markedly after IAV infection, 117 whereas decreased viral clearance is observed after challenge of SP-A knockout (SP-A -/-) mice with RSV 96 or SP-D knockout (SP-D -/-) mice with RSV and IAV. 29, 82 In neonatal lambs infected intratracheally with parainfluenza type 3 (PI-3) virus, SP- A and SP-D mRNA levels are significantly elevated in the lung. 51 Regarding bacterial infection, SP-A -/- mice exhibit delayed microbial clearance of group B streptococcus, Haemophilus influenzae, and Pseudomonas aeruginosa. 96 SP-D -/- mice clear tested bacteria as efficiently as wild-type mice; 57 however, there is increased inflammation and oxidant production, and reduced macrophage phagocytosis in response to intratracheal instillation of group B streptococcus and H.influenzae. 29 In rats, after the intratracheal inoculation of lipopolysaccharide (LPS), message levels and BAL fluid contents of both SP-A and SP-D significantly increase at 24 hours and 72 hours postinoculation, respectively, underscoring the role of these 2 proteins in the acute phase of lung inflammation. 98 In lambs, during the progression of Mannheimia haemolytica-induced bronchopneumonia into chronicity, the SP-D protein is reduced to near absence in hyperplastic bronchiolar epithelial cells. 52 indicating that the state of epithelial differentiation may influence the SP-D expression. Our recent studies indicate that elevated levels of SP-A and SP-D (as well as sheep β-defensin-1) mRNA in the lungs of neonatal lambs infected with PI-3 virus are associated with simultaneous decrease in PI-3 virus replication. 51 These findings may suggest that lung collectins (and defensins) are synthesized to bind to PI-3 virus, as collectins bind to other viruses, and to neutralize it directly or indirectly.
The second role of lung surfactant proteins is anti-inflammatory. This role can be explored by using SP-A -/- and SP-D -/- mice. The phenotypic features of SP-D -/- mice include chronic low-grade pulmonary inflammation, ATII cell hyperplasia with giant lamellar bodies, excess alveolar surfactant phospholipid, and emphysematous changes with dilated airspaces and pulmonary fibrosis. 25 The inflammation is characterized by peribronchial monocytic infiltrate with an excessive number of alveolar macrophages that produce increased amounts of ROS and matrix metalloproteinases (MMP). Inflammation in this model can be explained by increased lung expression of monocyte chemoattractant protein-1 (MCP-1) mRNA, which can be corrected by the treatment of SP-D -/- mice with recombinant SP-D. 25 Another mechanism of lung inflammation would be increased numbers of apoptotic macrophages present in SP-D -/- mice, because failure in clearing apoptotic cells can lead to triggering of inflammation. 25 SP-A -/- mice do not show detectable abnormalities in histology, lung wet weight, lung volume, compliance, or surfactant lipid metabolism, but clearly demonstrate increased susceptibility to infection by respiratory pathogens, enhanced lung inflammation, and increased incidence of splenic dissemination after intratracheal instillation of bacteria. 53, 62 In addition, surfactant isolated from these mice lacks tubular myelin, but surprisingly these mice have normal postnatal survival with no detectable alteration in pulmonary function. 62
SP-D is capable of regulating an inflammatory response to microorganisms and microbial products. 29 After bacterial challenge, SP-D inhibits pulmonary secretion of the proinflammatory cytokines such as IL-1, IL-6, and TNF-α. 57 SP-A -/- mice show increased production of TNF-α and nitric oxide (NO) upon intratracheal challenge with P. aeruginosa or LPS. 53 In addition, peptidoglycan- and zymosan-induced secretion of TNF-α can be inhibited by direct interaction of SP-A with Toll-like receptor (TLR)-2 on macrophages. 79
