
Abstract
A colony of golden hamsters had an ongoing problem with hydrocephalus. In an attempt to clear the colony of the problem, new breeders from another supplier had been purchased. At termination of a behavioral study, the brain was collected from 35 animals (four of which had died with hydrocephalus during the study) and was examined macroscopically and by light microscopy. Although no animals manifested obvious behavioral changes, 31 of 35 (88.6%, 13/15 males and 18/20 females in control and manipulated groups) had hydrocephalus. Twenty-five animals had macroscopically identifiable hydrocephalus, and six had hydrocephalus identified microscopically. Neither teratogenic concentrations of metals nor mycotoxins were detected in tissues or food, and sera from breeders tested negative for antibodies to Sendai virus, reovirus 3, and lymphocytic choriomeningitis virus. Trial matings of breeders expected to produce hydrocephalic offspring resulted in affected offspring, and mating of breeders expected to produce normal offspring resulted in normal or less-affected offspring. Hydrocephalus was confirmed retrospectively in some breeders. Hereditary hydrocephalus appears to be widespread in hamster stocks in Central Europe. Affected animals do not manifest signs of disease and usually die without obvious premonitory signs. Despite severe hydrocephalus, the animals can breed, and animal handlers do not identify motor deficits or abnormal behavioral activity. This entity is unlike the previously described, hereditary hydrocephalus of hamsters that is phenotypically identifiable and usually is lethal before they attain breeding age.
Hydrocephalus is an accumulation of cerebrospinal fluid (CSF) that is the result of an imbalance between its synthesis and absorption in the central nervous system (CNS). 12, 16 With few exceptions, it is the result of increased resistance to absorption. Often, it is the result of obstruction to CSF flow within the ventricular system: noncommunicating or obstructive hydrocephalus. Hydrocephalus can be of a communicating type when meningitis, subarachnoidal neoplasia, or trauma external to the ventricular system increases resistance to CSF flow. Hydrocephalus may be congenital or acquired. In humans, where it is diagnosed in almost one of every 1,000 births, most causes of congenital hydrocephalus are unknown. Hydrocephalus is a common malformation and has been described in several domestic and laboratory animal species. 16, 17 Although the condition may be hereditary, a number of infectious, chemical, and physical agents have been documented to cause hydrocephalus. The golden hamster (Mesocricetus auratus) not only has a heritable form of hydrocephalus, but also was one of the first laboratory animal species studied for experimental induction of hydrocephalus via intracerebral inoculation with several pathogens. 5, 8, 10, 11, 13, 14, 18, 19, 21 Additionally, in utero exposure to several metals and chemicals has been associated with hydrocephalus in hamster offspring. 1, 6, 7
A university pathology diagnostic service was contacted to evaluate an ongoing problem of hydrocephalus in the university's hamster colony. Over a year-long period, sporadic cases of hydrocephalus had been diagnosed in animals of the colony that died. Despite buying new breeding stock from a second, commercial supplier, cases of hydrocephalus continued to be detected. We describe the findings of a study of spontaneous, hereditary hydrocephalus in golden hamsters.
Materials and Methods
All observations were done on golden hamsters raised in the Institute of Animal Genetics, Nutrition and Housing, Division of Animal Housing and Welfare, Bern, Switzerland. Hamsters were reared using standard practices, and all handling followed guidelines approved by the Institution's Animal Welfare Committee.
When a pathologist became involved for a more in-depth study of the problem, a group of 31 (17 female, 14 male) golden hamsters in a study were scheduled for termination. These animals had been selected from nine litters. The remaining littermates of these nine litters had been used in other studies and were not available for examination. Four hamsters (one male and three females) had already died acutely of hydrocephalus during the experiment. The parents of the experimental animals were derived from animals purchased from two suppliers.
