
Abstract
The response of the preterm and newborn lung to airborne pathogens, particles, and other insults is initially dependent on innate immune responses since adaptive responses may not fully mature and require weeks for sufficient responses to antigenic stimuli. Foreign material and microbial agents trigger soluble, cell surface, and cytoplasmic receptors that activate signaling cascades that invoke release of surfactant proteins, defensins, interferons, lactoferrin, oxidative products, and other innate immune substances that have antimicrobial activity, which can also influence adaptive responses. For viral infections such as respiratory syncytial virus (RSV), the pulmonary innate immune responses has an essential role in defense as there are no fully effective vaccines or therapies for RSV infections of humans and reinfections are common. Understanding the innate immune response by the preterm and newborn lung may lead to preventive strategies and more effective therapeutic regimens.
With the first breath after birth, the lung is exposed to a wide variety of airborne substances and microbial agents. Despite this, most newborns do not develop extensive pulmonary inflammatory/immune responses that would impair airflow and gaseous exchange. This is due, at least in part, to the innate immune system’s clearance of particulate matter and discrete handling of other substances and microbial agents. The response to pathogens and foreign material, vapors, fluids, aerosols, mists, and other substances by the perinatal lung is especially dependent on innate immune responses since adaptive immune responses are not fully mature and can require weeks for sufficient responses to antigenic stimuli. The innate immune system of the lung includes the mucociliary apparatus, air-surface liquid and its contents, epithelia, serum products, dendritic cells, and other leukocytes (Fig. 1). Foreign material and microbial agents trigger soluble, cell surface, and cytoplasmic receptors that activate signaling cascades that trigger release of surfactant proteins, defensins, interferons, lactoferrin, oxidative products, and other innate immune substances that have antimicrobial activity and can also invoke adaptive responses. For viral infections such as respiratory syncytial virus (RSV), innate immune responses are increasingly appreciated for their role in reducing disease severity. RSV is a common respiratory pathogen worldwide and can cause severe bronchiolitis and respiratory disease resulting in hospitalization in infants (especially preterm), immunosuppressed individuals, and the elderly.21,97 Each of these categories of people has less than optimal adaptive immune responses, and thus innate immune responses become even more vital since there are no approved vaccines or fully satisfactory therapies. Vaccine development for RSV in humans has been hindered by deaths that occurred in RSV-infected infants previously vaccinated with a formalin-inactivated vaccine in a trial in the 1960s. 21 Although RSV is a ubiquitous virus and most women have circulating antibodies, preterm infants may have a limited amount maternal antibody because the shortened gestational time of an infant born preterm reduces the amount of time for transplacental passage of maternal immunoglobulin. Therefore, some preterm infants have limited maternal antibody, their immune system has not ever been exposed to RSV or many other antigens, and their adaptive immune system is not fully mature. Such individuals are heavily dependent on their innate immune response for protection against severe RSV infection. Also, RSV and other viruses such as influenza infect bronchiolar epithelial cells and, once infected, become inflamed, impairing airflow and thus the gaseous exchange function of the lung. Reducing bronchiolar epithelial cell infection is vital, especially for RSV since there are no fully satisfactory therapies or approved vaccines. Cattle and sheep are also susceptible to RSV infections, especially during times of stress, as can occur with shipping, weaning, overcrowding, poor management, and improper environmental and housing conditions. Such stress can also impair adaptive and innate immune responses, leading to increased susceptibility. Our laboratory has developed a lamb model of RSV infection that closely mimics key features of RSV infection in newborn infants. 23 This review addresses key features of the respiratory innate immune system of the perinatal lung, comparing newborn and adult. It highlights responses to one specific virus, RSV, which is a serious and common pathogen of infants, cattle, and sheep.
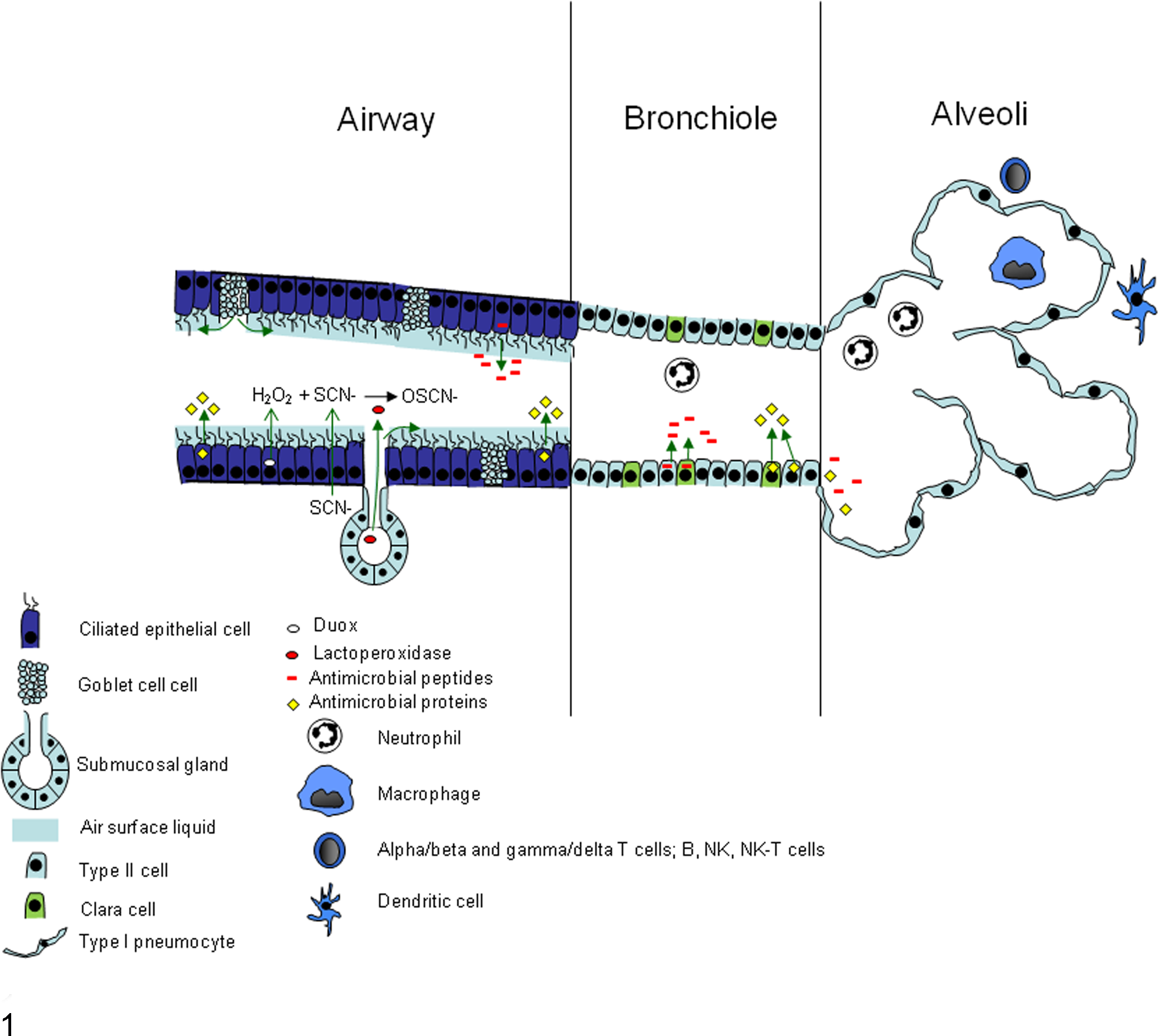
Schematic illustration depicting some of the innate immune systems of the lung airways and alveoli. This image was drawn by M. R. Ackermann and published in Ackermann MR, Derscheid RM, Roth JA. Innate immunology of bovine respiratory disease. Vet Clin North Am Food Anim Pract. 2010;26:215–228.
