
Abstract
Locoweed poisoning has been reported in wildlife, but it is unknown whether mule deer (Odocoileius hemionus)are susceptible. In areas that are heavily infested with locoweed, deer and elk (Cervus elaphus nelsoni)have developed a spongiform encephalopathy, chronic wasting disease (CWD). Although these are distinct diseases, no good comparisons are available. The purpose of this study was to induce and describe chronic locoweed poisoning in deer and compare it with the lesions of CWD. Two groups of four mule deer were fed either a complete pelleted ration or a similar ration containing 15% locoweed (Oxytropis sericea). Poisoned deer lost weight and developed a scruffy, dull coat. They developed reluctance to move, and movement produced subtle intention tremors. Poisoned deer had extensive vacuolation of visceral tissues, which was most severe in the exocrine pancreas. Thyroid follicular epithelium, renal tubular epithelium, and macrophages in many tissues were mildly vacuolated. The exposed deer also had mild neuronal swelling and cytoplasmic vacuolation that was most obvious in Purkinje cells. Axonal swelling and dystrophy was found in many white tracts, but it was most severe in the cerebellar peduncles and the gracilis and cuneate fasciculi. These findings indicate that deer are susceptible to locoweed poisoning, but the lesions differ in severity and distribution from those of other species. The histologic changes of locoweed poisoning are distinct from those of CWD in deer; however, the clinical presentation of locoweed poisoning in deer is similar. Histologic and immunohistochemical studies are required for a definitive diagnosis.
Keywords
Locoweeds are plants of the Astragalus and Oxytropis genera that induce a storage disease similar to genetic mannosidosis. Toxicosis is attributed to swainsonine, an endophyte-produced indolizidine alkaloid. 4, 7 Swainsonine inhibits lysosomal α-mannosidase and Golgi mannosidase II, resulting in cellular vacuolation and degeneration. 2 Because prolonged enzyme inhibition is required to develop toxicity, poisoned animals must ingest locoweed for several weeks before they become depressed, anorexic, and lethargic, with muscular weakness, intention tremors, proprioceptive deficits, and obvious behavioral change. Poisoning-induced lethargy and anorexia combined with malabsorption and decreased feed efficiency result in wasting and cachexia. 20, 26 Reported histologic lesions include widespread neuronal vacuolation and axonal dystrophy, as well as vacuolation of the kidney, liver, and various endocrine cells. 20 Some tissues appear to be more susceptible to locoweed-induced lesions because changes are first seen in the proximal convoluted tubules of the kidney, transitional epithelium of the urinary bladder, reticular cells or macrophages of the lymph nodes, liver, spleen, and finally neurons of the cerebellum, cerebrum, basal ganglia, hippocampus, thalamus, midbrain, and autonomic ganglia. This cellular vacuolation is probably better described as microvesiculation because the vacuoles are small and often give affected cells a swollen granular appearance. Initially, vacuolation is confined to the neuropil, but with longer durations of poisoning, both glia and neurons are affected. 28
Locoweed poisoning is relatively common in cattle, horses, sheep, and goats, and the annual economic losses in those species has been estimated at over 20 million dollars. 6 Less is known of the effect of locoweed poisoning on wildlife. Poisoning has been reported in elk (Cervus elaphus nelsoni), with anecdotal reports of poisoning of pronghorn antelope (Antilocapra americana), but it is unknown whether mule deer (Odocoileius hemionus) are susceptible. 1, 31 Although there are minor differences in species susceptibility to locoweed poisoning, swainsonine's remarkable solubility and method of action through enzyme inhibition suggests that all species, including wildlife, will be affected. The relatively recent emergence of chronic wasting disease (CWD) in deer and elk in locoweed endemic areas in Wyoming and Colorado raises questions as to the effects of locoweed poisoning in confounding CWD diagnosis, the spread of CWD, and potential interactions of locoweed poisoning in CWD progression. Although spongiform diseases and locoweed poisoning produce distinct clinical signs and histologic lesions in other species, no comparative studies documenting these differences in deer are available. The purpose of this study was to induce and describe the clinical signs and histologic lesions of chronic locoweed poisoning in deer.
Material and Methods
Oxytropis sericea was collected in 2000 from Union County, New Mexico (Utah State University Herbarium voucher 229290, GPS coordinates 36°43.45′N, 104°1.54′W). Dried locoweed was finely ground and mixed extensively, and random samples were collected for analysis. According to previously described techniques, the swainsonine concentrations were determined to be 0.057%. 3 Two pelleted total rations were formulated. The locoweed ration (ground O. sericea 15%, ground barley 19.25%, ground corn 21.25%, soybean meal 8.75%, wheat bran 16.25%, ground alfalfa 15%, Maxibond 0.2%, molasses 2.5%, Dical 1%, and TM salt 1%) was formulated to obtain a swainsonine dose of 1.5 mg swainsonine/kg body weight (bw)/day. The control ration was similar, with ground alfalfa replacing the locoweed (ground barley 19.25%, ground corn 21.25%, soybean meal 8.75%, wheat bran 16.25%, ground alfalfa 30%, molasses 2.5%, Dical 1%, TM salt 1%). To further verify the swain-sonine dose, the locoweed ration was sampled and analyzed for swainsonine and found to be 0.009%. No swainsonine was detected in the control ration.
Eight hand-raised or trapped, tamed mule deer were randomly divided into two groups of two bucks and two does. The deer were all adults between 2 and 7 years of age. After vaccinating for clostridrial diseases and treating both groups for intestinal parasites, the deer were allowed 3 weeks to become acclimated to the pens and pelleted ration. Both groups were initially fed ad libitum, and the amount eaten was monitored daily. After acclimation, the locoweed group was fed a similar amount of locoweed ration to what the deer ate previously. The amount was monitored, and the control deer were fed equal amounts of control ration the following day (isocaloric controls). The deer were monitored daily for changes in attitude and ability to ambulate. All were tranquilized with xylazine (0.1 mg/kg IM) every 2 weeks to be weighed, and blood and serum were collected for analysis. On study day 180, all the deer were tranquilized with xylazine (0.1 mg/kg IM), euthanized with sodium pentobarbital (0.5 mg/kg IV), and necropsied. At necropsy, tissues were collected and fixed for light and electron microscopic evaluation. Serum and tissues were also collected for swainsonine analysis.
Whole blood was analyzed with an impedance cell counter (Serono Diagnostic 9100, Baker Serono Diagnostics, Allentown, PA). Serum biochemistry was measured with an automated chemistry analyzer (Beckmann Synchron CX-5 chemistry analyzer with Beckman reagents and following the manufacture's recommendations; Beckman Diagnostics, Brea, CA). Serum and tissue swainsonine concentrations were determined with a Jack bean mannosidase inhibition assay that has been previously described. 20, 21 Standard curves for these assays were generated for each tissue, and swain-sonine concentrations were verified by capillary gas chromatography. 3, 8
Tissues were processed for both light and electron microscopy following commonly used techniques. Serial sections of all neurologic sections were also stained with a modified Bodian silver stain and luxol fast blue/periodic acid Schiff stains. 5, 32 For morphometry studies, sagittal sections were cut randomly from the left cerebellar hemisphere and stained with the modified Bodian silver stain. The numbers of missing Purkinje cells seen as empty baskets were counted and enumerated as the numbers of empty baskets per 100 Purkinje cells (1,000 Purkinje cells were counted per animal). All sections of the central nervous system with lesions and sections of thyroid, pancreas, and kidney were stained with biotinylated lectins (Sigma Chemical Co., St. Louis, MO): Arachis hypogea (PNA), Concanavalia ensiformis (Con-A), Glycine max (SBA), Phytolacca americana (PWM), Triticum vulgaris (WGA), and Ulex europaeus (Uea-1) according to previously described techniques. 24 Sections of the obex, including the dorsal motor nucleus of the vagus nerve, tonsil, and mesenteric lymph node, were also stained immunohistochemically for proteinase-resistant protein. 17 For the electron microscopic studies, ultrathin sections of cerebellum were examined with a Zeiss 902 electron microscope.