The hamsters had been weaned and placed on a 12-week, behavioral experiment. For that experiment, they had been housed singly in cages of various sizes and with a variable depth of litter to observe the effect of these variables on behavior. Animals were videotaped principally during their nocturnal, active phase, and their time spent in various behaviors was monitored and quantified (time spent chewing, grooming, climbing). Their running activity on an in-the-cage, running wheel was continuously monitored. They were euthanized when they were 15 weeks old, and were examined for macroscopic lesions. The brain and samples of liver, lungs, kidneys, and heart were fixed in buffered formalin and processed in routine manner for light microscopic examination of 5 µm-thick, HE-stained sections of paraffin-embedded tissues. One section of each visceral organ and four coronal sections of each brain were examined. Brain coronal sections were at the level of the cranial portion of the caudate nucleus, the nucleus habenularis, the rostral colliculus, and the cerebellar roof nuclei. Brains were graded as normal, mildly hydrocephalic (if lesions were barely visible macroscopically or only noted histologically), or severely hydrocephalic (if lesions were macroscopically obvious).
One male animal in the colony was observed to be smaller than all males of the same age. It was anesthetized (inhalation isoflurane), and the brain was examined by use of ultrasonography (Aloka Prosound SSD 5500, Zug, Switzerland). This male was found to have hydrocephalus and was used for later breeding studies. Another 30-day-old male in the colony was observed to have poor balance and possible doming of the skull. Prior to euthanasia, it was anesthetized (ketamine hydrochloride [1 mg/kg of body weight] and medetomine [0.15 mg/kg]). After confirming that the animal had hydrocephalus by use of magnetic resonance imaging (MRI; Hitachi AIRIS II, Tokyo, Japan), One-hundred microliters of gadodiamid (Omniscan; 0.005 mmol/ml) was inoculated into the right lateral ventricle, and the MRI procedure took 30 minutes to complete. Sequences performed included: a T2-weighted fast spin echo in transverse orientation, a dorsal, completely balanced, steady-state, free precision type sequence, and a dorsal T1-weighted, field echo plane after intraventricular contrast application.
Sera from two dams of the litters, one clinically normal dam and one with severe hydrocephalus, were tested for antibodies to reovirus 3, lymphocytic choriomeningitis virus (LCMV), and Sendai virus (MICROBIOS Gmbh, Münchenstein, Switzerland). Liver samples from the same two animals were tested for concentrations of Cu, Cr, Zn, Pb, Cd, and Hg, and feed was assayed for fumonisin and ochratoxin A (Interlabor Belp Ag, Belp, Switzerland).
Parents of the animals in the behavior experiment were still alive, but resources to examine all animals by ultrasonography or MRI were not available. Assuming that the problem was heritable, several matings were done to help elucidate transmission of the hydrocephalic trait. Results of necropsy of the original 35 experimental animals were used to predict the lesion status of the parents. The following matings were done (Table 1): mating A, repeat mating of two hamsters that produced affected offspring in the original experiment (the female ultimately was found to be severely hydrocephalic and the male was mildly hydrocephalic); mating B, mating of two hamsters that had produced clinically normal offspring in the original experiment (both breeders ultimately were found to be normal); mating C, severely affected male (diagnosed by ultrasonography) bred to a mildly affected female; and mating D, the known, severely affected male of mating C bred to a severely hydrocephalic female. These mated females delivered their offspring, which were euthanatized and examined at weaning. Permission was given to euthanize all parents mated to produce the litters of hamsters used for the behavioral experiment, and the brain and viscera from breeders and offspring were examined as described previously.
Hamster test matings to study hereditary hydrocephalus.

= severe hydrocephalus, macroscopically obvious;
= not affected;
= mild hydrocephalus, microscopically obvious;
= sire diagnosed by ultrasonography prior to mating;
= unknown status; live animals not examined.
Results
During the behavioral experiment, none of the animals, including the four that died before the scheduled termination date, manifested obvious clinical signs of disease. Phenotypically, none of the 35 experimental animals had domed skull or were identified as lagging behind littermates in growth rate. Of the 35 experimental hamsters, 25 (71.4%, 12/15 males and 13/20 females) had severe, macroscopically visible hydrocephalus. However, 31 of 35 (88.6%, 13/15 males and 18/20 females) had hydrocephalus macroscopically visible or histologically detectable (Fig. 1). Four animals had asymmetrically affected lateral ventricles (Fig. 2).
