
Abstract
A massive fish kill affecting exclusively common carp (Cyprinus carpio carpio) in the St. Lawrence River, Québec, Canada, during the summer of 2001 was investigated by use of laboratory diagnostic methods and by an attempt to experimentally induce the disease. The ultimate causes of mortality were opportunistic bacterial infections with Aeromonas hydrophila and Flavobacterium sp. secondary to immunosuppression induced by physiologic (i.e., spawning) and environmental (i.e., high temperatures and low water levels) stressors, and possibly enhanced by an infection causing lymphocytic encephalitis observed in 9 of 18 (50%) fish examined. Experimental induction of disease was attempted in captured wild carp by administration of crude and filtered (particulate <0.22 μm) inocula prepared from a homogenate of tissues from carp affected by the natural outbreak. Although significant clinical disease or mortality was not induced by experimental challenge, lymphocytic encephalitis similar to the one observed in naturally affected carp was induced in four of seven (57%) fish administered crude inoculum and four of seven (57%) fish administered filtered inoculum. None of the control fish inoculated with sterile phosphate-buffered saline (n = 6) were affected by encephalitis. The cause of the encephalitis observed in carp from the natural outbreak and in experimentally inoculated fish could not be determined by use of virus isolation and transmission electron microscopy.
Every year, North American freshwater bodies are affected by fish kills, which can result in mortality of a few hundred to several hundred thousand wild fish. 13 Fish kills can be a manifestation of environmental disturbances that are substantial for ecosystem and human health. These events often cause considerable concern among the public, and they can have economic consequences on the angling, commercial fishing, and aquaculture industries. Fish mortality can involve infectious diseases that have implications for national and international trade. 2 Therefore, determination of the cause of wild fish kills by field and laboratory investigations is important. Fish die-offs can be caused by a wide variety of human-related and natural factors, such as toxic spills caused by agricultural and industrial operations, climatic factors, and infectious diseases. In many instances, diagnostic investigation is complicated by the multifactorial nature of these events.
In the summer of 2001, one of the most important fish kills in recent Canadian history occurred in the St. Lawrence River, Québec, Canada. Epidemiologic characteristics of this mortality and results of field investigation were previously reported. 15 From 28 June to 31 July, dead common carp (Cyprinus carpio carpio) were observed at numerous locations on both shores of the St. Lawrence River and Saint-Pierre Lake, from Lac des Deux-Montagnes to Montmagny (approx. 250-km distance). During that period, 25,964 dead and moribund carp were removed from selected inhabited areas along the shores, with a peak of carcass removal 2 weeks after the onset of mortality. The actual number of carp affected was probably higher than 25,964, as this count of dead fish was obtained from a small portion of the affected territory. Mortality affected exclusively common carp and targeted predominantly spawning adults.
The common carp is a robust freshwater fish that is widespread in Canada and most parts of the world. 19 Although this fish is of minor economic importance in Canada, carp aquaculture for human consumption is a major industry in Europe and Asia. In 2002, the worldwide production of cultured carp was 3.2 million metric tons, for a value of US$2.9 billion. 3
Field observations such as temporal and spatial distributions of mortality, and identification of the affected species, can provide valuable information for the approximate determination of the cause of a fish kill. 14 In this instance, acute intoxication was considered to be unlikely because a single species was affected over a wide geographic area, and because mortality was prolonged, with an apparent peak 2 weeks after the onset. Accordingly, it was suspected that adverse environmental conditions (i.e., extremely high temperatures, oxygen depletion, and low water levels), infectious diseases, or a combination of both were involved. In addition, epidemiologic characteristics of this massive mortality suggested the possible introduction of a new pathogen in a naïve population.
In the last few years, two viral diseases of carp emerged in North America and caused high mortality. Since 1998, koi carp (Cyprinus carpio koi), an ornamental variety of carp, have been affected by epizootics caused by koi herpesvirus (KHV), a recently recognized infective agent. 9 Spring Viremia of Carp Virus (SVCV) was reported for the first time in North America in 2002, when it caused mortality in cultured and wild carp. 5, 6 SVCV, tentatively classified as a vesiculovirus of the Rhabdoviridae family, is listed as a reportable disease by the Office International des Épizooties. 1, 2
The purpose of the study reported here was to determine the cause of this massive carp mortality by diagnostic laboratory investigation, followed by an attempt to experimentally induce the disease.
Materials and Methods
Diagnostic investigation of the carp mortality in the St. Lawrence River
Pathology
Eighteen carp found dead or moribund on 3 and 4 July 2001 (5 and 6 days after initial observation of mortality) were submitted for necropsy to the Québec Regional Center of the Canadian Cooperative Wildlife Health Center, Département de Pathologie et Microbiologie, Université de Montréal, St-Hyacinthe, QC, Canada. Gross examination was performed, and samples from all organ systems were fixed in buffered 10% formalin. Tissues were processed in routine manner for histologic examination, cut at 5-µm thickness, and stained with hematoxylin, phloxin, and saffron (HPS). Sections of selected tissues were stained with Brown and Brenn, periodic acid–Schiff (PAS), Gomori's methanamine silver, and Warthin-Starry techniques. 12 Samples of gills and skin were placed in Karnovsky fixative and processed in routine manner for transmission electron microscopy (TEM). 18
Bacteriology and mycology
Fresh samples of gills, spleen, kidney, and liver were inoculated on blood and MacConkey agars, and selected tissues were inoculated on Sabouraud agar. Selected samples of gills previously frozen and stored at −70°C were subsequently thawed and inoculated on cytophaga agar. Aerobic bacterial and fungal isolation and identification were performed following standard procedures for bacterial and fungal pathogens of fish, 18 and using the API identification system (bioMérieux Canada, Montréal, QC, Canada).
Virology
Extracts from pooled gills, kidney, spleen, and heart tissues previously frozen at −70°C were inoculated on fat head minnow (FMH), epithelioma papillosum cyprini (EPC), and koi fin (KF-1) cell lines. Cell lines were incubated at 15°C for 40 days and observed for cytopathic effects. Three passages of the KF-1 line were performed.