Data were compared by several techniques. Single observations were compared by analysis of variance in a generalized linear model. Mean body weights and tissue swain-sonine concentrations were also compared by correlation analysis. Mean separations were determined with Duncan's multiple range test at P < 0.05 after a significant F test at α < 0.05 (Proc GLM and COR, SAS Institute, Cary, NC). Repeated hematology and serum biochemistry values were compared over time with the Proc Mixed procedure of SAS. The model included treatments, animals nested within treatments, with repeated measures over time. Animals were a random factor in the model, and various options (e.g., compound symmetry and auto-regressive) were examined to arrive at the best fitting variance–covariance matrix. Significant means (P < 0.05) were separated with Saxton's macro for Proc Mixed. 16 Although all data were evaluated, data from days 1, 60, 120, and 180 are reported. Nonreported data had similar variations from controls.
Results
The locoweed-treated deer initially avoided eating the locoweed ration. However, the ration was mixed with small amounts of rolled barley and molasses until the deer began eating the locoweed ration. The following day, the control deer were fed equal amounts control ration mixed with similar amounts of rolled barley and molasses. Within several weeks, consumption of the locoweed ration increased to amounts similar (∼90%) to the amounts of control ration that were consumed during the pre-exposure period. The control animals consistently ate their entire ration. Because the deer were pen-fed, it is likely that the deer within the group consumed different amounts. However, the estimated dose for the entire group was 1.46 mg swain-sonine/kg bw/day. All exposed animals were observed eating the locoweed pellet, and all seemed similarly poisoned.
Locoweed-poisoned deer were clinically normal for the first 30 exposure days. After 60 days, they began to look rough with dull coats. After 120 days, the poisoned deer were obviously wasting, and they were reluctant to move. No other neuromuscular problems were apparent clinically. However, at 180 days, the locoweed-poisoned deer were thin, with dry, rough hair coats that often did not shed and formed clumps (Fig. 1). Subtle intention tremors were apparent when the animals moved, and all were reluctant to move. Occasionally the poisoned deer would stumble when undisturbed and walking slowly. The control animals were clinically unchanged. All the locoweed deer had lost between 11 and 32 kg with a mean loss of 19 ± 10 kg. One control animal lost 3 kg, but the others gained weight, with mean weight gains of 7.9 ± 8.4 kg.

Deer No. 6947, a buck mule deer (Odocoileius hemionus) treated with a 15% locoweed ration for 180 days, resulting in a daily intake of 1.46 mg swainsonine/kg. Note the emaciation and roughened, irregular coat.
The locoweed-poisoned deer had significantly lower erythrocyte counts, blood hemoglobin, and packed cell volumes on study days 0, 60, 120, and 180 (Table 1). These tended to be lower on day 60, but on days 120 and 180, all were significantly lower (P < 0.05) than controls and pretreated values. However, locoweed treatment did not appear to affect the other erythrocyte parameters, including mean corpuscular volume, corpuscular hemoglobin, and corpuscular hemoglobin content. Although the hematology analyzer did not detect changes in white cell numbers or size, distinct lymphocyte vacuolation was evident after 120 days of locoweed treatment. The vacuoles were nonstaining and generally <1 μm in diameter (Fig. 2).
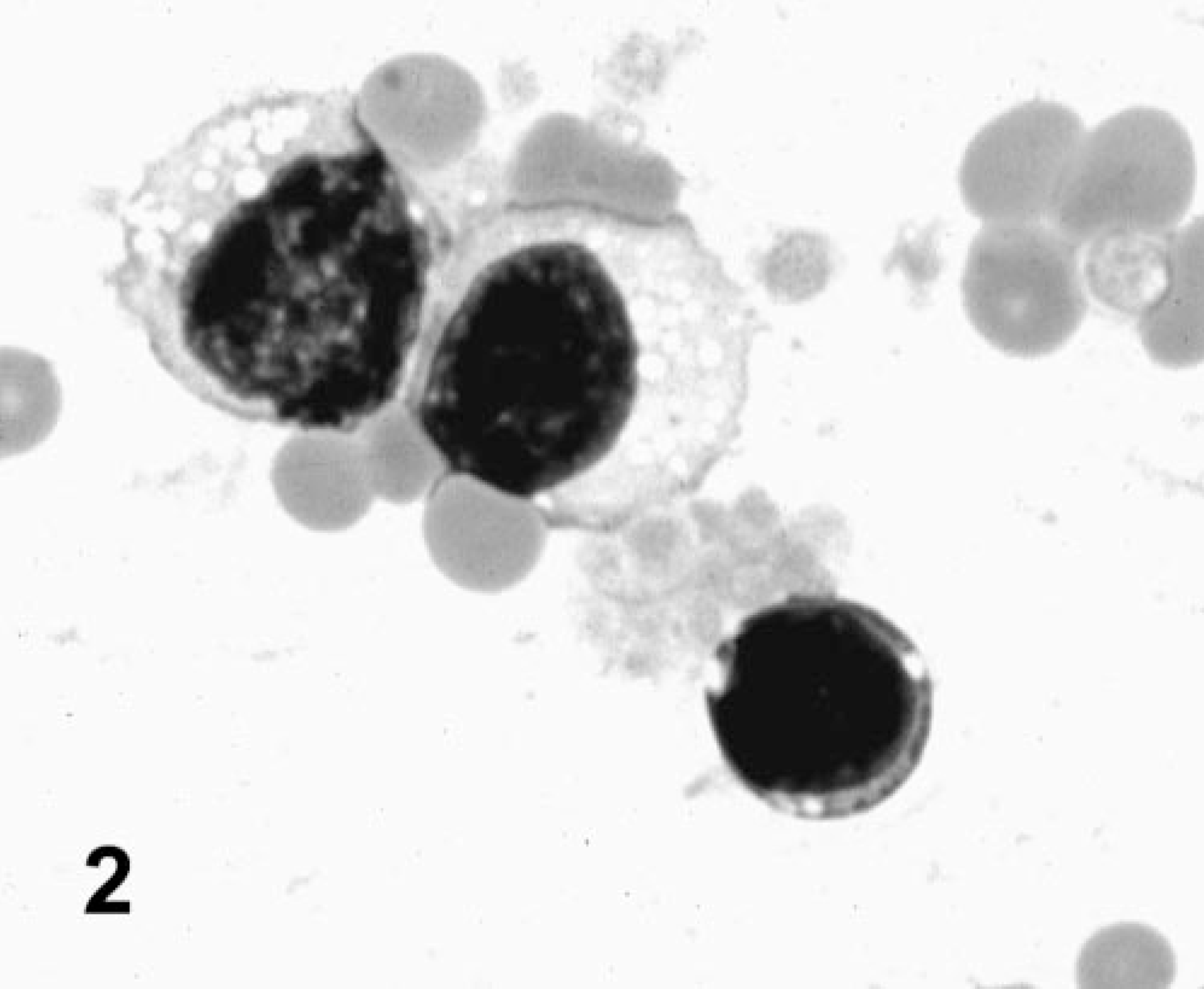
Peripheral blood smear; deer No. 6947. Poisoned with locoweed for 180 days. The lymphocytes contain typical locoweed-induced cytoplasmic vacuoles. Wright Giemsa stain.