The third role of lung collectins is immunomodulation. SP-D mediates inhibition of IL-2 secretion by monocytes, and leads to reduced T-lymphocyte proliferation. 57 It can bind to oligosaccharides associated with dust mite allergens and inhibit the binding of specific IgE molecules to those allergens. 29 Glycosylation of viral coat proteins such as those of IAV, an adaptation that is believed to help viruses evade antibody-mediated neutralization, is associated with enhanced reactivity of SP-D. 117 SP-A can also inhibit proliferation of stimulated lymphocytes and affect IL-2 production. 53 Other functions revealed by in vitro studies include chemotaxis of alveolar macrophages, neutrophils, and monocytes, antioxidant properties, and altered macrophage production of MMPs. 25 One study even suggests that exogenous SP-A can significantly protect lung from injury induced by ischemia-reperfusion of isolated rat lungs by reducing the SP-A loss and by activating the upregulation of endogenous NO synthetase (eNOS). 99
The fourth role, which was the first discovered functional role of lung surfactant proteins, is nonimmunologic and is related to the function of surfactant. 96 SP-A is more important than SP-D in this regard, and has been reported to regulate surfactant homeostasis by controlling the secretion and uptake of surfactant by ATII cells, to protect the surface activity properties of surfactant from functional inhibitors, to inhibit phospholipase A1, and to be required for the formation of tubular myelin. In addition, it has been suggested that an appropriate SP-A level in fetal lung may determine the time of labor. 26 According to this theory, when SP-A reaches a yet undefined critical threshold in the ASL of fetal lung, it stimulates macrophages via TLRs causing proinflammatory cytokine release and macrophage migration to the uterus, which in turn leads to increased uterine contractility and parturition.
Modulation of SP-A and SP-D function
Certain ligands and inflammatory products can serve as competing ligands for collectins that reduce their activity, or can cause their partial cleavage or complete degradation. 29, 65, 120 Glucose concentrations encountered in diabetes mellitus can interfere with the ability of SP-D to interact with specific strains of IAV in vitro and cause increased susceptibility of mice to certain influenza viruses in vivo. 115 Both SP-A and SP-D bind free DNA and the DNA present on apoptotic cells via their CRD's and collagen-like domains, although SP-D binds nucleic acids more effectively at physiological salt conditions. 111 This feature may allow lung collectins to play a role in decreasing the inflammation caused by DNA in lung, which at the same time may inhibit lung collectin carbohydrate binding ability. Interaction of SP-A with other proteins such as fibrinogen can induce inactivation of SP-A. 54 Phosphatidylinositol (PhI) from surfactant may serve as a ligand for SP-D and suppress interaction between SP-D and microorganisms in diseases in which PhI levels are elevated such as in acute respiratory distress syndrome (ARDS). 105 Furthermore, neutrophil serine proteases including cathepsin G, elastase, and proteinase-3 contribute to degradation of SP-A; this may be the reason for low levels of SP-A in patients with cystic fibrosis. 120 These proteases also cleave highly conserved CRD subregions of SP-D markedly reducing the ability of SP-D to promote bacterial aggregation and binding to yeast mannan in vitro. 65 In addition, bacterial products such as P. aeruginosa elastase, a zinc-metalloprotease, also cause formation of nonfunctional SP-D degradation products 4, 146 Finally, it has been shown that the function of SP-A depends on cation concentration, especially calcium, in the environment, which affects the quaternary organization of the SP-A protein. 118
Regulation of SP-A and SP-D expression
Synthesis and secretion of surfactant components are developmentally regulated. Surfactant starts to be synthesized in progressively increasing amounts during the last stage of lung development, the alveolar stage. 77, 100 In humans, SP-A is not detectable in fetal lung until the 30th week, whereas SP-D is first detected in the second trimester of gestation (40 weeks being the full term). 35, 77, 108 In sheep, mRNA content of SP-A is low to nondetectable at 99–100 days of gestation (145–150 days being the full term), and it progressively increases starting at about 120 days of gestation to term and into the postnatal period. 100, 139 Because pre- and full-term infants and animals lack fully effective adaptive immune response and the maternal immunoglobulin levels can vary from very high to low or absent, 37 regulated expression of surfactant is especially important in this age group that largely depends on its innate immune response.