Toxicology
Screening for pesticides was performed using a method for multiresidue analysis of 68 pesticides and metabolites. Purified tissue extracts from eight carp, including muscle, eggs, and liver, were analyzed by gas chromatography using electron capture (ECD) and nitrogen-phosphorus (NPD) detectors. For detection of heavy metals, tissues from three carp were digested by microwave heating in a solution of concentrated nitric acid and hydrogen peroxide, and analyzed by inductively coupled plasma mass spectrometry.
Experimental induction of disease
Animals
In June 2002, 20 wild-spawning, adult common carp, 15 males and five females, were angled from Last Mountain Lake, SK, Canada. Body weight ranged from 1.70 to 5.83 kg (average: 2.87 kg), and length ranged from 49 to 72 cm (average: 57 cm). From June to August 2002, fish were kept in three outdoor circular plastic tanks in Saskatoon, SK, Canada. Each tank contained approximately 3,000 liters of noncirculating water aerated with air stones. Every 4 days, approximately 1,500 liters of water was removed from each tank and replaced with fresh water. Fish were fed a commercial pelleted diet for koi carp (20 g/kg of body weight/day).
Preparation of inocula
A pool of 10 g of brain, skin, and gill tissues collected from carp affected by natural disease in the St. Lawrence River and stored at −70°C were thawed and homogenized in 40 ml of sterile phosphate-buffered saline (PBS). Half of this homogenate was used as a crude (nonfiltered) inoculum. The other half of the homogenate was passed through a 0.22-µm filter (PALL, Gelman Laboratory, Ann Arbor, MI), then was used for inoculation.
Experimental design
Fish were selected by opportune random sampling and assigned to experimental groups in a serial manner: group A (n = 7, fish ID: A1–7), group B (n = 7, fish ID: B1–7) and group C (n = 6, fish ID: C1–6). After 7 days of acclimatization, fish were individually netted, anesthetized by immersion for 5 minutes in water containing 150 mg of tricaine methanosulfonate (Syndel Laboratories, Vancouver, BC, Canada) per liter, marked by opercular clipping, administered 20 mg of prednisolone acetate (Vétoquinol Canada, Lavaltrie, QC, Canada) per kilogram intramuscularly to induce immunosuppression, and put back in appropriate water tanks for recovery. Twenty-four hours later, fish were anesthetized again and inoculated as stated in Table 1. Because the crude inoculum was presumed to contain high numbers of bacteria, it was suspected that its intravascular injection would rapidly induce septicemia and mortality. Therefore, intravascular injection was not performed with the crude inoculum, and the intraperitoneal route was preferred (group A). Intravascular injection was used for the filtered inoculum (group B), and in control fish (group C). After inoculation and for the remainder of the study, tanks were observed three times per day to detect lethargic or dead fish. Lethargic fish were euthanized by a 10-minute immersion in a bath of water containing 500 mg of tricaine methanosulfonate/liter, followed by decapitation with a knife. All fish that were alive at postinoculation day (PID) 54 were euthanized. Precautions were taken to avoid cross-contamination among tanks; distinct equipment was assigned to each tank, and observation, feeding, and removal of dead and lethargic fish always were performed in the following order: tank C, B, and finally, A.
Experimental inoculation: experimental design and results (presence of encephalitis).

sc = subcutaneous, ip = intraperitoneal, and iv = intravenous (administration).
PBS = phosphate-buffered saline.
Pathology
Necropsy was performed within 1 hour after fish were found dead or were euthanized. Gills, dorsal fin, and skin with subcutaneous tissue and muscle from inoculated areas, were fixed in Bouin's solution, and tissue samples from all organ systems were fixed in buffered 10% formalin. Formalin- and Bouin's-fixed tissues were processed in routine manner for histologic examination, cut at 5-µm thickness, and stained with hematoxylin and eosin. Selected tissues were stained with Brown and Brenn, Warthin-Faulkner, Gomori's methanamine silver, Fites, Giemsa, PAS, with and without diastase pretreatment, and Oil Red O techniques. 12 Histologic examination of all tissues was performed by one pathologist (S. Monette), except for the brain samples, which were examined by two pathologists (S. Monette, E. Simko). Histologic tissue sections were coded and analyzed blindly by pathologists without knowledge of treatment groups. In addition, brain from selected fish was fixed in buffered 2.5% glutaraldehyde, and processed in routine manner for TEM.
Bacteriology and mycology
Fresh samples of gills and skin from inoculated areas, as well as spleen and brain from all fish, and pectoral muscle and kidney from selected fish were inoculated on blood, MacConkey, and cytophaga agars. In addition, gills and skin from selected fish were inoculated on Sabouraud agar. Aerobic and anaerobic isolation and identification were performed as described previously.
Virology
Extracts of brain and pooled kidney, heart, spleen, and gill tissues, previously frozen at −70°C, were inoculated on FMH, EPC, KF-1, and Chinook salmon embryo (CHSE) cell lines. Cell lines were incubated at 15°C for 40 days and observed for cytopathic effects. Three passages were performed using the KF-1 line.
Results
Diagnostic investigation of carp mortality in the St. Lawrence River
Twelve of 18 submitted carp were spawning adults, and the remaining six were juvenile fish. Both sexes were equally represented (nine males and nine females). Body weight ranged from 1.20 to 8.18 kg (average: 4.53 kg), and length ranged from 43 to 80 cm (average: 69 cm).
Pathologic changes were observed in a range of organ systems with variable prevalence (Table 2). All submitted fish (18/18) had multifocal, random, 2- to 3-cm-diameter, circular, erythematous, dermal lesions characterized histologically by epidermal spongiosis, ballooning degeneration, and lytic necrosis associated with dermal congestion and hemorrhage, and moderate inflammatory infiltration of granulocytes and lymphocytes (Fig. 1).
Pathologic observations in common carp found dead or moribund in the St. Lawrence River in July 2001.