Mean hematology results of control deer and deer fed locoweed ration on days 1, 60, 120, and 180.∗
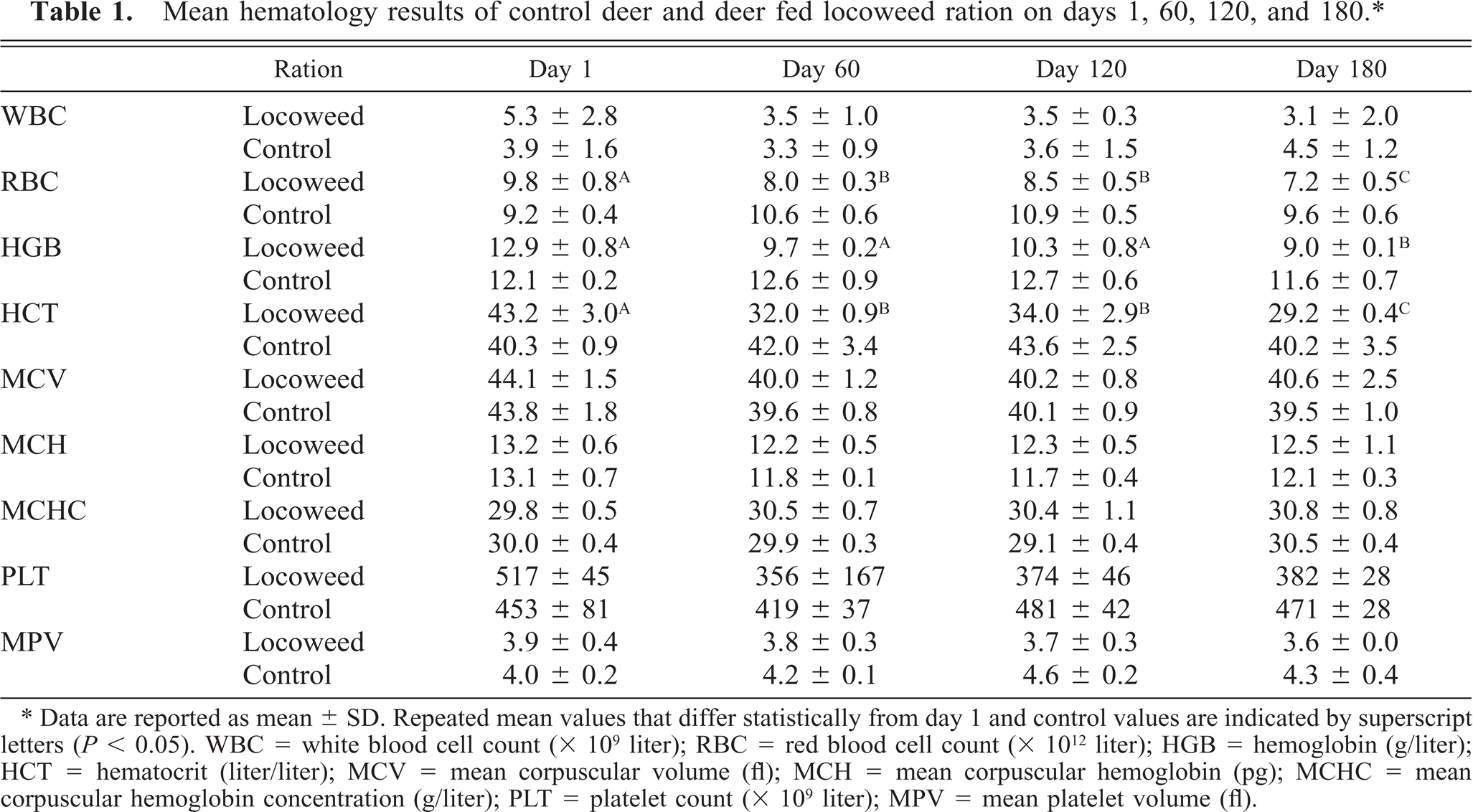
∗ Data are reported as mean ± SD. Repeated mean values that differ statistically from day 1 and control values are indicated by superscript letters (P < 0.05). WBC = white blood cell count (× 109 liter); RBC = red blood cell count (× 1012 liter); HGB = hemoglobin (g/liter); HCT = hematocrit (liter/liter); MCV = mean corpuscular volume (fl); MCH = mean corpuscular hemoglobin (pg); MCHC = mean corpuscular hemoglobin concentration (g/liter); PLT = platelet count (× 109 liter); MPV = mean platelet volume (fl).
Serum biochemical data are reported in Table 2. Blood glucose in the locoweed-poisoned deer was generally within the reference range (pretrial and control deer means ± 2 SD). However after day 120, the poisoned deer became more excitable, and several animals had blood glucose concentrations of 265 and 315 mg/dl. Rarely, the control deer became excited at sampling, and one animal had blood glucose of near 300 mg/dl on day 180. The blood urea nitrogen (BUN) of the locoweed group increased and was significantly elevated on days 60 and 120. The BUN remained slightly elevated on day 180, but the difference was not statistically different from controls (P = 0.11). Creatinine and phosphorus were not elevated in either group at any time.
Selected mean serum biochemical parameters and swainsonine data from control deer and deer fed locoweed pellets.∗
∗ Data are reported as mean ± SD. Data not included that did not differ from those of the controls or from reference values include calcium, albumin, bilirubin (total, direct, and indirect), magnesium, potassium, sodium, chloride, carbon dioxide, osmolality, anion gap, and BUN : creatinine ratio.
† Several animals had values that were out of the reference interval (data from normal deer, mean ± 2 SD).
‡ Locoweed-treated group means that are significantly different (P < 0.05) from initial values (day 1) on that study day.
On days 60, 120, and 180, the locoweed-treated deer had significantly higher alkaline phosphatase (ALP) and aspirate aminotransferase (AST) activities than the control deer. Little change was seen in alanine aminotransferase or lactic dehydrogenase. Gamma gluta-myltransferase (GGT) tended to be lower in the locoweed group, and it was significantly lower than the control group on day 120. Creatine kinase was sporadically higher in the locoweed group but varied widely by animal, so the differences were not statistically different from the control group activities.
The serum mannosidase inhibition assay for swain-sonine indicated that mean serum swainsonine concentration in the locoweed-treated group was about 125 ng/ml throughout the study. This was significantly different from controls and pretreatment concentrations on all of the study days. No significant alterations of mannosidase inhibition activity (swainsonine concentration) was detected in the control animals at any time.
Histologically, all of the locoweed-poisoned deer developed severe neurovisceral cytoplasmic microvacuolation. This vacuolar change was most prominent in the exocrine glands, with extensive vacuolation of the exocrine pancreas (Fig. 3). The endocrine pancreas was spared and appeared to be histologically normal. The mucous and serous salivary glands were similarly vacuolated, with fine vacuolation of the parietal gastric glands. The thyroid follicular epithelial cells were minimally vacuolated (Fig. 4), and the epithelium of the renal convoluted tubules also had minimal cytoplasmic vacuolation. The macrophages in the spleen, lymph nodes, and liver were also mildly vacuolated. Neurons throughout the central nervous system, as well as in most ganglia, were also vacuolated. This neuronal vacuolation was especially prominent in cerebellar Purkinje cells (Fig. 5). Axonal swelling and dystrophy were found in many white tracts but was most severe in the cerebellar peduncles and the gracilis and cuneate nuclei (Figs. 6, 7). This neuronal vacuolation was perikaryonic and microvesicular in nature, giving many neurons a ground glass cytoplasmic appearance. The vacuolation expanded the cytoplasm, with little tendency to displace or compress the nucleus. Bodian silver studies demonstrated that poisoned animals had fewer perineuronal processes and that there was neuronal death, seen as significantly increased numbers of empty baskets (Fig. 8). Locoweed-poisoned deer had 5.6 ± 2.3 empty basket cells/100 Purkinje cells. Control deer had 1.2 ± 1.4 empty baskets/100 Purkinje cells (Figs. 9, 10). Lectin histochemistry showed similar affinity, as seen in previous studies; the vacuoles stained prominently with WGA, PWM, and Con-A (Fig. 11). None of the control or locoweed-treated deer developed lesions suggestive of a spongiform encephalopathy, and there was no evidence of CWD by immunohistochemistry. Electron micrographs (Fig. 12) demonstrated that the Purkinje cell cytoplasmic vacuoles were membrane-bound clusters of amorphous debris.