Components of the Innate Immune System and Responses to RSV
The newborn lung is challenged by foreign material and airborne stimuli that can invoke cellular responses. The molecular machinery of receptor recognition, signaling, activation, or suppression of transcription factors, messenger RNA (mRNA) transcription, translation, protein translocation, and activity is triggered to a newly experienced level by the various lung cell types while the lung itself transitions to its primary function: gaseous exchanges. This dynamic activity is even more complicated in newborns that are preterm or premature and also in those with physiologic stress, inherited disorders, or pathologic conditions. Investigations of the perinatal lung of human infants have limitations in monitoring and measuring cytokine, chemokine, cellular, and humoral responses, particularly localized responses. The methods used must generally be minimally invasive, which limits the amount and type of data that can be collected from the pulmonary response to microbial agents. In addition, variations in disease diagnosis as well as method of sample collection create difficulties in comparisons between studies. Despite these challenges, much has been learned from human subjects and while knowledge gaps are addressed in a wide variety of animal models.
In perinatal, maturing, and adult lungs, the initial diseases or stresses caused by viruses, other microbial agents, and their products, antigens, allergens, foreign material, vapors, and substances arriving in the lung via the vasculature have an acute onset and typically a fairly short duration, although there can be protracted or recurring symptoms, depending on the inciting stimulus. Minimal or mild exposure of these insults is handled every moment by the pulmonary tract of healthy individuals without incident. Because airflow and gaseous exchange are optimal in the noninflamed lung, it is beneficial for the inflammatory response to be as minimal and rapid as possible. Mucociliary clearance and physiologic activity of innate immune products secreted onto the air-surface liquid (ASL) are of special importance in this regard to avoid/reduce epithelial cell damage, bronchoconstriction, vascular leakage, and an overt inflammatory or adaptive response. There is a range of responses by individuals and severity of disease, with some individuals at increased risk due to altered lung structure or function and potentially a reduced ability to develop an adaptive response compared with others who typically experience mild disease. Such variations between individuals range from differences in gross structure (eg, diameter of the nasal meatus or length of the trachea), epithelia (eg, numbers and distribution of goblet cells), immune system (eg, distribution of dendritic cells or mast cells), and genetics (eg, single-nucleotide polymorphisms and gene copy number of cytochrome P450 isoenzymes, defensins, or surfactant proteins A and D).
The physical innate barriers of mucous production, mucociliary elevator apparatus, submucosal gland secretions, and complex branching of the respiratory tree must initially be overcome by microbial agents such as RSV to reach bronchiolar epithelium and respiratory airways where the virus establishes infection and replicates. 98 Once a virus, microbial product, allergen, antigen, or foreign material deposits onto the mucosal surface of the respiratory airways, and it comes in contact with the mucosal secretions and the ASL.
In trachea, bronchi, and, to some degree, the bronchioles, the mucosal secretory layer of these airways is composed of periciliary sol and gel layers of the mucociliary apparatus formed from secretions from goblet cells, submucosal glands, and respiratory epithelial cells. The periciliary sol layer contains water that accumulates with chloride released from nearby epithelial cells, submucosal glands, and serous cells. The sol layer contains tethered mucins that form a “brush” layer on which the overlying mucus layer sits. 18 The sol (also termed ciliary brush) layer allows cilia beat activity maintained in the proper pH, and the amount of water in the sol layer is regulated by sodium (Na+) resorption by epithelial Na+ channels (ENaC). With dehydration, the sol (ciliary brush) layer becomes thinner and the tethered mucins aggregate, both of which decrease ciliary beat. Certain other conditions such as cystic fibrosis and chronic obstructive pulmonary disease (COPD) can alter the consistency of the sol (ciliary brush) layer. 18 The gel layer is composed of mucin glycoproteins encoded by 22 MUC genes, of which 16 have been identified in the human lung, including MUC1, MUC2, MUC4, MUC5 AC, MUC5B, MUC7, MUC8, MUC MUC11, MUC13, MUC15, MUC16, MUC18, MUC19, MUC20, MUC21, and MUC22. These proteins are linked to oligosaccharides and localize along the apical epithelial surface, some in a secreted form, others tethered to the cell membrane. Some, such as MUC5 AC, are secreted by goblet cells, whereas others, such as MUC5B, are secreted from mucus cells and submucosal glands. 64 MUC1 is membrane tethered and expressed by lung epithelial cells, including type II cells, 64 and has anti-inflammatory properties through inhibition of Toll-like receptor (TLR) signaling. With RSV infection, mucin production is regulated by TLR7, interleukin (IL)–17, and IL-23. 85 Expression of MUC genes can be altered by acute disease conditions such as preterm birth, dehydration, heat, smoke, and particulate matter and chronic conditions such as allergic conditions, COPD, recurrent airway obstruction (horses), toxins (eg, bleomycin), chronic infections, cystic fibrosis, and primary or metastatic neoplasia. 18,64