Skin; Carp 4, naturally affected carp from St. Lawrence River. Epidermal necrosis characterized by pyknosis and karyorrhexis (∗) with a mild inflammatory infiltrate (arrow). HPS. Bar = 30 µm.
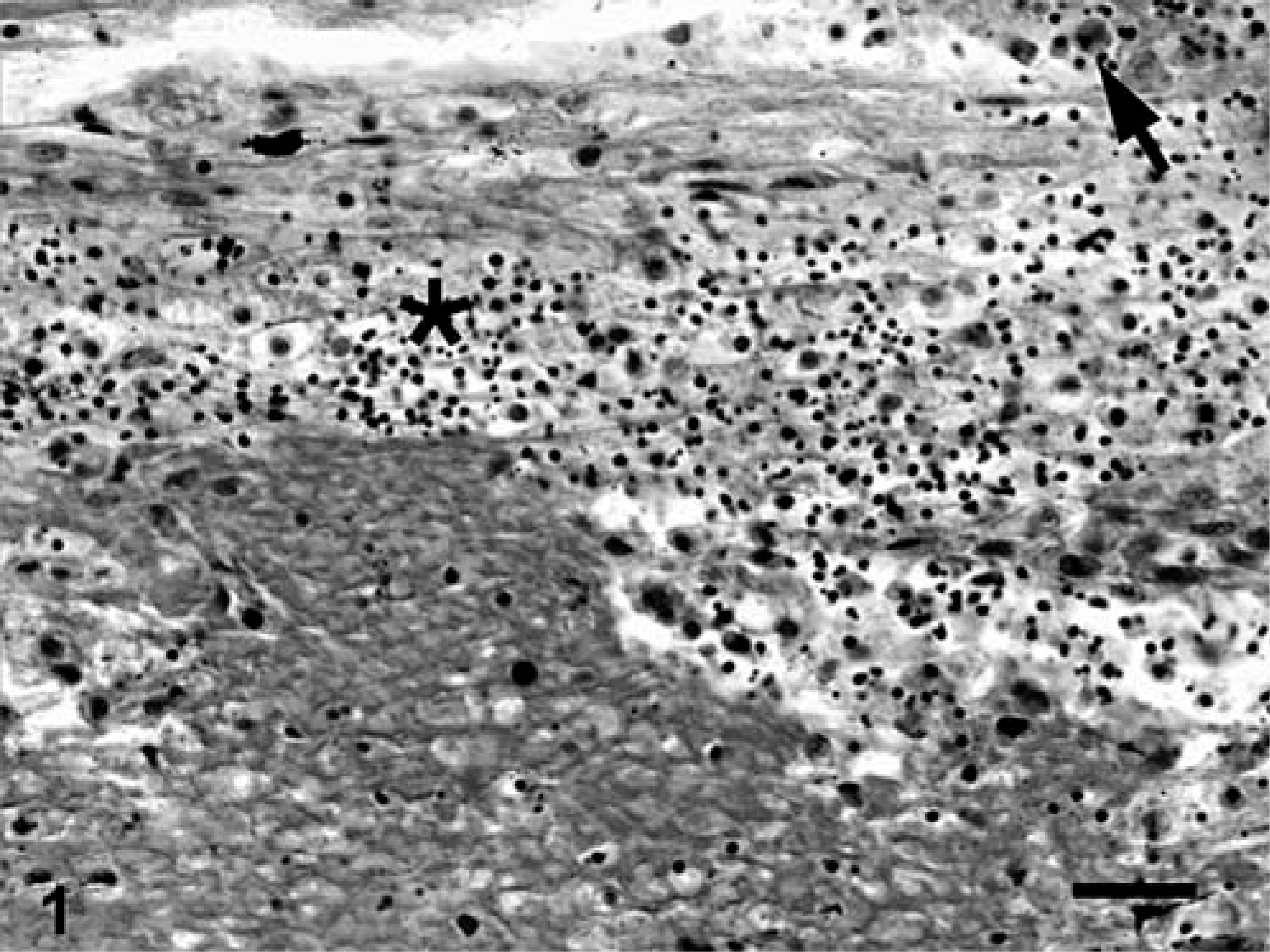
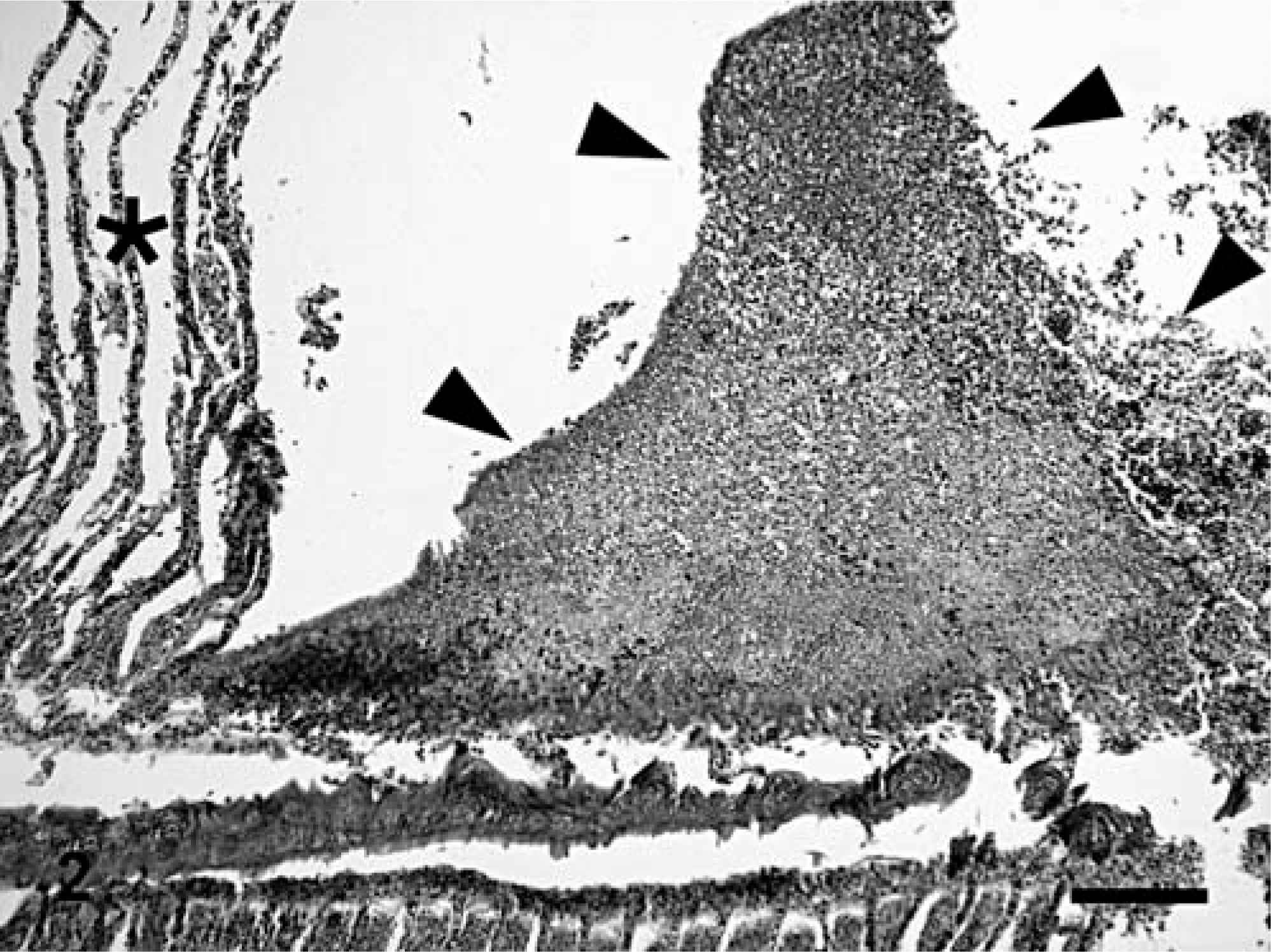
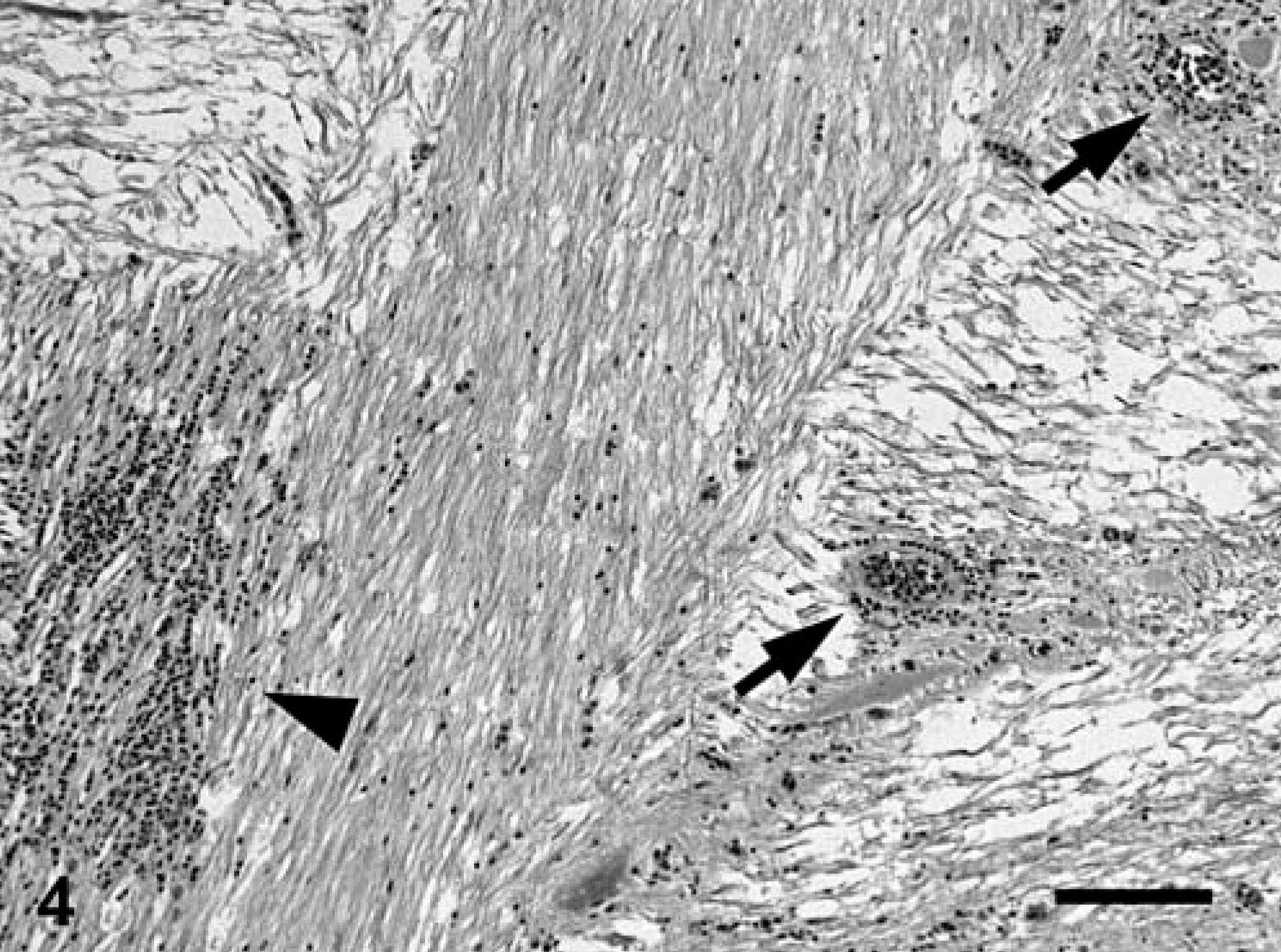
In two-thirds of submitted fish (12/18), branchial lamellae had multiple, pale-tan foci covered by abundant, brown-to-green, friable material. Affected gills were histologically characterized by large foci of granulocytic and lymphocytic necrohemorrhagic branchitis (Fig. 2). Multifocally, there was marked epithelial hyperplasia with fusion of lamellae. Mixed populations of gram-negative bacteria were associated with the foci of necrosis, including a few prominent clusters of gram-negative filamentous bacilli, consistent with Flavobacterium sp. (Fig. 3). In most fish, interlamellar spaces contained small numbers of unidentified species of trematodes, and protozoa that were morphologically consistent with Trichodina spp. These organisms were not associated with inflammation or necrosis.