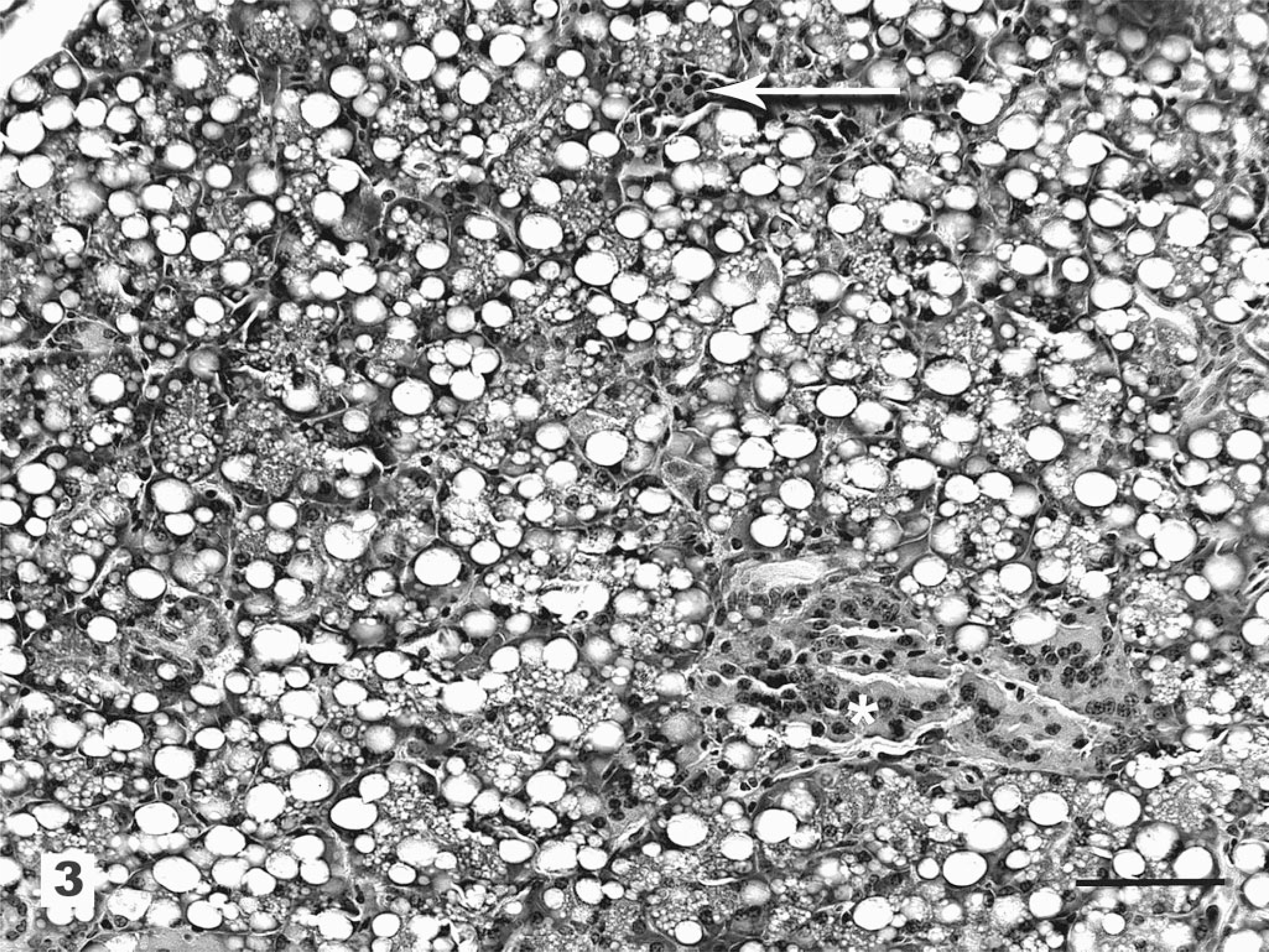
Pancreas; deer No. 6945. Fed a locoweed ration for 180 days. The exocrine pancreas has extensive swelling and vacuolation (arrow) with relative sparing of the islets (∗). HE. Bar = 100 μm.
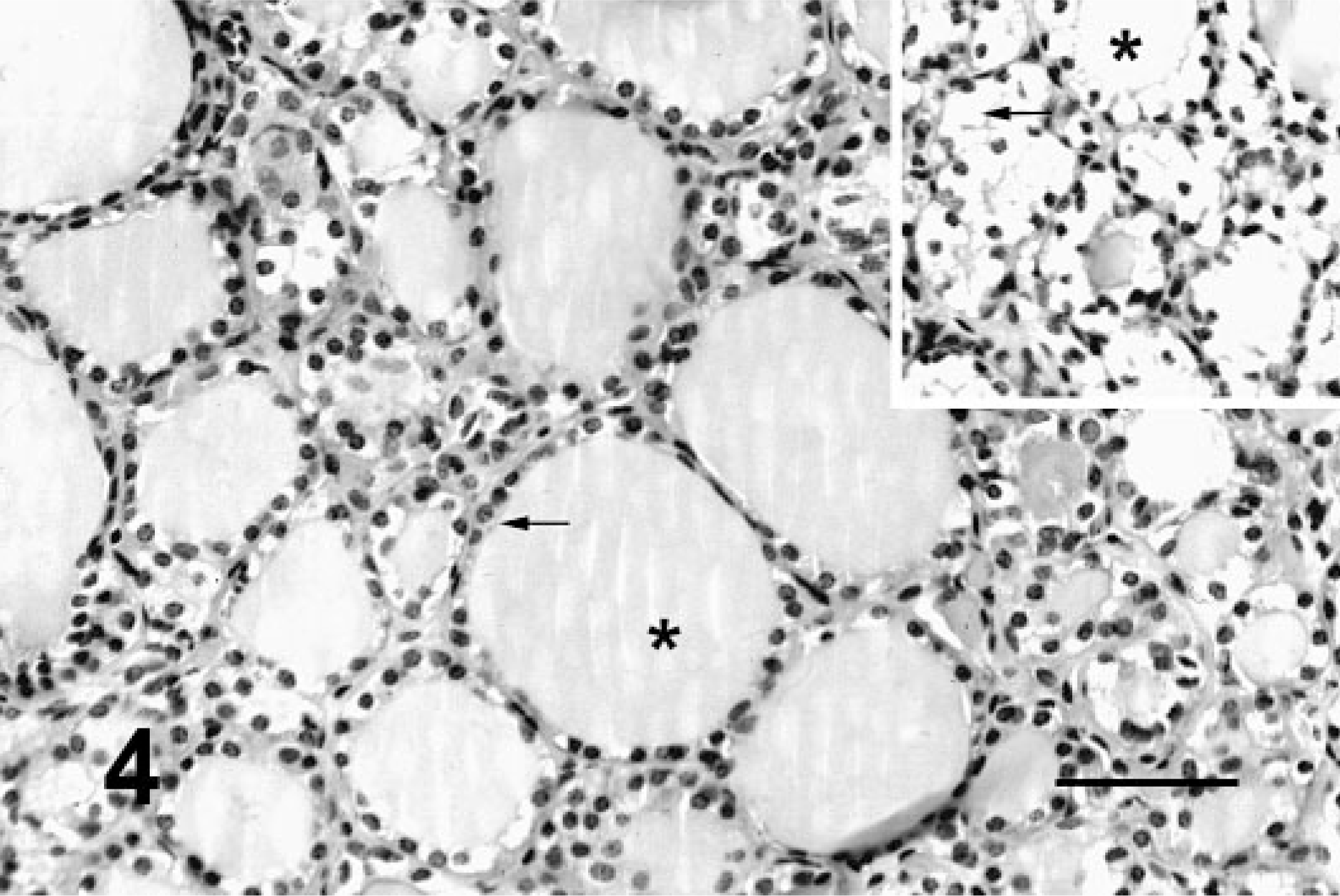
Thyroid; deer No. 6945. Fed locoweed ration for 180 days. The follicular epithelium is minimally vacuolated (arrow) with no change in the colloid (∗). HE. Bar = 100 μm. Inset. Thyroid; locoweed-poisoned horse for comparison. The follicular epithelium has extensive vacuolation. HE. Bar = 50 μm.