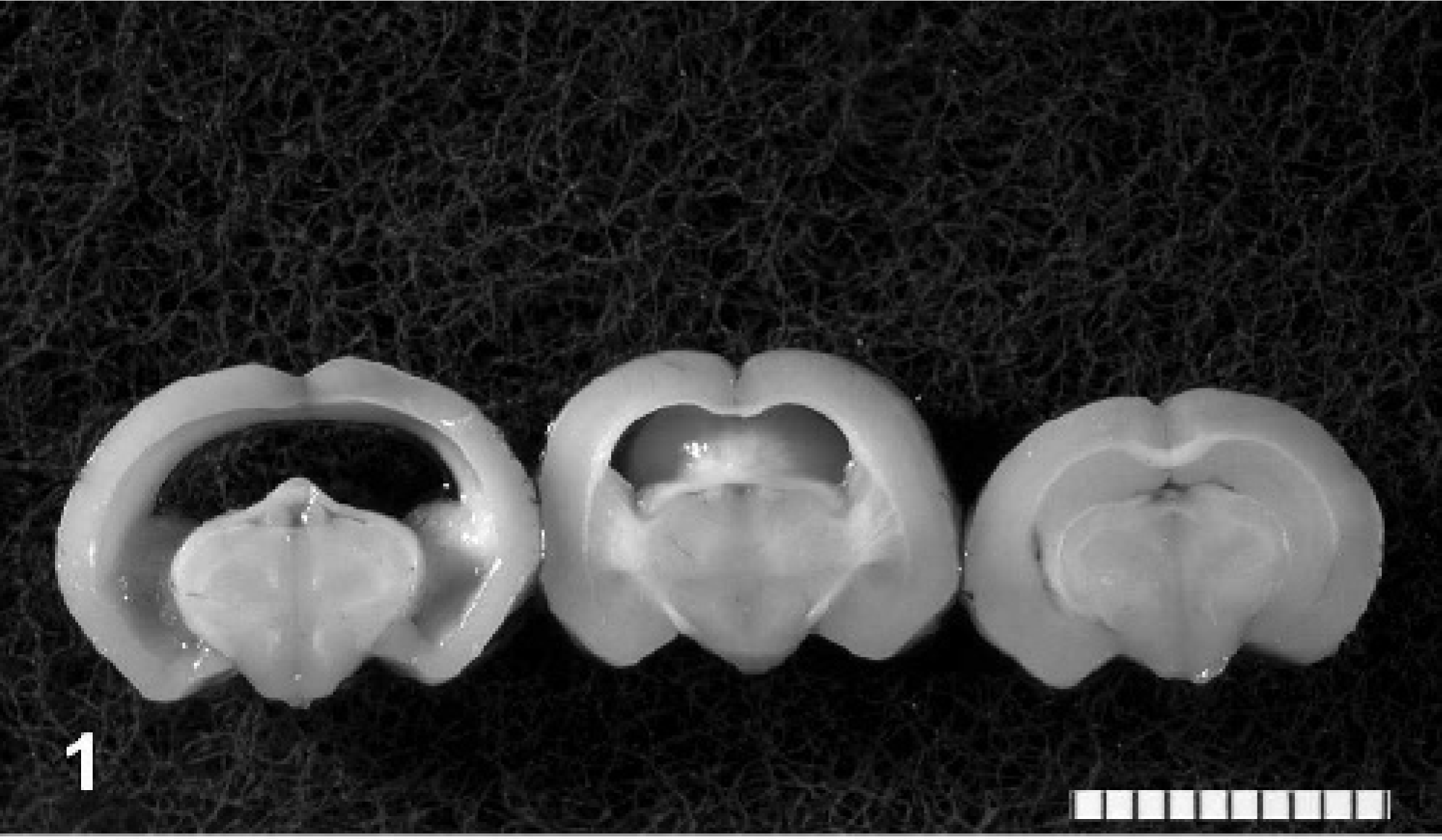
Brain; hamster. Cross sections from three hamsters with differing degrees of hydrocephalus. The section on the right has barely perceptible, macroscopic hydrocephalus. Bar = 1 cm.