Gills; Carp 2, naturally affected carp from St. Lawrence River. Necrotizing branchitis characterized by replacement of secondary lamellae by necrotic material and inflammatory exudate (area delimited by arrowheads). ∗Normal lamellae. HPS. Bar = 100 µm.
Gills; Carp 2, naturally affected carp from St. Lawrence River. Necrotic material covering the gills contains numerous filamentous bacilli, approximately 1 × 10 µm, consistent with Flavobacterium sp. Warthin-Starry.
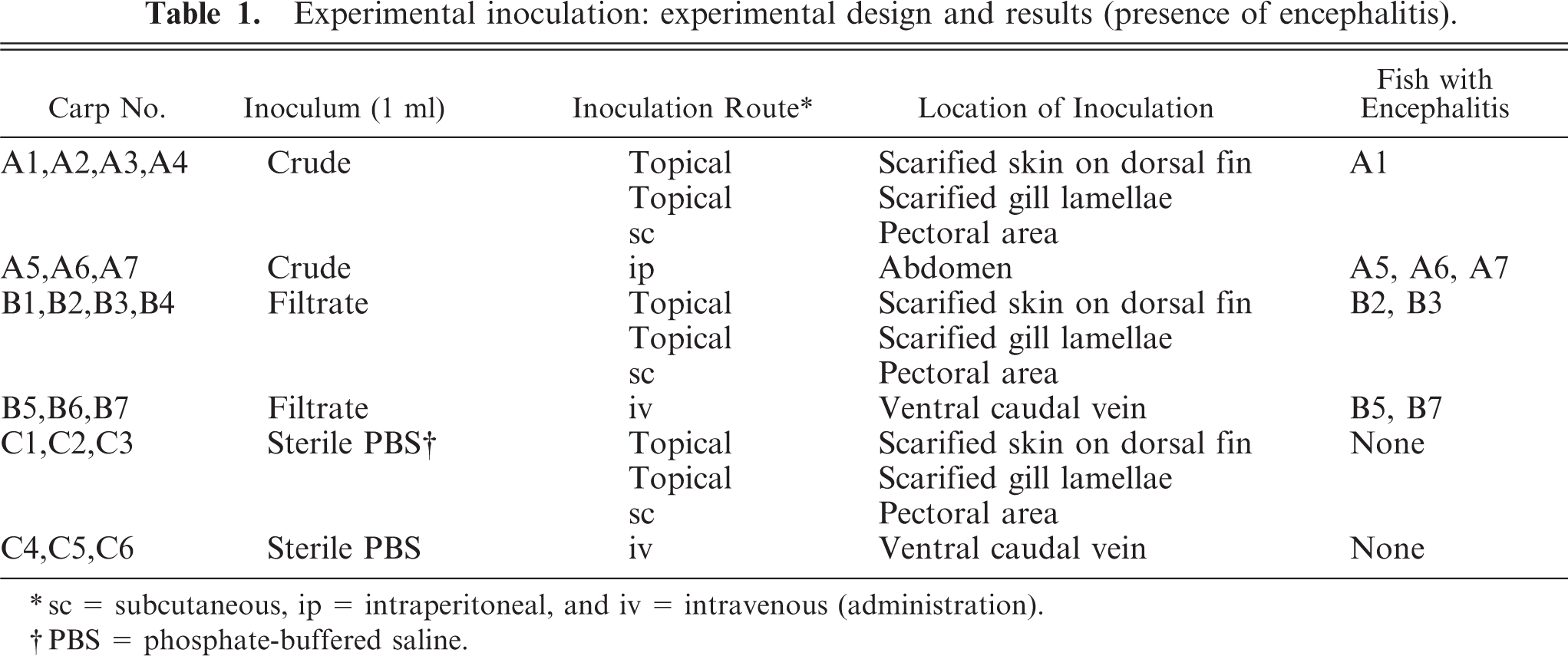
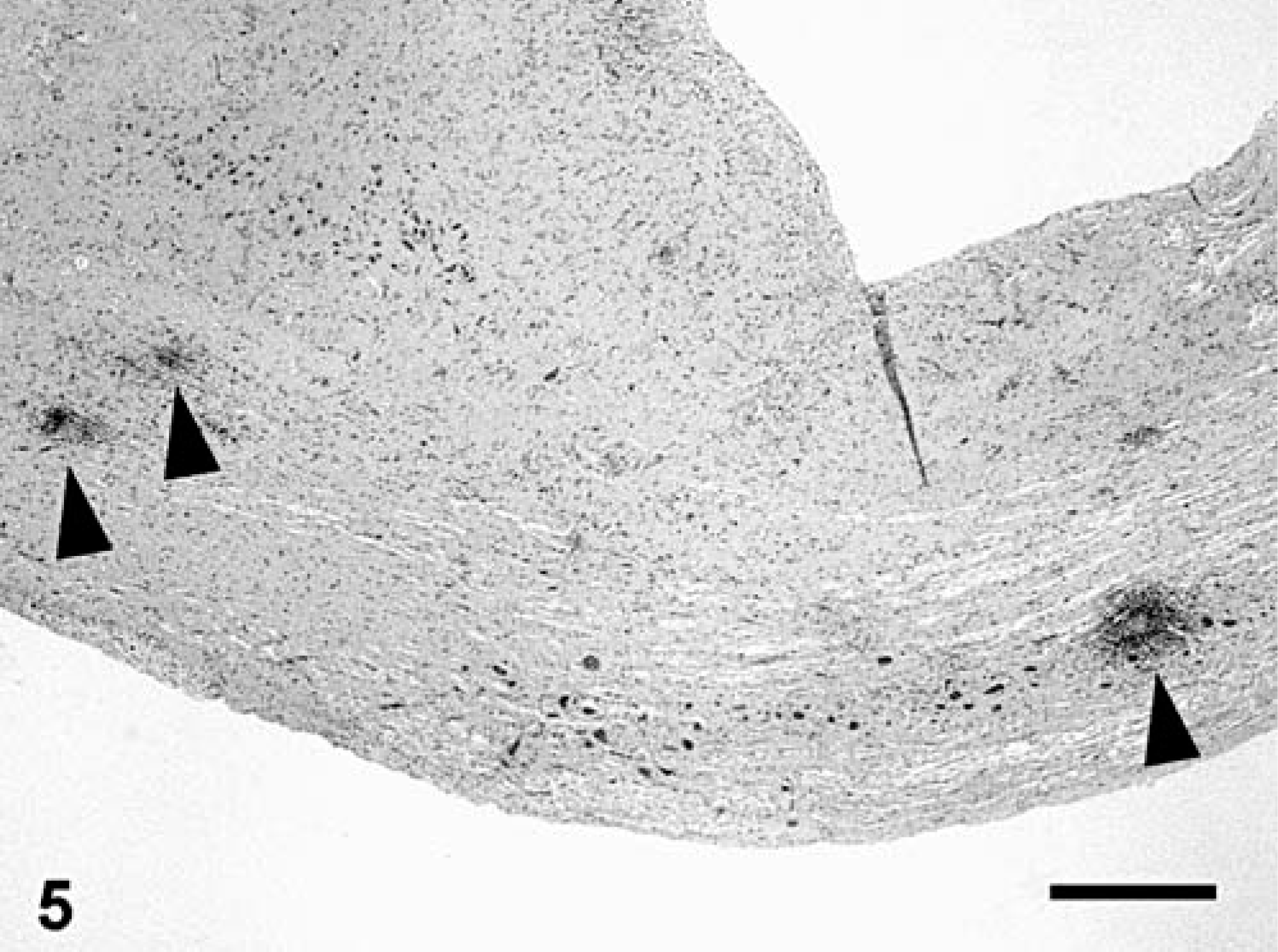
Half of the fish (9 of 18) had evidence of encephalitis characterized by mild-to-moderate multifocal gliosis, perivascular cuffing that was predominantly lymphocytic with fewer histiocytes, and neuronal satellitosis (Fig. 4). The coelomic cavity of 50% (9 of 18) of the fish contained a moderate amount of serosanguineous fluid. A third of the fish (6 of 18) had multifocal intestinal mucosal erosions associated with an inflammatory infiltrate of granulocytes, lymphocytes, and macrophages with a few intrahistiocytic gram-negative bacilli.
Brain; Carp 13, naturally affected carp from St. Lawrence River. Mononuclear perivascular cuffing (arrows) and gliosis (arrowhead). HPS. Bar = 100 µm.
Small to moderate numbers of Aeromonas hydrophila were isolated in pure or mixed culture from the internal organs of most fish. A mixed population of bacteria (A. hydrophila, Acinetobacter sp., Corynebacterium sp., Kluyvera sp., Pseudomonas sp.), and less commonly, fungi (Penicillium sp., Mucorales sp., Scedosporium sp.) was isolated from the gill lesions. Flavobacterium spp. were not isolated.
Inoculated cell lines did not have evidence of cytopathic effects consistent with viral infection. There was no evidence of viral particles on TEM of the gills and skin. Toxicologic analysis of tissues did not reveal abnormal amounts of heavy metals and pesticides.
Experimental induction of disease
Fish B1, A2, and A3 were found moribund and were euthanized on PID 2, 10, and 19, respectively. Fish A4 was found dead on PID 6. All other fish appeared clinically normal throughout the study, and were euthanized on PID 54.