Ciliary beat moves the pericellular ASL in an anterior direction in healthy individuals and various rates, which can be measured. Certain disease conditions, such primary ciliary dyskinesia (immotile ciliary syndrome, Kartagener syndrome), exposure to certain toxins (eg, ethanol, cigarette smoke), and infectious agents (eg, Mycoplasma sp), can drastically reduce ciliary activity and thus impair physical clearance of the materials trapped within the ASL. Other conditions, such as cystic fibrosis and severe dehydration, alter the hydration level of the ASL, resulting in increased viscosity and a thick, mucinous layer that is not adequately propelled in an anterior direction by the cilia. 18
Submucosal glands are present in the nasal cavity, trachea, and bronchi of humans, cattle, sheep, and pigs; however, in rodents, submucosal glands are limited to the upper airways. 17,64 Submucosal glands work in concert with respiratory epithelium to create an oxidative host defense system at the mucosal surface through production of secretory products that contribute to the mucosal gel and sol layers. Submucosal glands produce lactoperoxidase (LPO), which works in concert with other enzymes and molecules to form oxidative radicals within the ASL that contribute to antimicrobial defense. Airway epithelia adjacent to submucosal glands transport thiocyanate (SCN–) to the epithelial surface and also produce hydrogen peroxide (H2O2) via intracellular dual oxidases (Duox). 22,38,142 LPO secreted by the submucosal glands onto the air-surface liquid catalyzes the conversion of H2O2 and SCN– to oxythiocyanate (OSCN–). The Duox/LPO system has microbicidal activity against multiple bacteria and viruses. 22,142 In vitro assays that substitute I– for SCN– in the LPO/Duox system have shown killing of RSV to the same level of bleach. 37 Submucosal glands also secrete lactoferrin and lysozyme, 2 important factors in innate immunity, especially in the air-surface liquid ASL. Lactoferrin is an antimicrobial glycoprotein that not only binds pathogens directly but acts as an immunomodulatory protein and bridges the innate-adaptive immunity crossover. 73 –75 Pneumocytes (alveolar type I cells) make minor contributions to the amount of lactoferrin and lysozyme present in ASL. 27
Tracheal, bronchial, and bronchiolar epithelial cells have an active role in immunity through the secretion of immunomodulatory compounds with innate antimicrobial activity, as well as secretion of cytokines and chemokines upon infection to recruit immune cells. Epithelial cells produce innate immune molecules with anti-RSV activity. Surfactant proteins A and D (SP-A and SP-D, respectively) are collectins that bind pathogens with the globular head region (mannose-binding C-type lectin) and signal with the collagen-like tail. Both SP-A and SP-D are produced and secreted into the airway by primarily type II epithelial cells with a lesser contribution by Clara cells. SP-A and SP-D are reduced in infants with RSV disease who require ventilator assistance. 62 In cultured cell studies, SP-A and SP-D mRNA expression is increased by RSV infection, but protein expression is decreased, putatively through decreased translational efficiency. 16 SP-A binds mannose residues of microbial agents and to RSV F protein, resulting in increased viral clearance. 77,117 SP-D enhances phagocytosis and clearance of RSV in a mouse model. 76 Both SP-A and SP-D enhance RSV elimination. 43 Genetic polymorphisms in SP-A or SP-D are associated with altered severity of RSV infection, 3,28,69 underscoring the essential role of these collectins in antiviral defense. Expression of SP-A and SP-D increases throughout fetal lung development in lambs. 91,133 RSV infection of lambs enhances expression of SP-A and SP-D and is associated with viral clearance. 44,61,101,102 Drugs, such as ethanol exposure in utero, reduce SP-A production in lungs of newborns and may explain, at least in part, why ethanol consumption by pregnant mothers is a risk factor for severe RSV disease. 72
Many other proteins produced by respiratory epithelia contribute to innate defense; however, often their activity and function is not completely defined (Tables 1 and 2). Secretory leukocyte protease inhibitor (SLPI) is a serine antiprotease and, as such, protects the lung against enzymes released from neutrophils and other leukocytes, but SLPI also has antimicrobial and antiviral activity. 118,139 Palate, lung, and nasal epithelium (PLUNC) proteins are produced at high levels by upper respiratory epithelia and have homology to lipopolysaccharide (LPS) binding protein and bacterial/permeability-increasing protein (BPI), suggesting a role for PLUNC in LPS binding. 118,139 BPI protein can bind LPS and also has bactericidal activity against Gram-negative bacteria. Ribonuclease 7 (RNase 7) has been identified in skin epithelia (and thus is likely also present in the stratified squamous epithelium of the anterior nasal meatus) and has potent bactericidal activity. Lipocalin (also known as neutrophil gelatinase-associated lipocalin) can bind bacterial siderophores. 118
Antimicrobial products from epithelia of the respiratory airways and alveoli that contribute to innate defense.
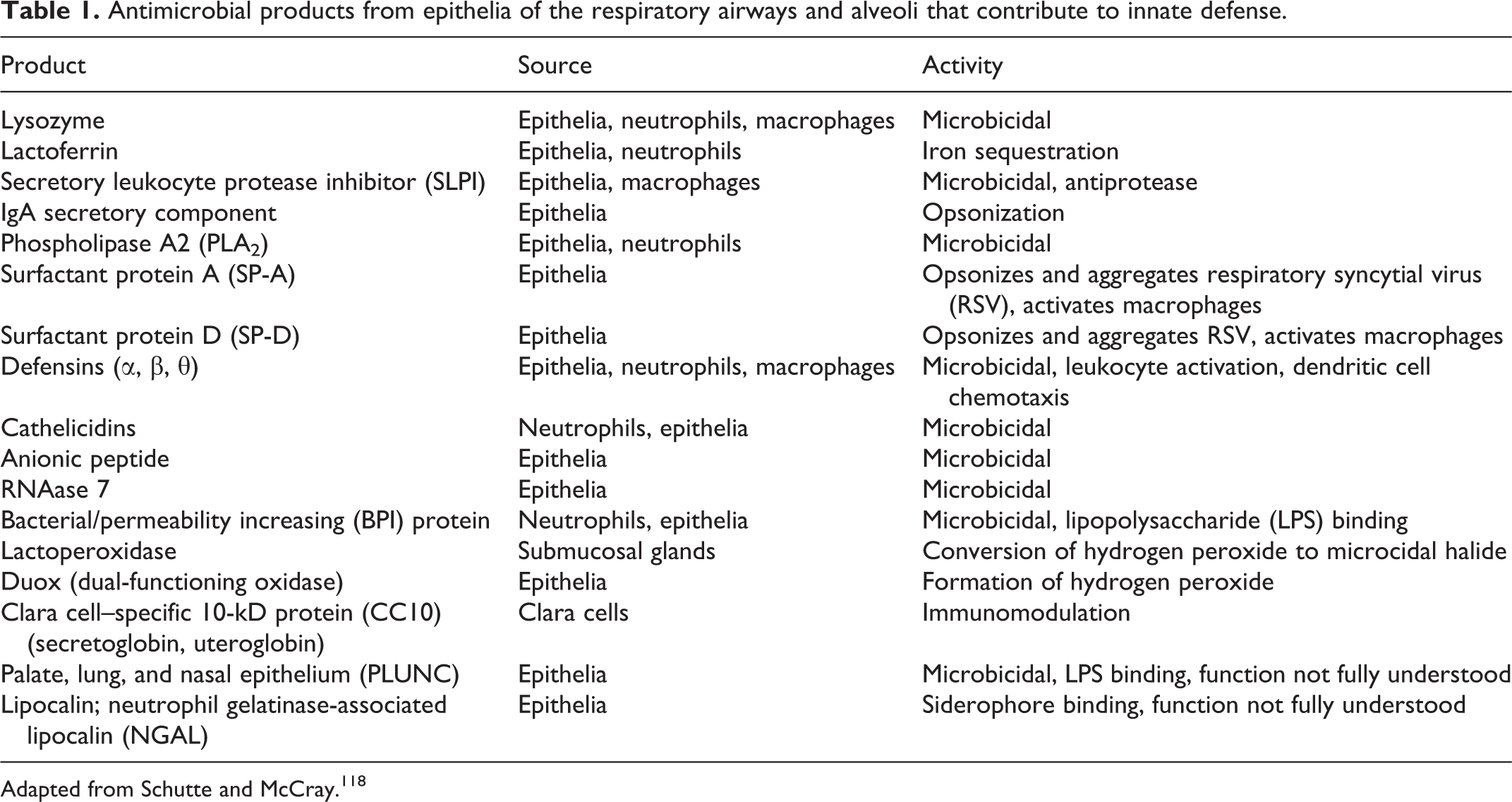
Adapted from Schutte and McCray. 118
Innate immune features of the respiratory tract of infants/neonates and responses to respiratory syncytial virus (RSV).