It was determined histologically that only the lateral ventricles were dilated. In affected animals, the periventricular parenchyma was rarified subjacent to broken ependymal linings, but inflammation, periventricular diverticulae or clefts, necrosis, hemosiderosis, vascular abnormalities, and gitter cells were not observed. The ventricular lining was intact in most areas, but segments of ependymal cells were attenuated or missing (Fig. 3). The choroid plexuses were judged not to be atrophic. Histologic examination of sections did not reveal overdistention, absence, or obstruction of the third or fourth ventricular system or the mesencephalic aqueduct. Usually, the lining of the nondistended mesencephalic aqueduct was folded, and the aqueduct profiles were often flat or star-shaped (Fig. 4). Because the brain lacked obstructive lesions and the third and fourth ventricles were not dilated, it was concluded that these hamsters had a communicating hydrocephalus. With the exception of one male with a urolith and hematuria, other lesions were not seen macroscopically or microscopically in visceral organs.
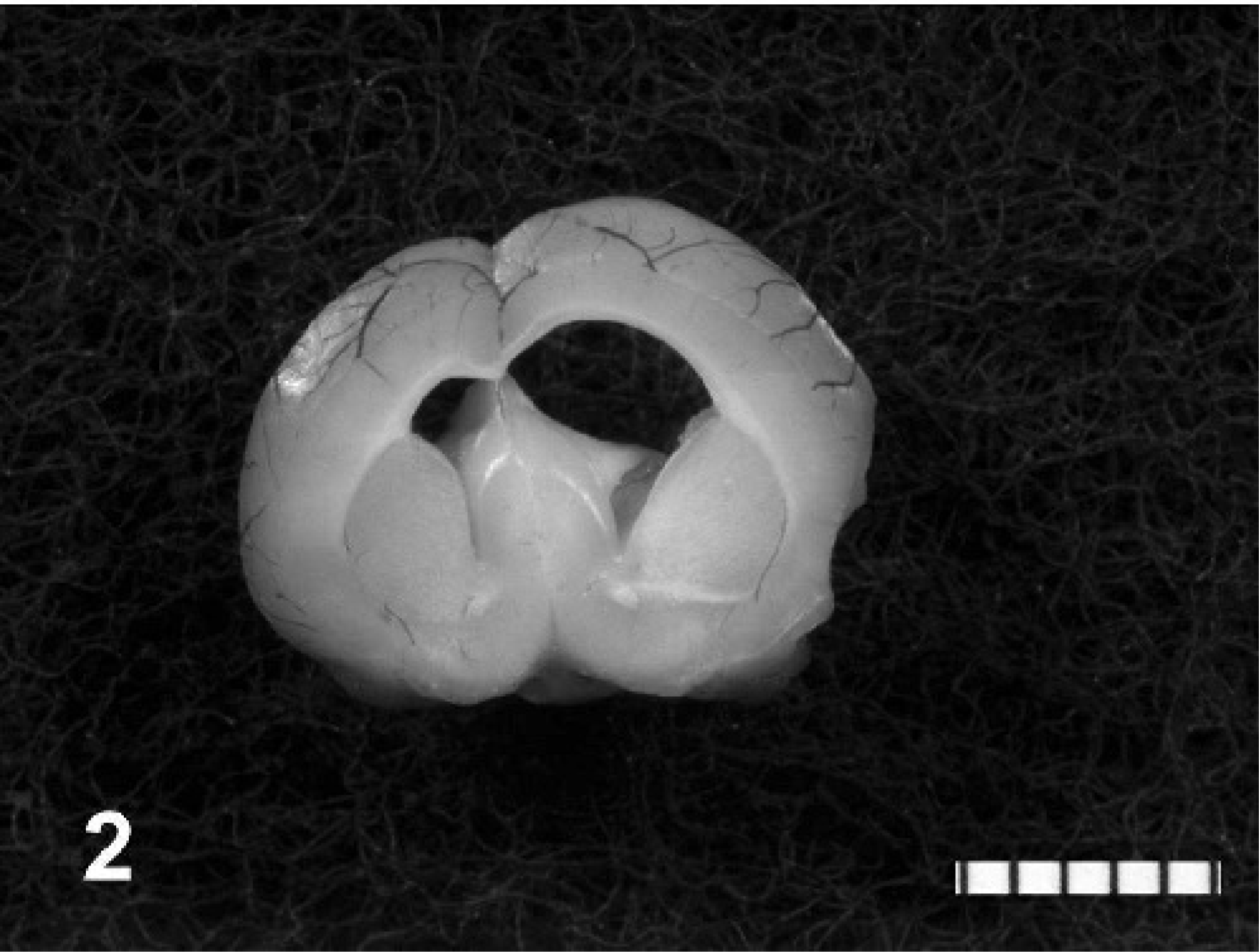
Brain; hamster. Hydrocephalus of the lateral ventricles is asymmetric. Bar = 0.5 cm.