The subcutaneous injection of fish A2, A3, and A4 with crude inoculum resulted in severe, locally extensive, cavitating, hemorrhagic, pyogranulomatous fasciitis and myositis. In addition, fish A4 had several, multifocal, 5- to 10-mm-diameter foci of ulcerative dermatitis and stomatitis, as well as severe multifocal, hemorrhagic, necrosuppurative branchitis affecting approximately 50% of the gills bilaterally. Histopathologic changes of skin and gill lesions were similar to those seen in the natural outbreak, and there were prominent colonies of gram-negative filamentous bacilli associated with those lesions. Fish B1 had widespread petechial hemorrhages of the intestinal mucosa.
Encephalitis was present in four of seven (57%) fish in group A, four of seven (57%) fish in group B, and none of six (0%) fish in group C (Table 1); however, none of the affected fish manifested any clinical signs of neurologic dysfunction during the study. Major histologic features of the encephalitis were moderate-to-marked, multifocal aggregates of mononuclear cells in the neuropil (gliosis) (eight of eight affected fish) accompanied by perivascular cuffing that was predominantly lymphocytic, with fewer histiocytes (three of eight) and neuronal satellitosis (two of eight) (Figs. 5, 6). These changes were most severe and most commonly present in the myelencephalon (eight of eight), followed by the metencephalon (six of eight), diencephalon (five of eight), mesencephalon (one of eight), and telencephalon (one of eight). In the fish that were not affected by encephalitis, including all fish in group C, a few, small, single aggregates of glial cells were present in the neuropil of various regions of the brain. These glial aggregates were distinct in nature and severity from the aforedescribed inflammatory lesions, and were interpreted as normal glia or a mild glial reaction. There was 95% agreement between independent interpretations of the two pathologists who evaluated the brains from experimental fish.
Brain, myelencephalon; carp A6, experimentally inoculated. Multifocal gliosis (arrowheads). HE. Bar = 500 µm.