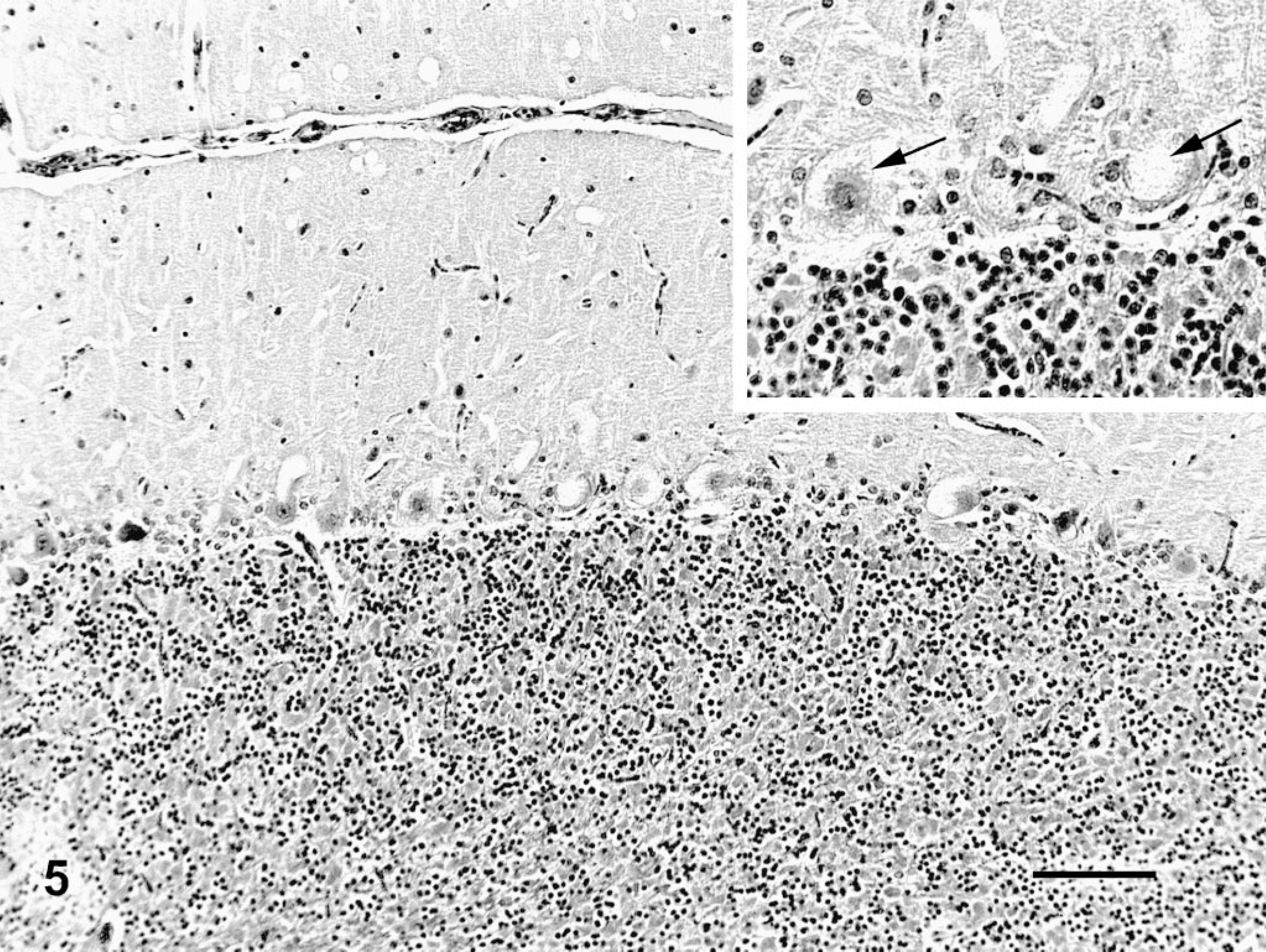
Cerebellum; deer No. 6947. Fed locoweed ration for 180 days. The Purkinje cells are swollen and vacuolated. Inset. Similar vacuolated Purkinje cells at higher magnification (arrows). HE. Bar = 100 μm.
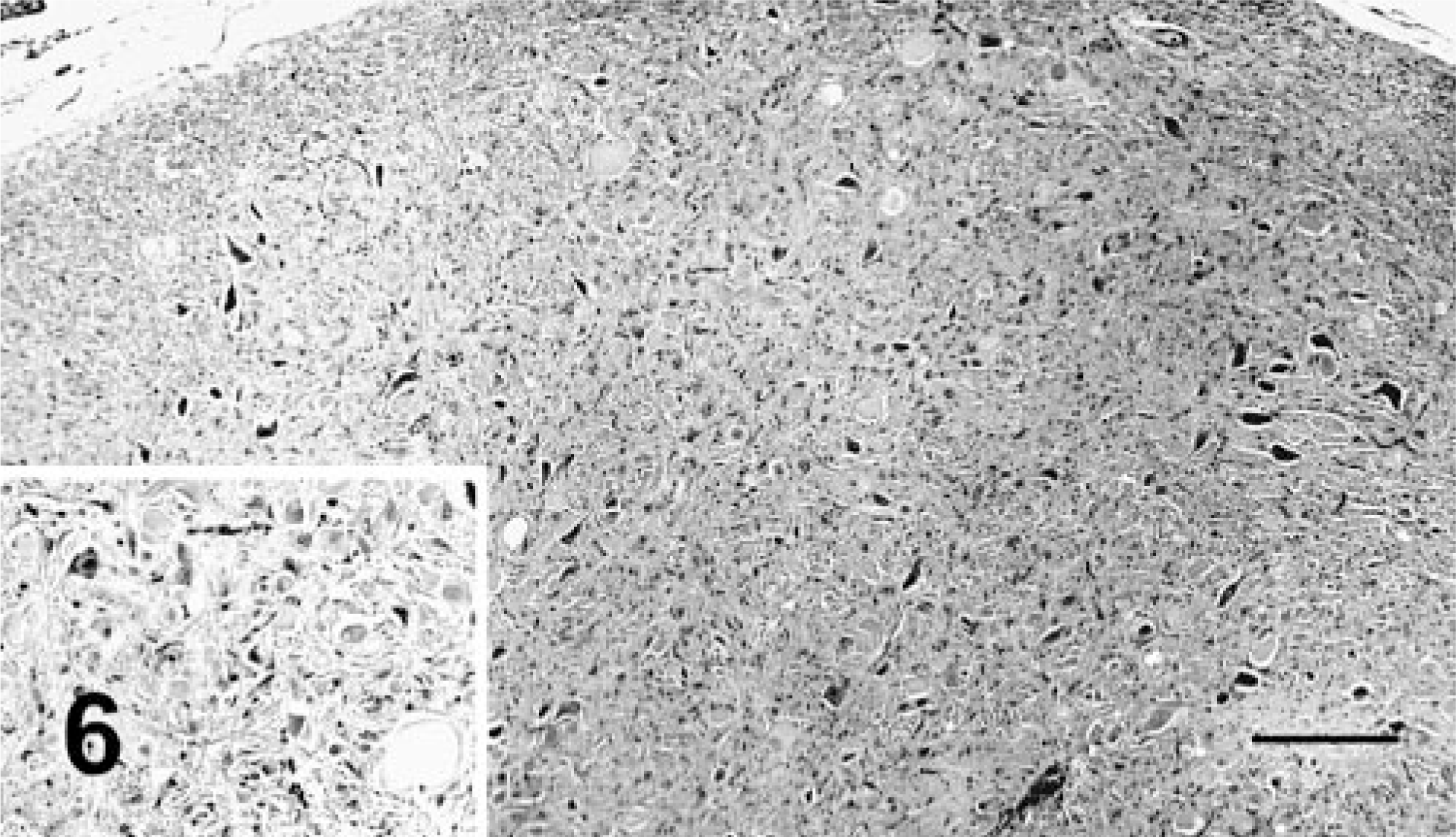
Caudal medulla oblongata; deer No. 6947. Fed locoweed ration for 180 days. The neurons and axons of the fasciculus cuneatus are swollen and vacuolated with numerous spheroids. Inset. A typical spheroid at a higher magnification. HE. Bar = 100 μm.