In addition to being dependent on the developmental stage of lung maturation, lung surfactant proteins are also regulated by hormones, growth factors, and other regulatory substances (Table 3). 44 In Table 3, discrepancies in the effect of different factors on SP-A and SP-D expression can be observed, suggesting that these 2 proteins are not coordinately regulated. 35
Regulation of SP-A and SP-D expression by hormones, growth factors, and other regulatory substances.

Tissue distribution
SP-A and SP-D are synthesized by ATII and Clara cells. 28 Alveolar macrophages show strong cytoplasmic and/or membrane labeling with antibody against SP-D; however, because macrophages express no detectable SP-D mRNA, it is likely that intracellular SP-D protein is caused by phagocytosis in conjunction with surfactant metabolism. 29 The lamellar bodies of ATII cells serve as storage organelles for surfactant phospholipids and the content of these organelles is rapidly associated with SP-A upon its secretion into the liquid lining alveoli (hypophase), which leads to formation of tubular myelin. 62, 108 Regulated exocytosis of the lamellar body content involves about 10% of the intracellular surfactant pool per hour, but the availability of SP-A that is free to interact with ATII cell receptors in the intact lung is unknown. 62 The secretory granules of Clara cells, on the other hand, are primarily responsible for storing SP-D and it seems likely that SP-D secretion is regulated via granule exocytosis at this level of the respiratory tract. 29 Although both SP-A and SP-D are expressed in highest amounts in the distal lung, expression at various extrapulmonary sites has been reported and it appears to be species specific. 62, 87 The extrapulmonary sites of SP-A expression include porcine eustachian tube epithelium, 110 rabbit sinus and middle ear mucosa, 38 human small and large intestine, 86 vaginal mucosa, 92 trachea, thymus, pancreas, and prostate. 93 The extrapulmonary sites for SP-D include porcine eustachian tube epithelium, 110 human salivary gland, kidney, heart, small intestine, prostate gland, and pancreas, rat gastric mucosa, and many other sites. 62
Protein structure
Pulmonary collectins are organized as oligomers assembled from multiple copies of a single polypeptide chain consisting of 4 structural domains. 27, 62 The first domain is a short amino-terminal disulfide-rich domain of 7 (SP-A) to 25 (SP-D) amino acids which is involved in the interchain interactions. The second domain is a collagen-like domain that forms an extended fibrillar triple helix 20 (SP-A) to 46 (SP-D) nm long. The third domain forms a short trimeric coiled-coil domain, and links the collagen-like sequence to the fourth domain, globular CRD. SP-A is the most abundant surfactant protein in the lung (5.3% of total surfactant weight) and its monomer measures 28–36 kd. 53, 63 This protein is usually assembled into octadecamers that consist of 6 trimeric subunits that form a “bunch-of-tulips” shape. 27, 63 SP-D is predominantly assembled as dodecamers that contain 4 homotrimeric subunits arranged in a cruciform shape. Each SP-D monomer weighs 43 kd.