β-Defensins are antimicrobial cationic peptides also produced by respiratory epithelium, including pneumocytes and submucosal glands, 95,118,139 of many species, including all domestic animals, rodents, and avian species. The β-defensins have direct antimicrobial activity against a wide variety of microbial pathogens and also have numerous other functions, including leukocyte and dendritic cell chemotaxis, epithelial proliferation, mast cell degranulation, and immunomodulatory activity. Human β-defensins (HBD) 1 to 4 are produced in the respiratory tract. HBD-1 is constitutively expressed, whereas HBD-2 to HBD-4 are inducible. 52 In vitro infection of lung epithelia induces HBD-2 production by tumor necrosis factor (TNF)–α via a nuclear factor–κβ (NF-κβ)–dependent mechanism. Secreted HBD-2 then disrupts the viral envelope, interfering with viral entry into host cells. 65 α-Defensins are produced by neutrophils and enterocytes, including Paneth cells, and θ-defensins are present in some nonhuman primates but are a pseudogene in humans. Similar to the β-defensins, the human cathelicidin LL37 is a cationic antimicrobial peptide. 139 LL37 is stored in neutrophils and produced by other leukocytes as well as respiratory epithelium at a low level. LL37 can be upregulated in disease and plays a role in direct microbial killing as well as in immunomodulation and apoptotic signaling. 8,13,81,139 Sheep produce a cathelicidin, SMAP29, which has potent antibacterial activity, and sheep also express sheep β-defensin-1 (SBD-1), which increases with paramyxoviral infection (RSV is a paramyxovirus) and is associated with viral clearance. 15
Clara cells are nonciliated bronchiolar epithelial cells with multiple roles in the airways. Clara cells biometabolize xenobiotics, 19 secrete immunomodulatory substances, 128 and act as progenitor cells. 11 Just as type II pneumocytes serve as a proliferation pool to replace dead and damaged type I pneumocytes, Clara cells act as a progenitor for type II cells, forming a proliferation pool that is vulnerable to exhaustion, especially in neonates, 123,124 but also in chronic smokers (chronic toxin exposure). 14 Furthermore, damage to or dysfunction of Clara cells creates a proinflammatory environment due to the loss of their immunomodulatory secretions. 29 In their immunomodulatory capacity, Clara cells secrete a unique protein: Clara cell secretory protein (CC10), also known as CCSP, CC16, secretoglobin, and uteroglobin. CC10 expression is increased throughout ontogeny (Fig. 2) and CC10 levels in bronchoalveolar lavage fluid and serum during acute injury such as smoke inhalation or application of pneumotoxicants (naphthalene, 4-ipomeanol), but it is decreased in chronic or dysplastic airway dysfunction, such as asthma, COPD, or bronchopulmonary dysplasia (BPD). 112 Infants who developed BPD had lower levels of CC10 at birth than age-matched infants who did not develop BPD. 112,121,129,130 Gene knockout studies performed in mice have shown increased inflammation and viral persistence in CC10-deficient mice when challenged with RSV; restoration of CC10 abrogated these effects. 141 Amniotic levels of CC10 during mid-trimester were significantly higher in women who had preterm premature rupture of membranes compared with women who did not, and this elevation of CC10 could be in response to a proinflammatory event. 106 Although CC10 is the most studied secretion of Clara cells, Clara cells also produce SP-A and SP-D. 100,128 Clara cell cytochrome P450 enzymes such as CYP1A1 and CYP1A2 detoxify inhaled compounds and compounds entering the lung hematogenously. This is beneficial for some toxins; however, a toxic metabolite, 3-methylindololam-ine, is a toxic metabolite formed by Clara cell metabolism of 3-methyl indole.
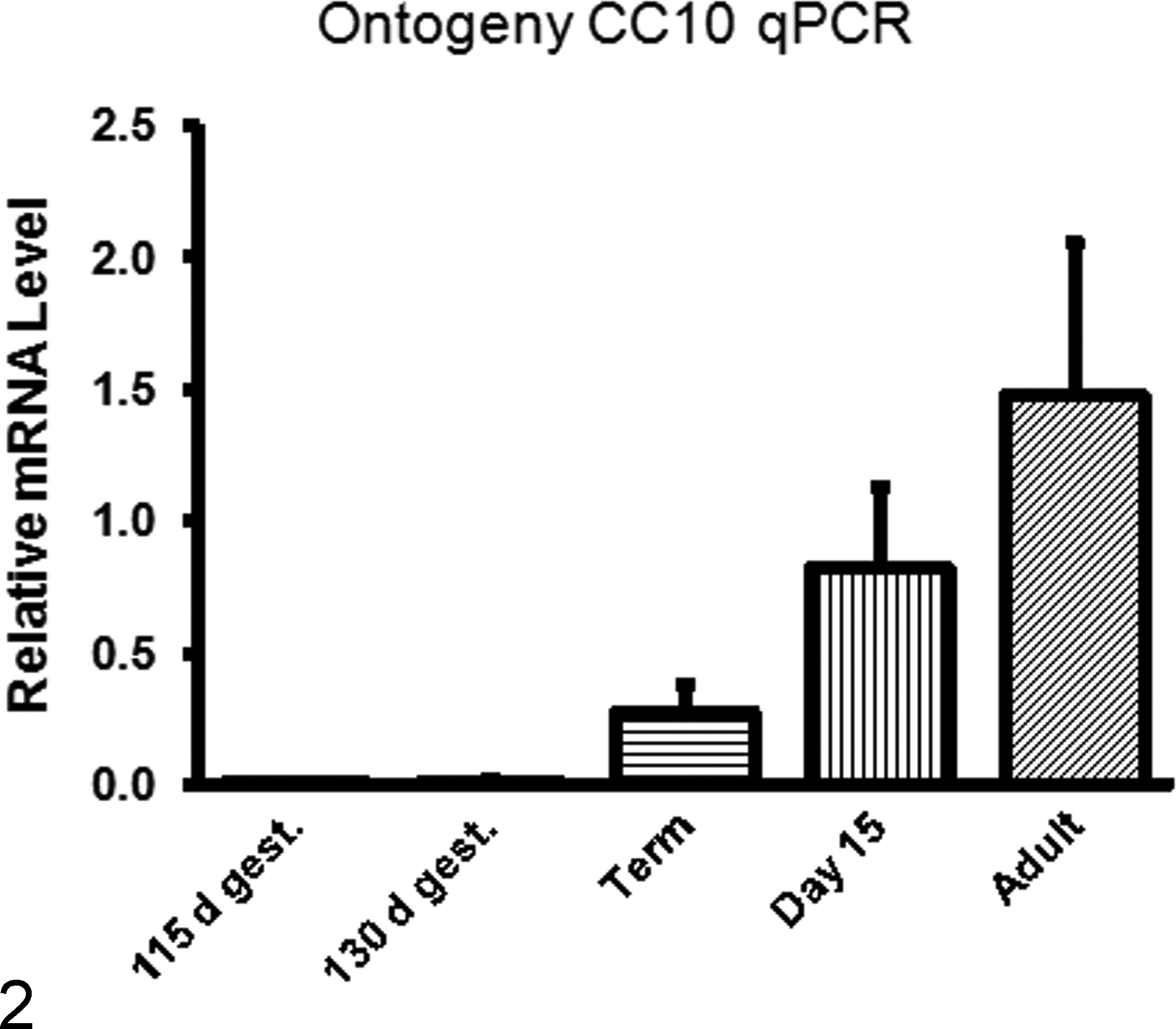
Expression of CC10 in the lung of lambs during ontogeny. CC10 messenger RNA (mRNA) levels in preterm lung are very low in fetal lung and increase progressively with age (manuscript in preparation). Susceptibility to respiratory syncytial virus (RSV) is increased in preterm infants (and lambs) and reduced expression of innate immune products, such as CC10 may underlie the increased susceptibility to RSV infection. For each time point, n = 4 lambs/group and RNA levels were determined by reverse transcription quantitative polymerase chain reaction (RT-qPCR) using 0.784 ng RNA/μl per sample and assessing levels to a Stock I–derived standard curve for CC10. Results were normalized to total lung RNA-loaded RT-qPCR. Results were assessed by GraphPad Prism 6 (GraphPad Software, La Jolla, CA) with a 1-way analysis of variance followed by Tukey’s posttest.