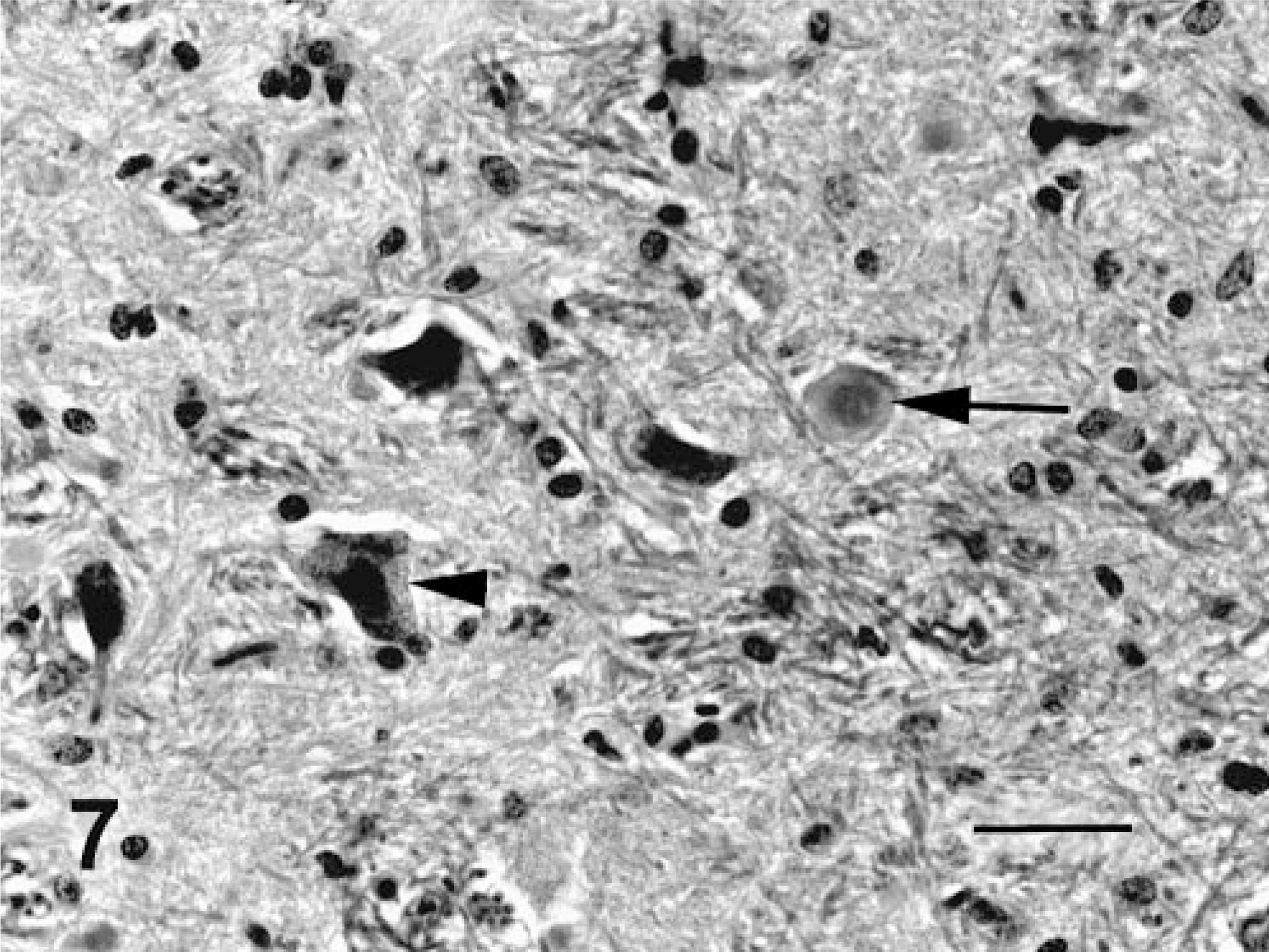
Nucleus gracilis; deer No. 6945. Fed locoweed ration for 180 days. The neurons of the gracilis are swollen and degenerative (arrowheads) with numerous large spheroids (arrows). HE. Bar = 100 μm.
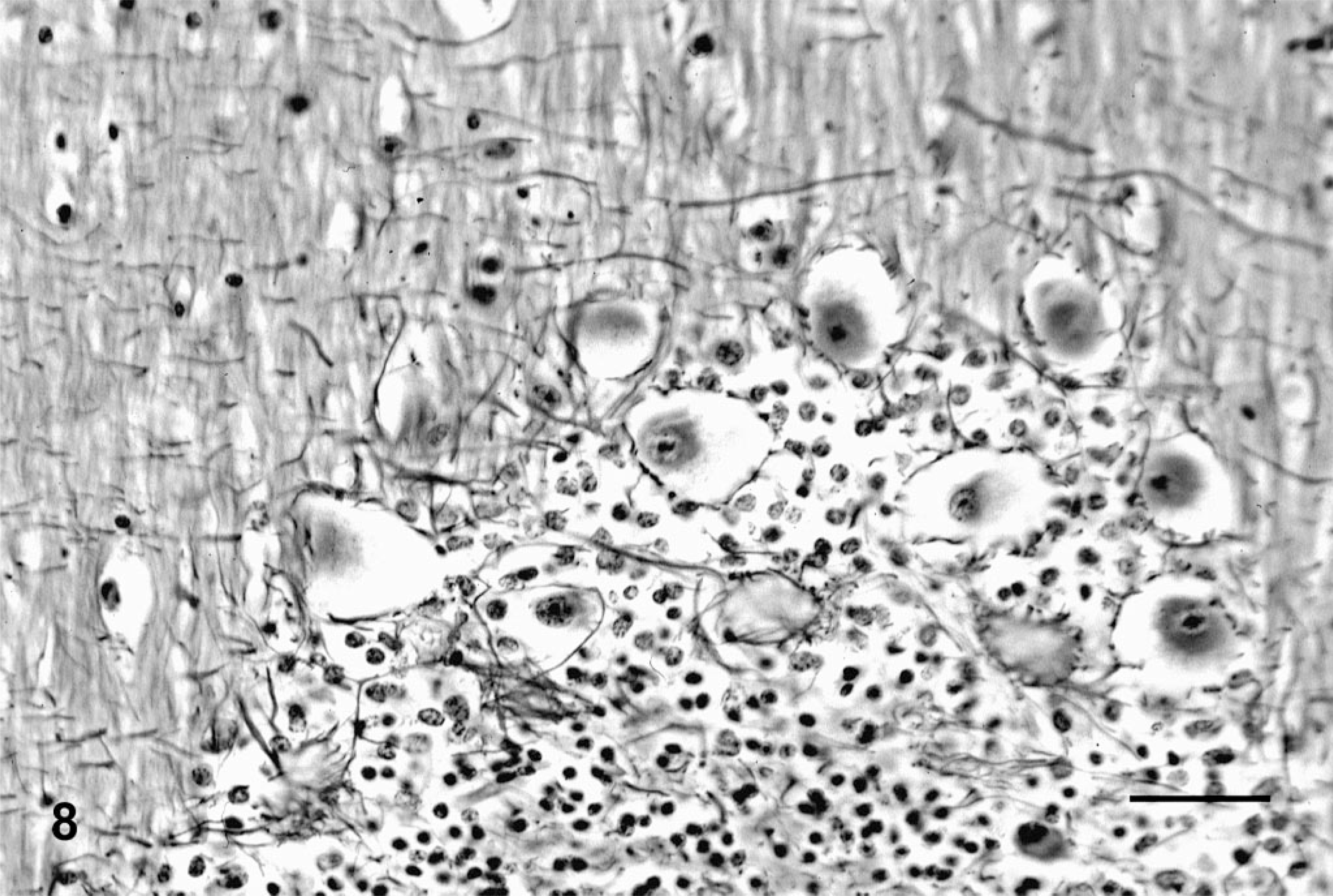
Cerebellum; deer No. 6945. Poisoned with locoweed ration for 180 days. The Purkinje cells are swollen and vacuolated with a paucity of dendritic/axon interactions. Bodian silver stain. Bar = 100 μm.