Receptors
The best characterized host defense receptor for SP-A is a 210-kd cell surface protein (SP-R210) present on rat ATII cells and alveolar macrophages. 68, 69 The interaction of SP-A and SP-R210 occurs through the collagen-like domain of SP-A, and leads to binding of SP-A to ATII cells and alveolar macrophages, SP-A-mediated uptake and killing of Mycobacterium bovis, and inhibition of phospholipid secretion. 68, 96 Both SP-A and SP-D bind to glycoprotein gp-340, a member of the macrophage scavenger receptor cysteine-rich protein family that has both soluble and alveolar macrophage membrane-bound components. 96 The interaction of lung collectins with gp-340 leads to binding and aggregation of some bacteria, such as Streptococcus mutans. 113 SP-A also binds to TLR2 resulting in reduced TLR2 binding to peptidoglycans. 87 Other proposed receptors for collectins include cC1qR (calreticulin), CD14, and 30-kd alveolar cell membrane protein. 57, 63
Lung collectin genes
In humans and nonhuman primates, SP-A is encoded by 2 genes, SP-A1 and SP-A2, with several alleles at each locus. 76, 83 In other species such as rabbits, rats, mice, and dogs SP-A is encoded by a single-copy gene. 83 Human SP-D is encoded by a single gene, which has also been confirmed in rodents and ruminants. 27, 48 Both human SP-A and SP-D genes are found on the long arm of chromosome 10 at q22.2–23.1 region, near the MBL gene. 27 Bovine SP-D gene is found at the same locus as the conglutinin gene on Bos taurus chromosome 28 (BTA 28) at position q1.8, and comprises 9 exons spanning approximately 10.5 kb. 49 The bovine gene encoding SP-A is located proximal to the bovine SP-D gene at BTA 28 at position q1.8–1.9. 48 The order of bovine genes for SP-A, MBL-A, and SP-D is homologous with the order of the analogous genes found in humans and mice. Regulation of lung surfactant protein gene expression is multifactorial and controlled by transcriptional and/or post-transcriptional (mRNA stability) mechanisms. 14 The function of surfactant protein promoter depends on the combined activity of many transcription factors, one of them being thyroid transcription factor 1 (TTF-1/Nkx2.1), a common positive regulator of surfactant protein promoter activity. 14
Cationic Antimicrobial Peptides
Two major families of cationic AMP are defensins and cathelicidins. 78 Defensins can be further divided into 3 classes, α-, β-, and Θ-defensins based on pairing of their 6 cysteine residues in the disulfide bridges. 32, 140, 149 Human α-defensins are 29–35 amino acids in length and contain 3 disulfide bridges in a 1–6, 2–4, 3–5 arrangement. 78 These peptides were first described in human tissues and include 6 members. 140 Four members, named human neutrophil peptides (HNPs) 1–4, have been found in the azurophilic granules of neutrophils. The other 2, human defensin (HD)-5 and HD-6, are located in secretory granules of intestinal Paneth's cells and in female genital epithelial cells. In the early 1990s, β-defensins were identified. 155 β-defensins are 36–42 amino acids in length and have 3 disulfide bridges arranged in a 1–5, 2–4, 3–6 pattern. 78 More than 28 human β-defensins have been identified 7 among which the best studied include human β-defensin (HBD)-1 expressed in the epithelial cells of respiratory tract including lung and trachea, pancreas, kidney, gastrointestinal epithelium, 72, 97 urogenital tract, 157 and reproductive tract; 84 HBD-2 expressed in skin, trachea, and lung; 130 and HBD-3 expressed predominantly in the skin but is also expressed in the respiratory tract. 74 In mice, over 40 β-defensins have been identified including mouse β-defensin (MBD)-1, -2, -3, and -4. 