Type II cells of lung increase in number and distribution and also differentiate progressively in developing ovine fetal lung 17,89,91 and lungs of other animals. Type II cells proliferate to replace themselves and also differentiate into type I cells that line pulmonary alveoli. Type I cells produce surfactant; surfactant proteins A, B, C, and D; and also a myriad of innate immune products, including defensins, cathelicidins, lactoferrin, and also other inflammatory mediators such chemokines, interferons, and cytokines. Once infected with RSV, type II cells of term lambs have increased expression of SP-A, SP-D, and SBD-1; expression of these genes is reduced in RSV-infected type II cells of preterm lambs and may, in part, explain the increased susceptibility and disease severity of preterm lambs and infants to RSV infection. 61
Proteoglycans of the extracellular matrix can also regulate pulmonary inflammation and the innate immune response. Proteoglycans consist of a protein backbone attached to a glycosaminoglycan (GAG) side change. In the lung, GAGs include hyaluronan (14%), chondroitin sulfate/dermatan sulfate (31%), heparin sulfate (40%–60%)/heparin (5%), and keratan sulfate (<2%). 42 In the lung, the 3 families of extracellular matrix proteoglycans include large aggregating chondroitin sulfate proteoglycans (CSPGs), small leucine-rich CSPGs, and heparan sulfate proteoglycans (HSPGs). Perlecan, an HSPG, is present in the basal lamina; versican and decorin (CSPGs) are present in interstitial spaces; and syndecans are membrane proteoglycans. These proteoglycans and GAGs bind to numerous cytokines, chemocines, and growth factors, sequestering the molecules in a latent, inactive state. These include CXC chemokines (CXCL1-4, CXCL8, CXCL10, CXCL12), CC chemokines (CCL2-5, CCL11), proinflammatory cytokines (IL-1α and β, IL-2, IL-5, IL-6, IL-7, IL-12, TNF-α, interferon-γ [IFN-γ]), anti-inflammatory cytokines (IL-4, IL-10), and growth factors (fibroblast growth factors, vascular endothelial growth factor [VEGF], granulocyte macrophage colony-stimulating factor, and transforming growth factor [TGF]–β). 42 With inflammation and release of enzymes such as many of the matrix metalloproteinases, these inflammatory mediators can be released and become active, modifying the inflammatory response, including leukocyte adhesion molecules, and regulating both the innate and adaptive immune responses.
Further Triggering of Innate Immune Responses by RSV
If a virus such as RSV, another microbial agent or its product, an allergen, or foreign material surpasses the ASL and its contents, it can come in contact with the apical surface of epithelial cells of the lower airway. 98 Once epithelial cells are infected and damaged by RSV, the blood-gas exchange is compromised due to cell degeneration, altered cell physiologic activity, and altered airflow through the airways due to intraluminal accumulation of necrotic cell debris, mucin, vascular fluid, and airway constriction. Infected epithelial cells are not passively dependent on rescue by leukocytes, but the epithelial cells themselves have an active role in modulating the immune response as well as the release of anti-RSV compounds such as SP-A and inflammatory mediators and interferons that prevent viral replication. Such responses by epithelial cells are initially triggered by activation of pattern recognition receptors (PRRs), many of which are TLRs. TLRs are highly conserved molecules that recognize pathogen-associated molecular patterns (PAMPs) common to many general groups of pathogens and classes of microbes. PRRs activated by RSV include TLR-4, 47,64 TLR-2, 96 TLR-6, TLR-3, and retinoic acid–inducible gene I–like receptor (RIG-I). 83
TLR-4 is a cell-surface molecule associated with CD14 that recognizes LPS from Gram-negative bacteria and lipoteichoic acid from Gram-positive bacteria, as well as the F protein of RSV and other PRRs. 39,51,68,95 The F protein of RSV activates TLR-4 through binding of MD-2 protein, and this can be inhibited by lipid A analogue antagonists. 111 Binding of the TLR-4/CD14 complex activates NF-κβ, eventually leading to secretion of IL-8, IL-10, and IL-6, as well as increased expression of TLR-4 on epithelial cells. 39,68 There is scientific dispute over the impact of 2 TLR-4 single-nucleotide polymorphisms (SNPs) with regard to RSV disease severity. Multiple studies found that a single SNP or a haplotype with 2 SNPs in the TLR-4 gene is associated with increased susceptibility to symptomatic RSV infection and premature birth, 7,109 whereas other studies had contrary findings. 26,84,105 Two dissenting articles examined a single SNP as opposed to the 2-SNP haplotype. The third looked at immune response to RSV in 7- to 9-year-olds’ peripheral blood monocytes, as opposed to comparing epidemiologic RSV disease to the haplotype.
TLR-2 and TLR-6 form a cell-surface heterodimer. 95 Binding of TLR-2 activates NF-κβ through a MyD88-dependent pathway, initiating production of IL-1β. 119 Studies in knockout mice demonstrate TLR-2 and -6 signaling in leukocytes that stimulates an immune response to RSV. 96
TLR-3 is an intracellular TLR, present on the endosomal surface and recognizing double-stranded RNA (dsRNA). 95 RSV is a single-stranded RNA virus, but in vitro studies have demonstrated that TLR-3 is activated during replication (pairing of template and daughter strands) and leads to increased CCL-5 (regulated on activation normal T cells expressed and secreted [RANTES]) production. 116 Downstream signaling typically leads to IFN-α and IFN-β transcription. In lambs, TLR-3, 4, 7, and 8 are differentially expressed during fetal development. 133
RIG-I is an intracellular helicase that binds noncapped 5′ triphosphated RNAs and subsequently activates IFN regulatory factors 3 and 7 (IRF-3 and IRF-7), which are transcription factors for IFN-α and IFN-β. 95 Recently, nucleolin has been shown to be an important cellular receptor for RSV 136 F protein. After RSV enters lung epithelial cells, RIG-I is activated and upregulates CCL-5 and IFN-inducible protein (IP10) in addition to IFN-β. 83 Melanoma differentiation associated gene–5 (MDA-5) is also a helicase that recognizes dsRNA released from some viruses. Both RIG-I and MDA-5 have activated NF-κB, IRF-3, and IRF-7 via the mitochondrial antiviral signaling adaptor (MAVS), IFN-β promoter stimulator (IPS-1), virus-inducing signaling adaptor, and Cardif. Nucleotide-binding domain, leucine-rich repeat (NOD)–like receptors can also detect viral, bacterial, and other pathogens that enter the cytoplasm through the leucine-rich repeat domains. More than 20 NOD-like receptors have been identified. Some, such as NAPLP3, IPAF, and NALP1, activate the inflammasome in macrophages and other cell types, resulting in caspase 1 activation of IL-1β and IL-18. Endogenous molecules such as heat shock proteins 60 and 70, urates, and adenosine activate epithelial cells, leukocytes, endothelial cells, and other cell types in the lung. These endogenous substances are termed danger-associated molecular patterns (DAMPs) or alarmins. DAMPs activate inflammatory transcription factors; however, their signaling is modulated by simultaneous binding to CD24, which interacts with Siglec, a regulator of NF-κB. In fetal life, adenosine and other soluble factors have immunomodulatory activity and can skew cytokine responses through reducing production of IFN-γ and IL-12 and enhancing production of IL-1β, IL-6, and IL-23.