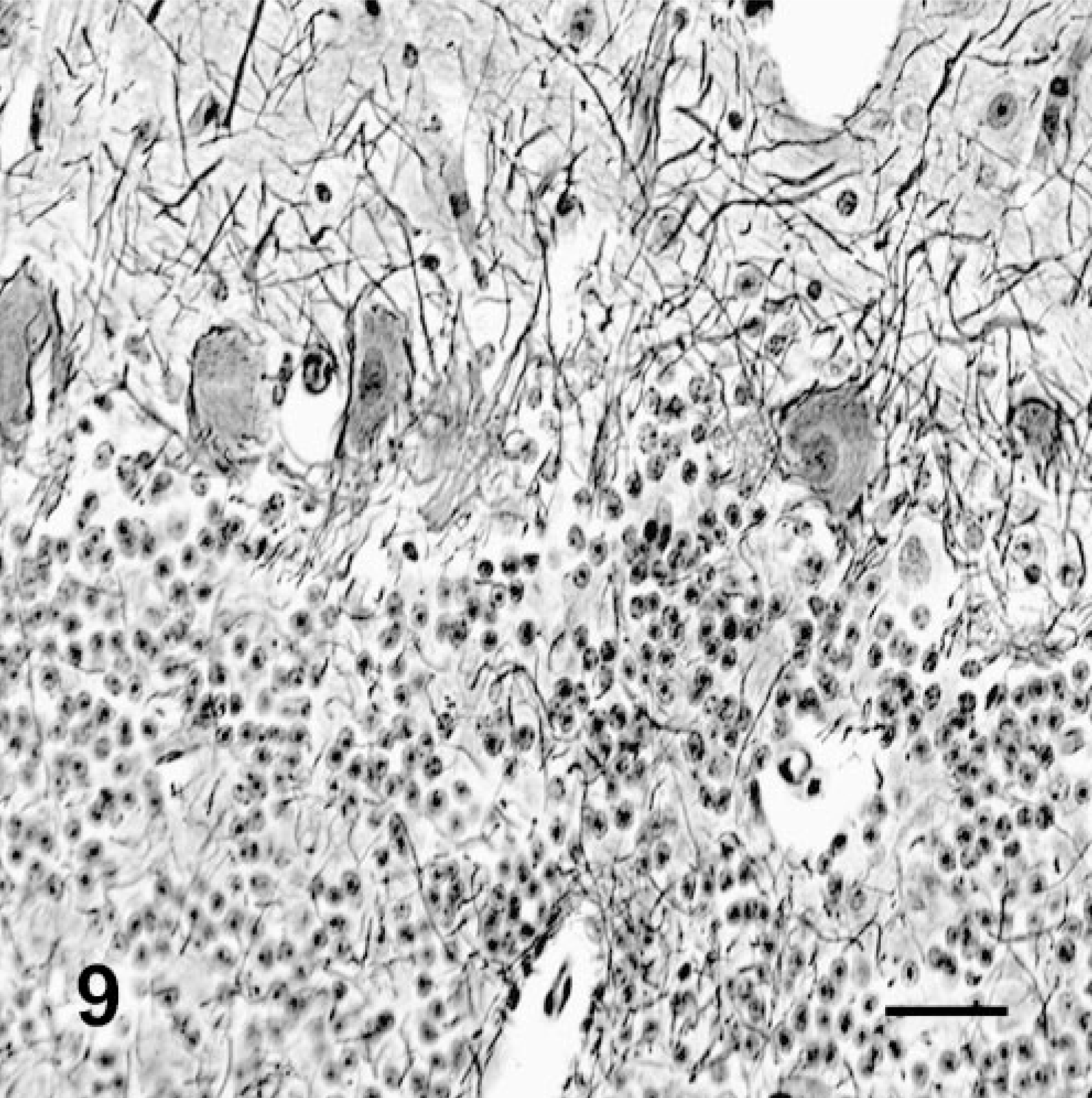
Cerebellum; deer No. 6946. Fed control ration for 180 days. The Purkinje cells have abundant dendritic/axon interactions and lack cytoplasmic vacuolation. Bodian silver stain. Bar = 30 μm.
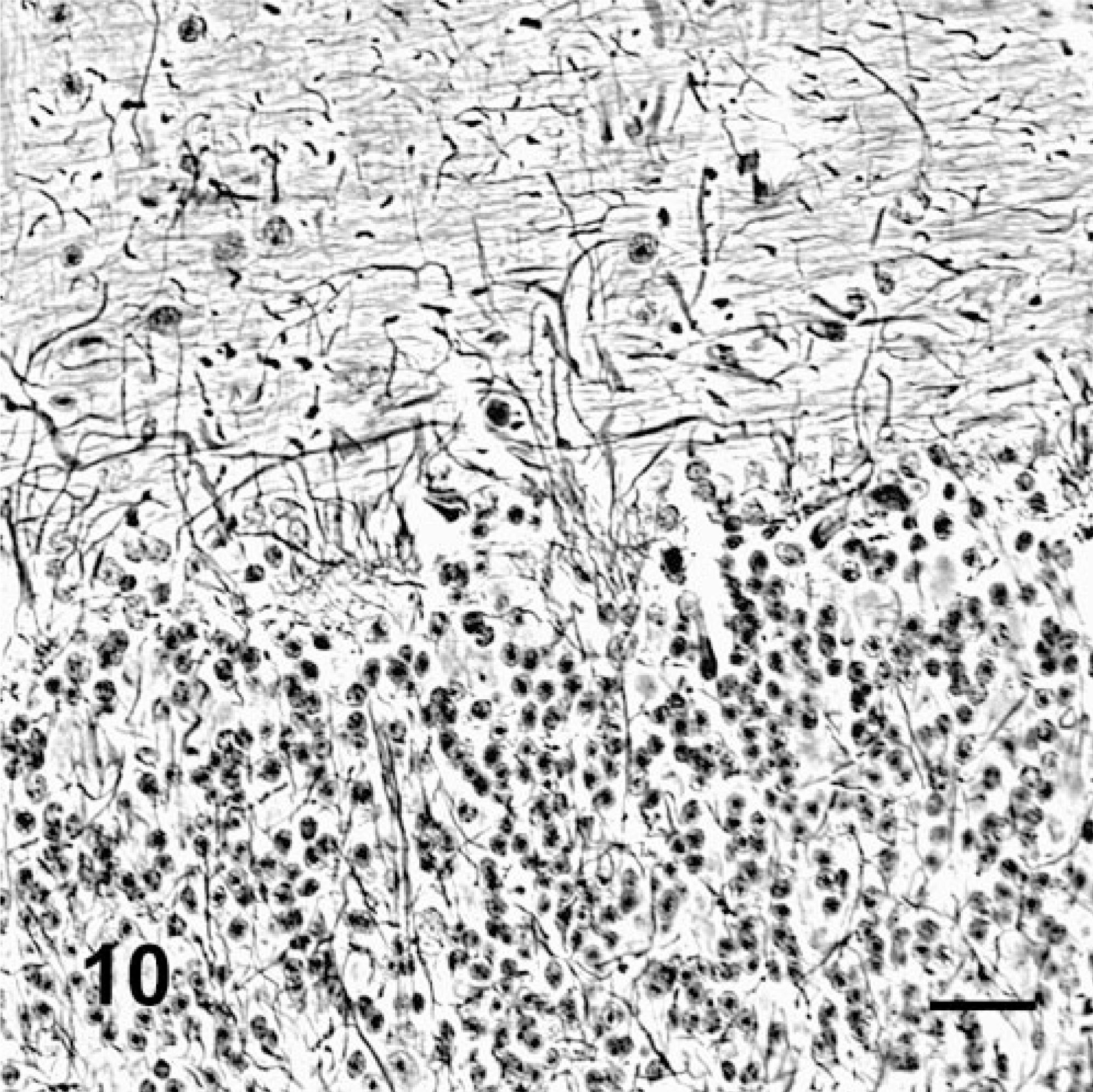
Cerebellum; deer No. 6945. Fed locoweed ration for 180 days. Many of the Purkinje cells are missing with empty Purkinje cell baskets, indicating their previous locations. Bodian silver stain. Bar = 30 μm.