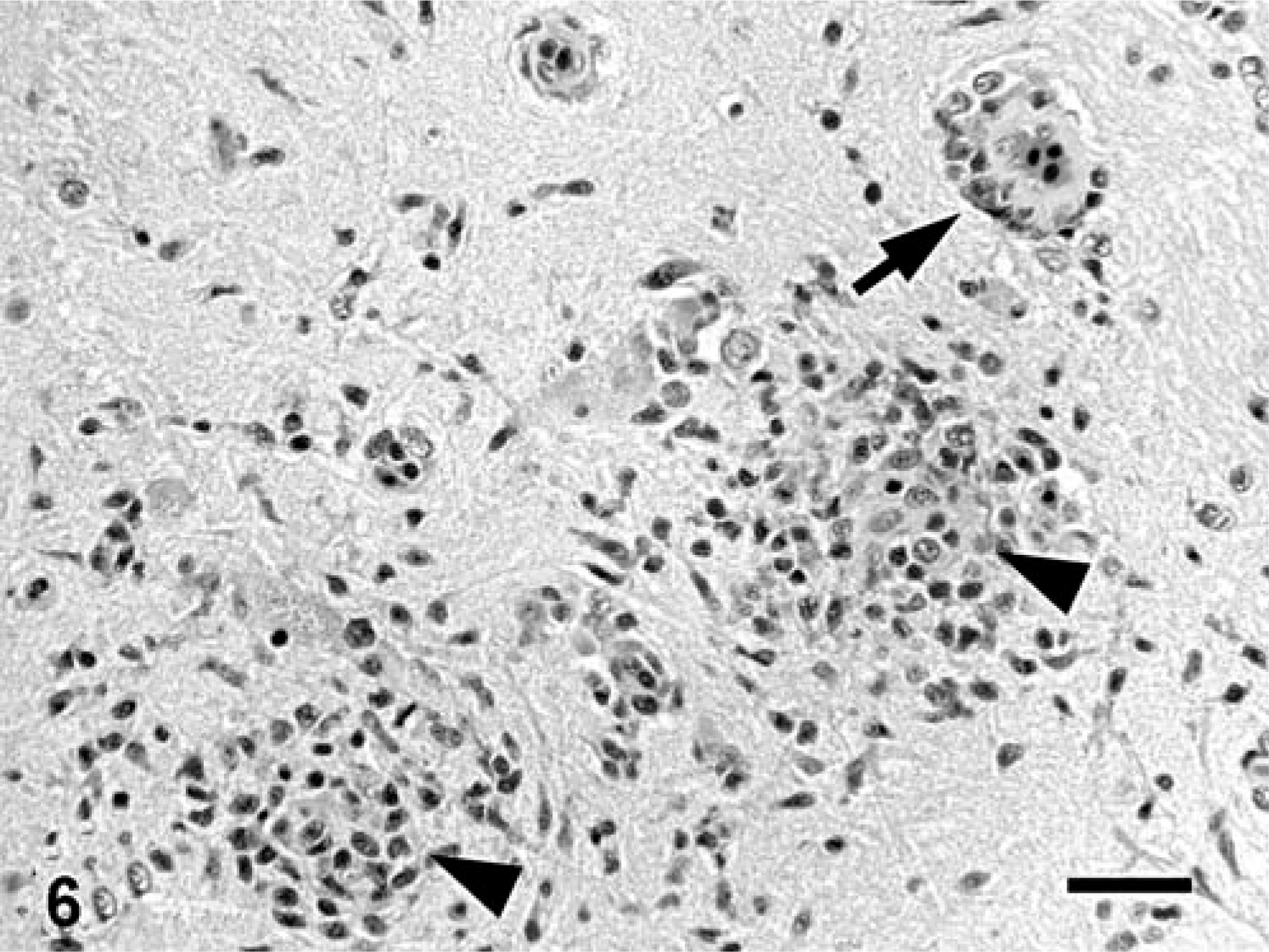
Brain, diencephalon; carp B7, experimentally inoculated. Gliosis (arrowheads) and mononuclear perivascular cuffing (arrow). HE. Bar = 25 µm.
Mixed bacterial agents, including A. hydrophila, Klebsiella sp., Enterococcus sp., and Escherichia coli were isolated from the site of necrotizing myositis of fish A2, A3, and A4 and from the gill lesions of fish A4. In addition, Flavobacterium columnare was isolated from the gill lesions of fish A4, and from normal gills and skin of several fish in all groups. Other bacteriologic findings observed in most fish were consistent with normal flora of the skin and gills.
Virus-induced cytopathic effects were not observed in inoculated cell lines. TEM of the brains of two fish affected by marked encephalitis did not reveal any evidence of viral particles.