7 MBD-1 is homologous to HBD-1 at the gene level, and is found in multiple epithelia with highest levels in kidney, 8 whereas MBD-3 exhibits homology at the gene level to HBD-2, and is detected in lung, salivary gland, and reproductive organs. 8 Bovine β-defensin family members include neutrophil defensins (BNBD) 1–13 that are expressed in bovine neutrophils and macrophages; tracheal antimicrobial peptide (TAP) expressed in nasal epithelium, trachea, bronchioles, and alveolar macrophages; and lingual antimicrobial peptide (LAP) expressed in the epithelial cells of tongue and alveolar macrophages. 17, 43 Two ovine β-defensins have been described, sheep β-defensin (SBD)-1 and -2. SBD-1 expression has been detected from the tongue to colon with the exception of distal ileum where SBD-2 predominates. 72, 101 SBD-1 is also expressed in the trachea, 72 whereas both SBD-1 and SBD-2 are expressed in the lung. 2, 51
Cathelicidins, another important family of cationic AMP, have been found in cows, pigs, sheep, goats, horses, mice, guinea pigs, rabbits, and humans. 17, 132 Their main structural feature is a high-level sequence identity in the 5′ region, named the N-terminal cathelin domain because it is also found in cathelin, a cysteine protease inhibitor. The C-terminal domain of cathelicidins provides them with antimicrobial activity. In most species, cathelicidins are expressed in myeloid precursor cells. Other cells and tissues known to express cathelicidins include mature peripheral porcine neutrophils, human airway epithelium, and the testis. 9, 114 Some examples of cathelicidins include bovine myeloid antimicrobial peptide (BMAP)-34, BMAP28, Bac5, Bac7 in bovine; OaBac11, sheep myeloid antimicrobial peptide (SMAP)-29, SMAP34 in sheep; porcine myeloid antimicrobial peptide (PMAP)-23, cecropin P1, PR-39, protegrin-1 in pigs; equine cathelicidins eCATH-1, -2, and -3 in horses; and LL-37 (also known as hCAP-18) in humans. 17, 154
Functional roles
Cationic AMPs have multiple roles and the best-studied ones are included in Table 4. These peptides have the potential to kill or neutralize bacteria (Gram-positive and Gram-negative), fungi (including yeasts), viruses (even enveloped, like human immunodeficiency virus [HIV]), and parasites (including planaria, nematodes, trypanosomes, and plasmodia). 132 The peptides are selective in that normal host cells are relatively resistant to their action. This is based on a few important differences between cytoplasmic membranes of prokaryotic and eukaryotic cells. 95 The most striking difference is the composition and topological arrangement of lipids. The outer surface of mammalian cell membranes is composed of electrically neutral (zwitterionic phospholipids), mainly phosphatidylcholine (PC) and sphingomyelin. Bacterial membranes, on the other hand, contain large amounts of negatively charged phospholipids, phosphatidylglycerole (PG) and cardiolipin (CL). In addition, the outer membranes of Gram-negative bacteria are covered with polyanionic LPS. Therefore, cationic peptides prefer binding to bacteria because of electrostatic interactions between positive charges of the peptides and negative charges of the lipids. The second important difference is that unlike eukaryotic cell membranes, prokaryotic cell membranes lack cholesterol, which also contributes to prokaryotic cells being a better target. The last striking differentiating factor of prokaryotic cells is that they possess a large transmembrane potential of -140 mV, compared to the low membrane potential of eukaryotic cells (15 mV). Unfortunately, there are some exceptions to these rules and certain cationic AMP peptides, like melittin from bees, mastoparan from wasps, or charybdotoxin from scorpions, are potent toxins. 132
Multifunctional roles of cationic antimicrobial peptides (AMP).