Many viral PRRs describe induce upregulation of type I interferons, IFN-α and IFN-β. Many cell types in lung produce type I interferons, but plasmacytoid dendritic cells (CD123+, CD303+, CD304+), which are present in lung, produce large amounts of IFN-α and lesser amounts of IFN-β after stimulation. Type I interferons reduce viral replication in cells, typically through JAK-STAT pathways, as well as by upregulating major histocompatibility complex (MHC) I, thereby indirectly promoting the killing of virus-infected cells, 48,49,67,95 and by producing downstream IFN products such as protein kinase R, which disrupts viral translation 107 ; 2′,5′-oligoadenylate synthetase 1 (OAS1), which activates an RNase L to cleave viral RNA 67 ; ISGylation, which inhibits viral replication 49 ; and MxA protein, which binds viral nucleoprotein to prevent replication and transcription. 48 Unlike other myxoviruses, RSV is resistant to MxA protein, although there are increased levels of MxA in the peripheral blood of RSV-infected children. 6,20,48 RSV’s nonstructural proteins NS1 and NS2 disrupt interferons through IRF-3. NS1 protein inhibits IFN transcription, and NS2 disrupts IFN signaling in the target cell. 135 A similar disruption of type I interferons by NS1 and NS2 working in concert at multiple points in the IFN pathway occurs in bovine RSV infection. 123 Since 2 of the 11 proteins of RSV aim to disable the interferon system, there may be therapeutic potential in counteracting the action of NS1 and NS2 to allow the immune system to better control RSV. RSV infection also decreases IFN-β (as well as IL-10 and TGF-β) in term lambs. 131 Type III interferons (IGN-λ) have been described and include IFN-λ 1, 2, and 3 (also termed IL-29, IL-28A, and IL-28B, respectively). These interferons signal through IL-10R2 and IFNLR1; however, their function is still being fully elucidated.
IFN-γ is a type II interferon and considered a Th1 cytokine, primarily produced by CD4+ T cells to activate macrophages, but in early RSV infection, it is produced by natural killer (NK) cells, dendritic cells, and macrophages. 95 TGF-β and IL-10 regulate IFN-γ by reducing IFN-γ expression and functional activity. 95 Such reductions in IFN-γ expression affect viral infection since IFN-γ inhibits viral replication and increases MHC I and MHC II cell surface expression as well as increasing transcription of MHC I–associated peptides involved in antigen processing. Also, stimulation of macrophages by IFN-γ induces secretion of IL-1, IL-6, IL-8, and TNF-α. 41 IFN-γ has been associated with increased CC10 protein, and there is evidence of its involvement in regulation at the transcriptional level. 113 Low IFN-γ in nasopharyngeal secretions has been associated with increased disease severity in RSV-infected infants. 127 This may indicate a skewing toward a Th2-type response as stimulation of immature lung macrophages with IFN-γ reduced RSV titers in mice. 31 IFN-γ is increased in term lambs infected with RSV and yet further increased in preterm lambs (over that of term lambs). 131,132
IL-1β, IL-6, and TNF-α are proinflammatory cytokines produced by macrophages and epithelial cells that act to increase acute-phase protein production from the liver, recruit neutrophils, and activate complement. A direct relationship has been demonstrated between IL-1β levels in pharyngeal secretions and severity of RSV disease. 82 Infection of healthy adults’ dendritic cells with RSV stimulates IL-1β production and secretion, as well as the proinflammatory cytokines TNF-α and IL-6. 45 High levels of TNF-α and IL-1β also are present in infants with RSV bronchiolitis. 46,88 TNF-α levels are increased in preterm lambs infected with RSV compared with term lambs. 131
IL-8 (CXCL-8) is a chemoattractant for neutrophils, basophils, and T cells that is secreted by monocytes, macrophages, fibroblasts, keratinocytes, and endothelial cells. 93 IL-8 is increased in nasal wash secretions of infants 1 to 6 months old infected with RSV, and this increase is more pronounced in those with more severe disease. 71 In the same study, IL-8 levels were correlated with an increased required duration of supplemental oxygen as well as higher peak fractional inspired oxygen. 71 In a study comparing term with preterm infants who experienced RSV infection before 6 months of age, a similar correlation between clinical severity score and IL-8 was found but not in preterm infants. 14 This highlights the complexity of understanding how the immune response contributes to disease in infants. As previously mentioned, signaling through TLR-4 via NF-κβ is one mechanism leading to increased IL-8. 68 IL-8 is increased significantly also in lambs infected with RSV and likely contributes to the neutrophil infiltration into bronchioles of RSV-infected lambs and infants. 131,132
IL-10 is primarily produced by leukocytes, particularly monocytes, and overall has a dampening effect on cellular recruitment through downregulation of class II MHC molecules and downregulation of Th1 cytokine production. 95 IL-10 is increased in nasal wash secretions of infants during RSV infection. 94 Intriguingly, elevated IL-10 during acute RSV infection is associated with postbronchiolitis wheeze. 126 Heterozygosity of an SNP in the IL-10 gene was associated with a decreased incidence of severe RSV but not associated with any difference in the incidence of postbronchiolitis wheeze. 56,126 IL-10 expression is decreased in term lambs infected with RSV. 132
TGF-β has a mixed effect on the immune response, activating neutrophils and typically inhibiting macrophage activation, although TGF-β does have the ability to activate or deactivate macrophages and monocytes depending on the rest of the cytokine milieu. 5,95 TGF-β is produced by NK cells and macrophages in innate immunity as well as T and B cells as part of the adaptive immune response. 54,134,140 Proliferation and differentiation of CD4+ T cells are blocked by TGF-β, which may be associated with a general dysregulation of Th1/Th2 response in infants. 138 TGF-β also plays a role in immunoglobulin production and class switching. 134 With RSV infection, TGF-β expression is decreased in term lambs. 132
Macrophage inflammatory protein–1 (MIP) is a 4-member family of CC chemokines: MIP-1α (CCL3), MIP-1β (CCL4), MIP-1δ (CCL9/10), and MIP-1γ (CCL15). 86 MIPs are expressed by lymphocytes, monocytes or macrophages, and epithelial cells and are generally proinflammatory, recruiting macrophages. 84 MIP-1α and MIP-1β are the most thoroughly studied and well characterized of the MIPs. Murine and in vitro studies show that MIP-1α expression is increased in RSV infection. 12,25,47,103 MIP1-α was increased in the nasopharyngeal secretions of infants with severe bronchiolitis 40 and also increased in preterm lambs infected with RSV. 131
Monocyte chemotactic protein (MCP-1/CCL-2) is produced by epithelial cells and functions as a chemoattractant for monocytes, eosinophils, and T cells. In vitro infection of human bronchiolar epithelial cells with RSV induces MCP-1 production. 103,104 In adults with upper respiratory infection due to RSV, MCP-1 is increased in nasal secretions during periods of viral shedding, 99 although no publications have indicated a similar finding in infants. MCP-1 is increased in term lambs infected with RSV and further increased in preterm lambs (beyond that of term lambs) infected with RSV. 131,132 Increased expression of MCP-1 may contribute to infiltration of neutrophils and other leukocytes into bronchioles and the lung.