Cerebral cortex; hydrocephalic hamster.

Brain, mesencephalic aqueduct profiles; hamster.

Litter of hamsters from the test mating of two hydrocephalic hamsters. One (center) of five animals, all of which had hydrocephalus, is stunted and has a dome-shaped skull.
A small male in the colony was observed and, using ultrasonography, severe hydrocephalus was detected. This male recovered from anesthesia unremarkably and was used in subsequent breeding studies before it was euthanized, after which severe hydrocephalus was confirmed by necropsy and histologic examination. Another male in the colony with mild ataxia was observed to have a slightly domed skull, and hydrocephalus was diagnosed by use of MRI. Gadodiamid injected into the right ventricle was observed to travel from the right lateral ventricle into the left ventricle, down the third ventricle and aqueduct, and into the fourth ventricle. This hamster was euthanized after the injection study, and severe hydrocephalus typical of that in other hamsters of the colony was observed.
Remating of two parents (severely affected female and mildly affected male), the previous mating of which had produced severely affected animals used in the behavior experiment (Table 1, mating A), resulted in a litter of five pups, all with severe hydrocephalus. One male of this litter was stunted and had a domed skull (Fig. 5). Mating of two clinically normal parents that had previously produced some clinically normal offspring used in the behavior experiment (mating B) produced a litter of three clinically normal pups and one pup with histologically detectable hydrocephalus. The mating of the ultrasonographically diagnosed affected male with a female with mild hydrocephalus (mating C) resulted in a litter of eight pups, one with severe and seven with mild hydrocephalus. Mating of the same affected male with a severely affected female (mating D) resulted in six offspring, four of which had severe hydrocephalus. The other two offspring of this litter were retained for future study, and although not examined to date, they have produced severely affected offspring. Affected animals of either sex were produced from all matings of affected parents.
The 35 experimental animals were selected from nine litters. Examination of the parents of the experimental group indicated that three of nine dams had hydrocephalus (one mild and two severe), and that three of five sires had mild hydrocephalus. Four matings of phenotypically normal parents had resulted in only two clinically normal offspring and 11 affected animals (seven of which were severely affected). Mating of mildly affected animals to clinically normal animals had resulted in two clinically normal and eight affected offspring. One litter from a mildly affected male mated to a severely affected female resulted in only severely affected offspring. Severely affected animals had not been mated to each other to provide animals for the original experiment. Unfortunately, not all offspring from the initial nine litters were used in this study and were available for study.
Antiviral antibodies to reovirus 3, Sendai virus, or LCMV were not detected in sera, and Zn, Cu, Cd, Pb, Cr, and Hg were not detected in toxic amounts in the liver samples. The tested food was negative for ochratoxin A and fumonisin.