Given the diversity of AMP and of target microorganisms, it is likely that there is not one unique mechanism for their mode of action. 39 The only constant is that these peptides do not need a metabolically active cell to exert their effect. Based on studies done with magainins, cationic AMP isolated from frog skin, a general mechanism of action on Gram-negative (2-cell membrane) bacteria has been proposed. 95 It includes 2 steps, with a note that the second step could also be applied to Gram-positive (1-cell membrane) bacteria. In the first step, positively charged AMP interacts with polyanionic LPS at sites of LPS where divalent cations usually bind to cross bridge adjacent LPS molecules and stabilize the outer membrane. The competitive displacement of these divalent cations (Ca2+, Mg2+) by the bulkier peptides leads to lesions in the outer bacterial membrane visualized as surface blebbing. This model has been described by Robert Hancock, and is called “self-promoted uptake.” 56 In the second step, AMP associates with the negatively charged phospholipid membrane and form holes (“aggregate channels”) in the membrane when certain, critical AMP concentrations have been reached. Examples of peptides that cause formation of membrane channels are cecropins and defensins. 17 Other cationic AMP like Bac5 and Bac7 alter transport and energy metabolism in the cytoplasmic membrane of pathogens. Once the peptides are within the cells, they can bind to cytoplasmic polyanions such as DNA and interfere with DNA/protein synthesis (e.g., PR-39), and then stimulate autolytic enzymes. 17, 39 It has also been demonstrated that AMP can inactivate bacterial heat-shock proteins (DnaK) without binding to human analogous heat shock protein Hsp70 and affecting its normal function. 109 The mechanism of antiviral action is less understood, but studies show that β-defensins can cause disruption of the viral envelope, whereas α-defensins may also interfere with intracellular cell signaling inhibiting viral replication. 23, 102
Modulation of AMP function
In vitro experiments have shown that defensins are microbicidal at μM concentrations against many Gram-positive and Gram-negative bacteria, yeast, fungi, and some enveloped viruses if there is low-salt concentration (e.g., 10 mM sodium phosphate) in the environment. 43 As the salt concentration increases, the activity of lung innate antibacterial factors such as HBD-1 and -2, as well as lactoferrin, lysozyme, histatin, and cathelicidin can be completely inhibited, which likely occurs in the lungs of patients with cystic fibrosis. 127 Using the osmolyte xylitol to reduce airway surface liquid salt concentration, 153 and/or exogenously introducing cationic AMP that are not salt-sensitive (e.g., sheep cathelicidin SMAP29) are showing promising results in enhancing AMP activity. 134 Another factor influencing AMP activity is pH, which can affect the electrical charge of peptides. 136 This may lead to changes in peptide conformation and activity. The ability of some peptides to be active at low pH conditions can be used for drug generation, because this feature would allow the peptides to act in phagolysosomes. Furthermore, because of their cationic properties, cationic AMP easily forms complexes with various serum proteins like serum albumin and α2-macroglobulin, or bacterial degradation products of extracellular matrix, which may abolish their activity. 1, 70
Regulation of expression
All cationic AMPs can have constitutive (normally present in animals) and/or inducible (upon stimulation) expression. 78 Two examples of peptides with constitutive properties are HBD-1 and MBD-1. The expression of inducible peptides can be stimulated during infection or inflammation with LPS, lipoteichoic acid (LTA), TNF, IL-1β, IFN-γ, and other mediators of inflammation. 55, 58 These AMP include human HBD-2, -3, -4, and LL-37, 74, 78, 89 bovine LAP and TAP, 33, 121 mouse MBD-2 and -3, 78 and others. In addition, like with collectins, the expression of pulmonary defensins seems to be developmentally regulated in late gestation and through the neonatal period, 101 which leaves a window of immature peptide expression in the infant that may provide an environment with increased susceptibility to respiratory tract infection.