Chemokines, such as RANTES (CCL-5), are chemotactic for T cells, dendritic cells, eosinophils, NK cells, mast cells, and basophils. Contrary to its name, RANTES is not limited to production by T cells; production of RANTES has been reported in platelets, macrophages, eosinophils, fibroblasts, and endothelial, epithelial, and endometrial cells. 78 Multiple studies on RSV severity and human polymorphisms in RANTES have yielded conflicting results, 1,50,145 although an interaction or compound effect of polymorphisms at multiple sites yielding specific haplotypes may explain those studies in which no significance was demonstrated. 50,55,58,93,137 A higher ratio of IL-1 to RANTES in nasopharyngeal secretions of infants with RSV was associated with a more severe clinical score, 53 and in another study, there was an inverse relationship between level of RANTES in tracheal secretions and markers of clinical disease. 120 In vitro recombinant RANTES inhibited RSV infection of HEp-2 cells. 30 Low levels of RANTES in RSV infection could be due to a predisposed deficit or defect in RANTES production (as may be indicated by the haplotype studies) or a direct blocking of RANTES by RSV, or it could indicate consumption during infection. RANTES is not significantly increased in term or preterm lambs infected with RSV, and PD-L1, a negative regulator of T-cell function through anergy and reduction of cytotoxicity, is increased in term lambs infected with RSV and further increased in preterm lambs infected with RSV (beyond that of term lambs). 131,132 Increased levels of CCL2, CCL3, CCL5, and CXCL10 are seen in infants with severe RSV bronchiolitis. 47,87
The cytokines and chemokines reviewed here are not exhaustive with regard to the full innate immune response of the lung to RSV but emphasize repeatable findings across multiple studies. Additional inflammatory mediators have potentially significant roles in innate defense to RSV infection; however, the current literature on their relationships to RSV is sparse. IL-17 stimulates neutrophil recruitment and stimulates fibroblasts and epithelial cells to secrete cytokines. 95 IL-17 was not detected among a population of healthy infants 1 to 6 months old, but it was detected in 15% of RSV-infected infants in the same age range. 71 IL-17 levels in nasopharyngeal secretions of infants hospitalized for RSV were increased at discharge compared with time of admission, suggesting that IL-17 may be involved with RSV convalescence. 33 In contrast, studies in mice studies suggest that IL-17 can affect regulatory T cells and contribute to increased lesions and inflammation. 24,46 IFN-γ–inducible protein 10 (IP10/CXCL10) is considered a marker of Th1 response and is increased during RSV infection, although its receptor, CXCR3, is decreased when compared with control subjects. 114 IL-19 and IL-20, which are related to IL-10 and induce TNF-α and IL-6, 95 are related to a postbronchiolitis wheeze similar to IL-10. 32 A polymorphism in IL-9 has an opposite effect in boys versus girls, associated with increased susceptibility in boys and protection in girls to severe RSV infection. 125
IL-21 is produced by activated CD4 T cells, natural killer T cells, T follicular helper cells, and Th17 cells. IL-21 induces production of inflammatory mediators from epithelial cells and fibroblasts and also mediates differentiation and activity of T, B, and NK cells and thereby restricts differentiation of regulatory T cells. IL-21 has little effect on primary RSV infection in mice, but in mice with RSV vaccine-enhanced disease, IL-21 reduced RSV disease severity and lung lesions, and expression of IFN-γ and IL-17 was associated with enhanced pathology. 24
Effector Cells of Pulmonary Innate Immunity and Effects of Age
The lung parenchyma has structural changes that occur during development and with age that influence innate immune responses and susceptibility to RSV and other pathogens. Premature infants and preterm lambs have increased RSV disease severity compared with healthy adults, and the reason for this is not fully elucidated but likely multifactorial. 23,59,66,70,90 The newborn has fewer alveoli that have increased wall thickness compared with adult alveoli. This reduces the efficiency of gaseous exchange and may contribute to more severe clinical symptoms. Submucosal glands are present in a primitive, unbranching structure in human respiratory airways at 13 weeks’ gestation and thereafter are present in more distal airways. 57 Submucosal gland expression and airway branching patterns of lambs are similar to those of infants. 23,122 The number and differentiation of Clara cells increase in the lung during fetal and perinatal development. 9,10 Clara cell CC10 protein is expressed as early as 10 weeks’ gestation in humans 63 and consistently by 15 weeks’ gestation. 9 Type II cells increase in number and further differentiate with fetal age in human and lambs. 89,108
The vascular endothelium serves a pivotal role in the innate-adaptive immunity interface, responding to innate signals to recruit and adhere to neutrophils, monocytes, dendritic cells, NK and NK T cells, eosinophils, basophils, and lymphocytes and also regulate the permeability of the vascular system to allow leakage of serum. Serum within airways, alveoli, and the pulmonary interstitum can dilute microbial agents and provide additional antimicrobial factors such as complement, antibodies, and collectins. There is a significant correlation between clinical severity score and total nasal wash leukocyte counts in full-term but not preterm infants. 4
Neutrophil infiltration into the lung can occur rapidly in response to certain stimuli and release of inflammatory mediators and chemokines such as IL-8. Neutrophil infiltration is a key feature of RSV-induced bronchiolitis (Fig. 3). Neutrophils of neonates have a reduced proliferation pool and storage pool, and neutrophils in circulation have an impaired response to chemotaxins, including reduced rolling adhesion, transmigration, and lamellipodia formation. Neutrophils of infants have reduced function at the site of infection compared with adults. 79 Neutrophils express nearly half the level of lactoferrin and 70% of BPI, and they have impaired oxidase activity. 2,80,110 Preterm lambs have reduced levels of myeloperoxidase. 131
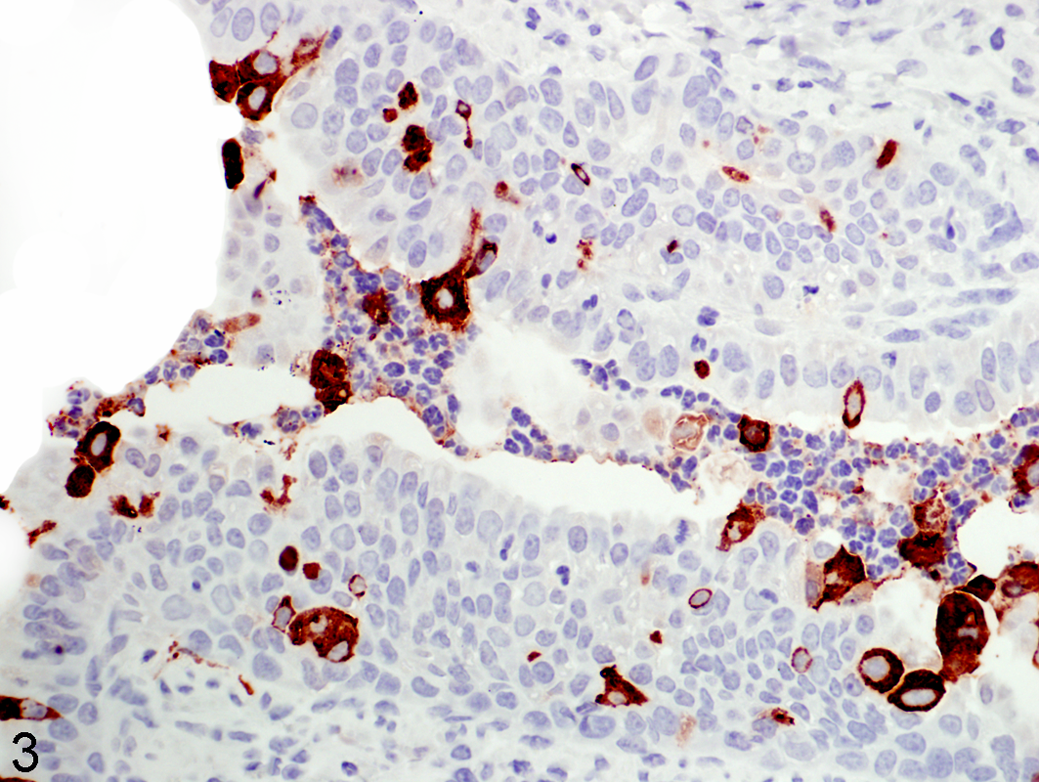
Lung, lamb. Lung airway depicting key features of respiratory syncytial virus (RSV) infection of the newborn lung. Neutrophils are present in the airway lumen infected with RSV, a human strain Memphis 37. Within the airway lumen are numerous neutrophils that are admixed with detached epithelial cells, many of which contain RSV antigen. RSV antigen is also present within multifocal epithelial cells lining the airway; neutrophils lack RSV antigen. Immunohistochemistry stain to RSV antigen detected by primary polyclonal goat anti-RSV (all antigens) antibody (EMD/Millipore/Chemicon, Billerica, MA); hematoxylin counterstain. ×40.