Discussion
A definitive single cause of the massive mortality that affected carp in the St. Lawrence River in 2001 was not determined either by diagnostic investigation or by an attempt to experimentally induce the disease. On the basis of our findings from both investigations, we concluded that mortality was caused by opportunistic infections secondary to immunosuppression induced by physiologic (i.e., spawning) and adverse environmental (i.e., extremely high temperatures, oxygen depletion, and low water levels) stressors, and possibly enhanced by an infection causing subclinical encephalitis observed in 50% of fish affected by natural disease and induced in 57% of experimentally inoculated fish. On the basis of epidemiologic features of the natural outbreak, the introduction of a new pathogen in the carp population of the St. Lawrence River also was initially suspected. However, KHV and SVCV, two highly pathogenic viruses of cyprinids that recently emerged in North America, were ruled out on the basis of the absence of typical lesions, 9, 20 and failure to isolate these viruses on the appropriate cells lines. 4, 9
All fish from the natural outbreak submitted for necropsy were affected by opportunistic bacterial infections causing septicemic and gill lesions. Multifocal necrotizing dermatitis, coelomic inflammatory effusion, and erosive enteritis are common findings in bacterial septicemia of fish. 16 Although the interpretation of bacteriologic results in fish from natural outbreaks is complicated by rapid postmortem bacterial overgrowth and contamination of tissues, the isolation of A. hydrophila in pure or mixed culture from internal organs of most fish was considered relevant and consistent with the septicemic lesions observed. A. hydrophila is a saprophytic organism present in most aquatic environments, and is a part of the normal intestinal flora of fish. If the immune system of a fish is compromised, A. hydrophila can invade tissues and cause septicemia. 16
Severe necrotizing branchitis, which was observed in 66% of fish examined from the natural outbreak, is not a typical feature of infection with A. hydrophila. 16 Gill necrosis associated with high mortality is a common syndrome in cultured common carp in Europe, and is believed to be a multifactorial disease caused by a combination of infectious and noninfectious factors. Of the multiple agents that have been isolated from carp affected by this syndrome, F. columnare has been suggested to be the most important. 8 This organism is a common cause of gill necrosis and cutaneous ulceration in many species of freshwater fish. 16 The necrotizing gill lesions associated with prominent, filamentous, gram-negative bacilli that were observed in carp from the St. Lawrence River are consistent with Flavobacterium sp. (most likely F. columnare) infection. Since initial bacterial culture using fresh tissue samples from naturally affected fish was not performed on cytophaga agar (required for growth of Flavobacterium spp. 16 ), a limited number of frozen gill samples were subsequently thawed and inoculated on this medium, which did not result in growth of Flavobacterium spp. Failure to isolate the organism could have been caused by freezing and thawing, or by the selection of nonrepresentative samples. On the basis of histologic findings, Flavobacterium sp. probably contributed to necrotizing branchitis, despite lack of confirmation by culture results. F. columnare can be part of the bacterial flora of healthy fish. Development of disease is influenced by strain pathogenicity, host susceptibility, and environmental conditions. Water temperature is particularly important, as most outbreaks occur at temperatures >18°C. 16 Infections with A. hydrophila and F. columnare are often the ultimate cause of death in fish kills. As these are opportunistic pathogens, determination of the predisposing factors causing immunosuppression is crucial when investigating fish mortality.
The most common factor contributing to immunosuppression in fish is stress caused by adverse environmental and physiologic conditions. 10 The environmental conditions in southern Québec in the Spring and Summer of 2001 were atypical and have been reported. 15 From 1 May to 30 June, the air temperature in Québec City was highly variable and oscillated multiple times between extreme values that were seldom observed during the past 30 years. Unusual water temperature of 34°C was recorded in shallow waters of Saint-Pierre Lake in June, which is also constraining for fish activity (mainly for large adults) because of oxygen depletion. Water levels of the St. Lawrence River in Sorel were extremely low and close to historic records from the preceding 40 years. In early summer, adult carp gather in large numbers in grassy or weedy shallow waters for spawning. 19 Those areas are sheltered from water exchange, and are exposed to extreme variations in temperature, which might have been exacerbated by low water levels. However, those shallow waters were also populated by other species of fish, such as the largemouth bass (Micropterus salmoides), brown bullhead (Ameiurus nebulosus), and pumpkinseed sunfish (Lepomis gibbosis), which were not affected by mortality during this period, 15 suggesting that adverse environmental conditions were not the only predisposing factors for mortality. Since carp are robust fish that are relatively resistant to thermal stress and oxygen depletion caused by high water temperature, their reproductive behavior might have contributed to increased susceptibility to those adverse environmental conditions, resulting in immunosuppression and opportunistic infections.
Opportunistic infections in fish can also result from preexistent diseases that compromise the host defense system. The lymphocytic encephalitis observed in 50% of fish from the natural outbreak might have been a manifestation of an underlying viral infection, which could have contributed to immunosuppression. Systemic inflammatory conditions such as bacterial septicemia can result in cerebral gliosis in fish 17, 21 ; however, the changes observed in these carp met the criteria of a true encephalitis. 22
Attempt to induce disease and mortality by experimental challenge of captured carp with crude and filtrate inocula prepared from naturally affected fish was partially successful. Three of the four fish inoculated subcutaneously with the crude inoculum developed a severe localized inflammatory reaction, likely caused by the high number of bacteria present in crude inoculum. One of those fish (A4) also developed lesions consistent with F. columnare infection. One of the fish (B1) that received filtered material developed lethargy and was euthanized shortly after inoculation, but necropsy did not reveal the cause of lethargy. The most relevant finding in experimentally inoculated fish was the induction of encephalitis in 57% of fish injected with crude or filtrate inoculum. However, we did not observe a higher prevalence of opportunistic infections in these fish under the experimental conditions. All fish with encephalitis survived until the end of the experiment (PID 54) without any observable clinical signs of neurologic dysfunction, and 75% of them did not have any other organ system affected by a pathologic process. The brain lesions in experimental fish were similar to those observed in affected carp from the St. Lawrence River, which suggests that an infective agent causing lymphocytic encephalitis was experimentally transmitted. Since both groups that received either crude or filtrate inoculum were affected with encephalitis, the putative etiologic agent would be <0.22 µm in diameter, which is consistent with a virus. However, we could neither isolate any virus in four cell lines nor visualize any viral particles in the brain by TEM.
Failure of initial isolation or visualization, or both, of viral agents is not uncommon in the study of fish diseases. Infectious salmon anemia was described in 1988, 23 documented to be caused by a virus by experimental transmission using filtered inoculum in 1991, 24 viral particles were visualized in 1994, 11 and the virus was isolated in 1995. 7 Further investigation is planned to characterize the putative agent that caused encephalitis in carp of this study. The fish were administered an immunosuppressive dose of prednisolone acetate prior to inoculation to increase the probability of infection and development of disease. However, the role of immunosuppression in the development of encephalitis could not be determined, as all experimental groups received prednisolone.
In conclusion, the cause of massive carp mortality in the St. Lawrence River in 2001 was considered to be multifactorial. Opportunistic bacterial infections with A. hydrophila and Flavobacterium sp. were the ultimate cause of mortality. High temperatures and low water levels combined with spawning stress were considered to be relevant underlying factors in the development of these infections. The cause of encephalitis observed in the natural outbreak and subsequently induced experimentally was not determined definitively in this study, but a viral cause was suspected.