Discussion
The lesions do not provide evidence of an obvious cause or pathogenesis for this form of hydrocephalus. When examined, the process in the brain appeared inactive without inflammation or any of the lesions associated with chronic hydrodrocephalus. The lack of doming in most affected animals suggests that the condition may have developed after suture lines closed, 16 and the dilatation was limited to the lateral ventricles, suggesting that the condition was the result of aqueductal stenosis. The mesencephalic aqueduct (of “Sylvius”) is the site obstructed in more than two thirds of human cases of noncommunicating, congenital hydrocephalus, 12, 16 but blockage, focal gliosis, or forking of the aqueduct was not evident in these hamsters. The closed, folded, or star-shaped aqueduct profiles might be interpreted as collapse of aqueducts that were dilated in the live animal. However, aqueducts of nonperfused brains tend to appear slightly more collapsed than those of perfused brains. Even normal hamster brains in this study had folded aqueduct linings. Still, it might be that the folded lining represents a form of dysplasia of variable severity in these related hamsters, and only when it is sufficiently severe does it lead to obstruction and hydrocephalus.
Several viruses can induce hydrocephalus in hamsters; however, to experimentally induce hydrocephalus, the viruses are inoculated intracerebrally into newborn animals. 5, 8, 11, 13, 14, 18 Hydrocephalus can be induced rapidly in neonatal hamsters, but mature hamsters usually are not susceptible to viral-induced hydrocephalus. 13, 18 Animals with virus-induced hydrocephalus often develop domed skulls, manifest neurologic impairment, and commonly are physically stunted. Hamsters that recover from viral infection may maintain motor deficits, and depending on the virus, have residual histologic lesions such as periventricular gliosis, spongiosis, inflammation, or hemosiderosis. None of these lesions were present in the hamsters of this study; rather, the only lesion noted was patchy, multifocal disruption or attenuation of the ependymal lining, with subjacent neuropil edema. Because ependymal cells replicate little if any after birth, any process causing necrosis or tearing of the ependyma could result in the lesions seen in the ependymal lining. 2, 4, 15, 20
Support for an infectious cause would have come from the demonstration of circulating antibodies to viral pathogens that cause hydrocephalus in hamsters. Although only two animals were tested, viral antibodies were not detected in their serum. Usually, circulating antibodies in recovered breeders would be expected to protect the fetus by preventing viremia in the parents, providing lactogenic immunity in colostrum, and preventing shedding of virus to newborn pups. Although LCMV can persist in recovered hosts, there was no serologic or histologic evidence of LCMV in members of the colony. Ultimately, it was concluded that the hydrocephalus was not induced by an infectious agent.
A toxic cause of hydrocephalus was considered. Some toxins and metals can induce hydrocephalus if the developing fetus is exposed in utero, and we tested for several of these teratogens. 6, 7, 12 Although, only two liver samples were tested, metals associated with in utero induction of hydrocephalus in hamsters and rats were not detected. Food likewise tested negative for fungal toxins that can cause hydrocephalus in hamsters.
Evidence that this is a hereditary condition came from the results of test matings. The litters from these test matings were small, presumably because of the advancing age of the breeders and not because of fetal death. Initial litters from which experimental pups had been selected were of normal size for the colony (data not shown). The reproduction of a litter with severe hydrocephalus using affected breeders (matings A and D) was strong evidence that the problem was not infectious. Despite exposure to the same housing conditions, feed, and water, reduction in the occurrence and severity of lesions resulted from mating normal parents (mating B) in the same colony. A genetic basis for the condition was further supported because reduced severity of the lesions resulted from mating of less affected animals (mating C). Analysis of all of the data of this study does not permit conclusions regarding mode of inheritance or the presence or absence of a major locus or single gene controlling the condition. Information regarding the parents of the original breeding animals and the phenotype of all siblings in the litters was not available. More generations of animals and more specific backcrosses are needed to characterize the pattern of inheritance. This heritable problem is not sex associated and most probably is an autosomal recessive trait. Such a defect with variable penetrance would explain why breeding phenotypically normal animals had resulted in many severely as well as mildly affected offspring. Because breeding phenotypically normal animals may result in affected offspring, the condition will be difficult to detect in colonies.