Classical monocytes (CD14+, CD16–) can enter the lung to differentiate into alveolar macrophages, whereas non-classical monocytes (CD14lowCD16–) patrol along the vascular lumen for antigen uptake and presentation and subsequent recruitment of neutrophils and classical monocytes. Following exposure to RSV, monocytes from neonates have a more limited response than adults, suggesting a reduced level of adaptive immune response ability in the neonate. 66 In vitro stimulation of neonatal monocytes and antigen-presenting cells results in decreased expression of TNF-α, IFN-α, IFN-γ, IL-12, and IL-1β but increased expression of IL-6, IL-8, and IL-10. 79 Monocytes entering the lung can differentiate into pulmonary alveolar macrophages (PAMs) that are thereby well positioned to interact with inhaled substances. Although adult mice infected with RSV have a robust response by classically activated alveolar macrophages, responses are reduced in neonatal mice. 31 Stimulation of neonatal mouse alveolar macrophages reduces RSV titers and enhanced weight gain. 31 Alveolar macrophages of preterm lambs have reduced levels of nitric oxide compared with term lambs. 131 Certain species, including cattle, horse, pig, sheep, goat, cats, and whales, have significant numbers of pulmonary intravascular macrophages (PIMs). PIMs are not readily detected in human lung; however, they may be detected in lungs of humans with certain disease conditions such as hepatopulmonary syndrome. 124
The 2 major classes of dendritic cells (DCs) are myeloid DCs (mDCs), which express CD11c+, and plasmacytoid DCs (pDCs), which are CD11c–. As indicated, pDCs produce type I IFN, especially IFN-α with lesser amounts of IFN-β; however, IFN production by pDCs is less in neonates than in adults. mDCs include mDC1 (CD1c+CD141–) and mDC2 (CD1c–CD141+). All 3—mDC1, mDC2, and pDC—are infected by RSV and undergo maturation and cell-specific cytokine production; however, pDCs are infected at a lower level than mDC1 and mDC2. 60 DCs in neonates are reduced in number and distribution compared with adults, and the ratio of myeloid to plasmacytoid DCs is inversed as compared with adults. Neonatal DCs have a reduced ability to produce IFN and decreased ability to stimulate a Th1 response. 143 Dendritic cell responses of neonates to RSV have reduced activity compared with adults. RSV-infected DCs co-cultured with T cells of either adults or umbilical cord blood elicited markedly different cytokine profiles, with the primary differences attributed to differences in response to TGF-β. 138 In mice, neonatal lungs have a deficit in conventional and plasmacytoid dendritic cells along with a shift of cytokines and transcription factors toward Th2 responses. 115 Furthermore, infecting neonatal mice with RSV results in enhanced TNF-α initially followed by increased IL-13, mucus hyperproduction, and airway hyperreactivity. 144 Pulmonary DCs of mice include CD103+, which localizes along the epithelium, as well as CD11b+ and monocyte-derived DC (immature phenotypically), which localize in the interstitium. Pulmonary dendritic cells isolated from term lamb lung differ from those isolated from adult lung in terms of antigen expression and maturation. 34 –36 Ovine pulmonary dendritic cells from term lung support bovine RSV replication and have enhanced IL-4 and IL-10 gene transcripts. 34 –36
In utero, the fetal lung is not exposed to airborne antigens; however, some noninherited maternal antigens cross the placenta and can trigger regulatory T cells, which can constitute up to 15% of the peripheral T cells in the fetus. After birth, RSV and other viral infections and inhaled antigens stimulate infiltration of CD4+ and CD8+, which, along with DCs and other cell types, influence Th1 and Th2 cytokine responses, which are well studied by RSV immunologists. Formalin-inactivated RSV vaccines also enhance RSV disease in humans and many animal models. IL-21 production in vaccine-enhanced RSV disease, for example, can affect regulatory T cells, reduce IL-17, and, in this way, control inflammatory responses within bronchioles, the site of RSV infection and lesion development. 24 These are adaptive responses reviewed elsewhere and beyond the scope of this review of pulmonary innate immunity. 21,46 However, there is considerable overlap between the innate and adaptive immune systems in contributing and regulating T-helper cell responses as a number of the innate factors are integral to priming and directing the adaptive response. Advances in understanding innate immunity will likely contribute to more precise and effective adaptive responses as well as create a more complete picture of RSV disease and immunity.
Discussion/Conclusion
From the above review, it is clear that the innate immune response is important in RSV infection, and a better understanding of it, as well as how to modulate it, could make a significant contribution to RSV prevention and therapy in infants. Enhancement of innate immune responses may reduce RSV disease severity or prevent initial infection. Also, because many innate immune responses invoke adaptive immune responses, enhanced activation of innate immune responses may bolster/strengthen adaptive responses. Innate immune responses require some level of lung maturation and activity of single cell types in some cases (eg, surfactant protein A production by type II cells) or several cell types (epithelial and submucosal glands for Duox/LPO oxidative defense). 37,38 Optimal lung maturation is needed for an effective innate immune response, and studies in lambs have demonstrated that VEGF administration prior to RSV infection reduces disease severity. 92,102 VEGF expression by development lung is vital for maturation and differentiation, and exogenously administered VEGF upregulates SP-A, induces monocyte infiltration, and can contribute to vascular leak. 102 Therapies such as VEGF may activate key aspects of the innate immune response and simultaneously trigger other defense mechanisms with potential enhancement of adaptive responses. There are also innovative therapeutic approaches, including genetic vaccines; microparticles; therapies targeting the RSV receptor; L-polymerase inhibitors; therapies that promote enhancement of TLR-4 signaling intensity; therapies that target the recently discovered RSV receptor, nucleolin; and many others, giving hope for effective prophylactic and therapeutic strategies.
Despite what is known about pulmonary innate immune responses to viruses such as RSV by the neonatal lung, there is much to be learned. Many cellular mechanisms of innate immune products have yet to be fully defined in the newborn respiratory tract, including inhibitory RNA regulation of innate immune gene expression, expression and downregulation of innate immune gene receptors by the various cell types, the extent to which immature or poorly differentiated cells can produce innate immune products, and the extent to which sequestration of inflammatory and innate immune mediators by extracellular matrix proteoglycans differs in the newborn versus the adult respiratory tract.
Footnotes
Acknowledgements
We thank Dr Albert Van Geelen, Jack Gallup, and the many others who have contributed to the studies.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was funded in part by NIH/NIAID 1R56AI091000-01A1 and NIH/NIAID R01AI062787, Gilead Sciences, Meridian Bioscience, and MedImmune, LLC.