The mechanism of AMP gene induction is poorly understood. One of the proposed induction pathways is LPS-mediated upregulation of AMP genes, which has been directly shown for the TAP gene in bovine and HBD-2 gene in human airway epithelial cells. 133 Cationic AMP also bind LTA, although with a lower affinity compared to LPS, leading in some cases to production of TNF-α and IL-6 by murine macrophages. 132 In addition, recent studies indicate that retinoic acid (RA) may have an important effect on AMP gene expression. 59 In porcine bone marrow cells, RA induced expression of cathelicidin PR-39; however, in human keratinocytes, induction of HBD-2, -3, and -4 mediated by high Ca2+ concentration, proinflammatory cytokines, phorbol myristate acetate (PMA), and bacteria, was abolished when keratinocytes were preincubated with RA. The biological effect of RA is achieved by its binding to and activation of the nuclear retinoic acid receptors (RAR) that belongs to the steroid/thyroid receptor family. 42, 59 RAR, in turn, form heterodimers with the retinoid x receptors, which is followed by its direct binding to specific cis-acting retinoic acid response elements or by its antagonistic effect on AP-1 (c-Jun/c-Fos)-mediated gene expression. Because the promoter regions of all inducible HBD genes contain putative binding sites for AP-1, it is possible that AP-1 plays an important role in the signal transduction during induction and downregulation of defensin gene expression. 59
Peptide characteristics
Cationic AMP are 12–50 amino acids in length, and have a net positive charge (+2 to +7) owing to an excess of basic amino acids (arginine, lysine, and histidine). 55 Different AMP are needed to deal with different microorganisms and because of this, single animal species can have more than 24 different AMPs. 17 AMPs fall into 4 principal categories based on their size, conformational structure, or predominant amino acid structure. 17, 78 These comprise group I: linear, α-helical peptides without cysteines (e.g., SMAP and porcine cecropin P1); group II: β-sheet structures stabilized by 2–3 disulfide bridges (e.g., α- and β-defensins, rhesus theta defensin-1, and protegrins); group III: linear peptides rich in 1 or more amino acids (e.g., PR-39, Bac5, Bac7, indolicidin, and prophenin); and group IV: peptides with loop structures (e.g., bactenecin and ranalexin). In general, cationic AMPs with best antimicrobial activity are molecules that have the charged and hydrophilic portions separated from the hydrophobic areas. This means that either amphipathic or cationic double-wing structures with a hydrophobic core separating 2 charged segments are preferred, which is determined by their tertiary structure. 132 There is also a group of cationic AMP, which are formed by proteolytic digestion of larger peptides, such as lactoferricins that are derived from lactoferrin. 15
Cationic AMP genes
AMPs of vertebrates are ribosomally synthesized, gene-encoded peptides, meaning that 1 gene codes for 1 peptide. 78 In humans, mice, and rats, both α- and β-defensins colocalize on the chromosome region that is on human chromosome 8p21–23, mouse chromosome 8, and rat chromosome 16. 55, 90, 131 In sheep, β-defensin genes that comprise two exons (e.g. SBD-1 and -2) have been mapped to sheep chromosome 26, whereas the 4-exon cathelicidin genes have been mapped to sheep chromosome 19, suggesting homology between human, sheep, cattle, and mouse AMP gene families. 73 AMP genes have intron–exon structure with regulatory elements in their promoter regions. The primary translational product of exons is a prepropeptide, which consists of an N-terminal signal sequence for targeting of the endoplasmic reticulum, a pro segment for correct folding of the mature peptide, intracellular trafficking, and/or inhibition of activity of the mature peptide, and a C-terminal cationic peptide that has antimicrobial activity after cleavage. 78, 81 Cleavage of the propeptide occurs during later stages of intracellular processing or after secretion. AMP can be stored in cells as propeptides or as mature C-terminal peptides.
Conclusions
A significant amount of evidence from both in vitro and in vivo studies suggests that SP-A and SP-D serve numerous functions in innate immune processes. The use of recombinant lung surfactant proteins in the treatment of diseases such as respiratory distress syndrome in neonates, emphysema and chronic obstructive pulmonary disease, cystic fibrosis, and asthma has already shown promising results. 24, 25 Development of collectin-based therapies for various pulmonary diseases warrants further investigation.
With increasing development of antibiotic resistance, there is a need to develop new classes of antibiotics. Cationic AMPs have many of the desirable features of such substances, which include a broad spectrum activity, rapid killing of bacteria, activity against classic antibiotic-resistant variants, synergy with antibiotics, neutralization of endotoxins, and activity in vivo. 132 However, despite all these positive features, there are still many issues that need to be addressed in order for these molecules to be used as systemic therapeutic agents. These include production costs as a result of their high molecular weight, potential toxicity, and peptide liability to proteases in the body. 55
Footnotes
Acknowledgements
This work was supported in part by the National Institutes of Health NIAID Awards 5R01AI062787-02 and 5K08AI055499-03, USDA/CSREES/NRI-CGP 2003-35204-13492, and the JG Salsbury Endowment. Special thanks to Jack M. Gallup for his contributions to our projects.