Significant differences are apparent between the previously reported, hereditary hydrocephalus described in golden hamsters and the condition observed in the hamsters of this study. 21 In the published descriptions of hereditary hamster hydrocephalus, all hydrocephalic hamsters were stunted and had a characteristic domed skull. Those animals usually did not survive to breeding age. For the most part, the external phenotype of the hamsters of the present study was normal, and all animals lived to breed successfully. Additionally, the three animals identified as affected because of slower growth, domed skull, or balance problems had better survivability than those with the published form of hereditary hydrocephalus. The histologic lesions of the described, hereditary, hamster hydrocephalus have not been published; thus, further comparison is not possible.
The condition resembles the H-Tx hydrocephalus mutation in rats. 3, 9 The H-Tx mutation only affects the lateral ventricles and is not associated with any lesion that suggests aqueduct stenosis or malformation. However, unlike our hamster condition, affected H-Tx rats routinely develop domed skull, and affected animals do not thrive. Most affected H-Tx rats die within 14 weeks after birth. The histologic lesions of both conditions are similar. Similar to the hamster condition, H-Tx is not sex linked and backcrosses of normal animals result in affected offspring. The analysis of the inheritance of the H-Tx mutant suggested that H-Tx is a monogenic, recessive, autosomal condition with incomplete penetrance.
These animals' ability to thrive and perform normal motor tasks during the behavioral study despite severe loss of cerebral tissue was surprising. In rats with kaolin-induced (K-I) hydrocephalus, there appears to be a threshold of ventricle size only beyond which functional changes are detected. 4 In K-I hydrocephalus in hamsters, clinically observed, neurologic deficits are seen immediately after kaolin injection and persist for at least 15 days even though the kaolin causes less dilatation of ventricles and cortical loss, compared with that in the hamsters of the present study. 1 Of course, in the K-I hydrocephalus model, kaolin is injected into the brain stem and would affect different areas of the brain.
More work on antemortem diagnosis of this heritable hydrocephalus using non-invasive techniques is warranted. The condition may remain latent in a colony because many hydrocephalic animals result from breeding of clinically normal parents. Genetic markers would facilitate identifying carriers. If the colony experience of the present study is typical, one may expect occasional deaths in affected colonies, and if necropsy is performed on unscheduled deaths in a colony, the condition could be detected. The fact that affected animals or carriers were provided by two commercial suppliers suggests that the condition is widespread. Hydrocephalus is reported sporadically in hamsters. One study of a movement disorder of hamsters (the dstz mutant) initially associated the signs of the disease with hydrocephalus; however, the researchers discovered they had hydrocephalus in their control animals as well. 19 Perhaps, the conclusions of past studies using hamsters and having hydrocephalus as an endpoint should be reexamined in light of the use of adequate controls and the incidence of spontaneous hydrocephalus.
At the time of this study, funding was not available to investigate the problem further. Permission to examine the breeders provided important information, but the pathogenesis of the lesion remains uncharacterized. By the time the lesions were examined (after weaning), they appeared fully developed and quiescent, and examination at various points in the genesis of the changes would permit better characterization of the process. It is presumed that the remainder of the ventricular system of these hamsters is patent, at least partially. However, the study of one injected animal and our histologic examination done at one point in time late in the progression of the process do not definitively establish aqueduct patency throughout brain development. Additionally, the one animal injected was phenotypically different (domed skull) from the more typical hamsters with this condition. Hydrocephalus in golden hamsters remains an important problem about which researchers should be aware. The incidence of hydrocephalus in hamster colonies should be established. This form of hydrocephalus presents an opportunity that could be manipulated in the laboratory to study hydrocephalus. Given the extent of the condition at this time in suppliers' breeding stock, a breeding colony of affected animals may still be generated by examining animals in colonies and sharing of information among breeders.
Footnotes
Acknowledgements
We would like to thank Dr. Thomas Famula of the Department of Animal Science, University of California at Davis for performing statistical genetics analysis.