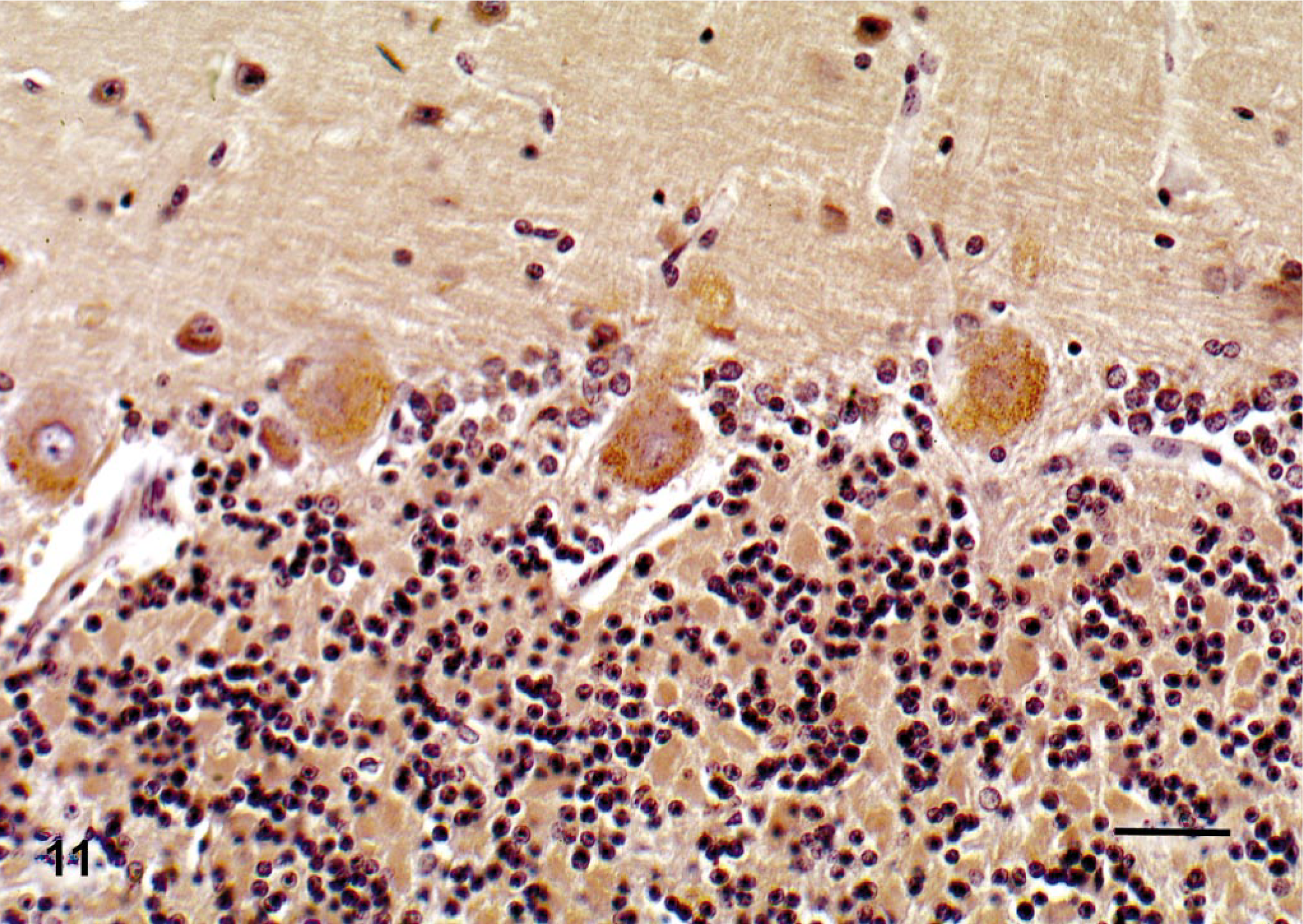
Cerebellum; deer No. 6947. Fed locoweed ration for 180 days. The Purkinje cells are vacuolated with extensive positive staining (mannose rich glucosaccharides) vacuolation indicating increased concentrations of mannose rich saccharides. Con-A lectin histochemistry with hematoxylin counterstain. Bar = 100 μm.
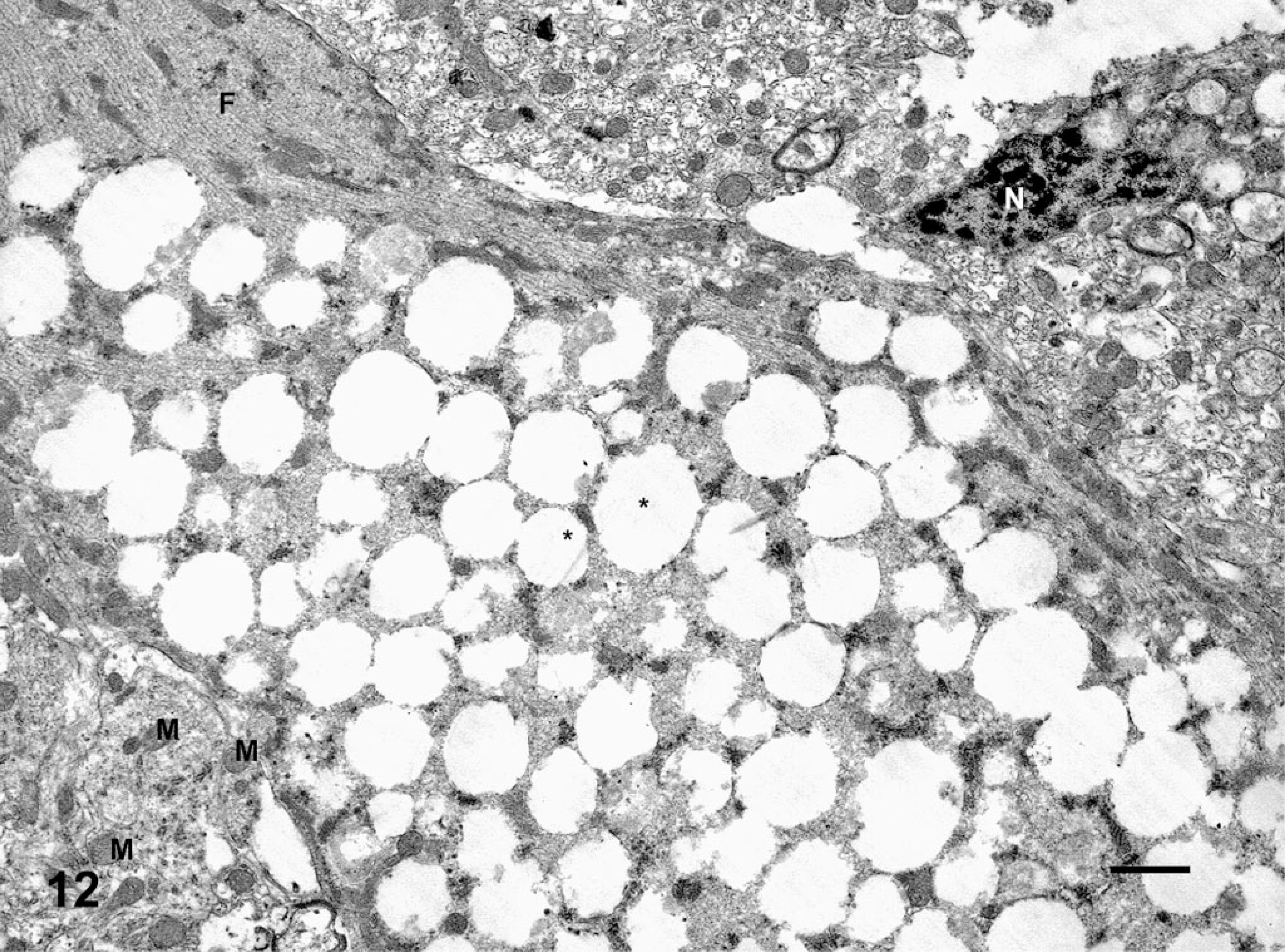
Cerebellar Purkinje cell; deer No. 6947. Fed a locoweed ration for 180 days. This Purkinje cell is swollen and it contains numerous membrane-bound cytoplasmic vacuoles (∗), mitochondria (M), neurofilament (F), and nuclear profile (N). Bar = 100 Å.
Discussion
As seen in other species, after 6–8 weeks of locoweed consumption, the poisoned deer in this study lost weight and became ill-kept. 12, 20–22, 26 Locoweed-associated weight loss and decreased production have been attributed to neurologic anorexia, depression, and loss of neuromuscular ability to find, prehend, and masticate feed. 11, 15 Additionally, locoweed alters glycosylation of glycoprotein, enzymes, and hormones, including many intestinal glycosidases that are likely to reduce metabolic efficiency. 2, 10, 17 Nutritionally, locoweed is comparable to alfalfa, and it is reported to be more nutritious than many similarly available forbs and grasses. 14 This suggests that the weight loss and loss of condition is a direct result of swainsonine toxicosis. Both the weight loss and remarkable changes in coat condition (Fig. 1) are similar to deer with CWD. Certainly deer in such condition should be tested for both diseases.
Locoweed-poisoned deer were depressed and reluctant to move, but they were relatively normal neurologically. When startled, they were able to run and even jump with little noticeable deficit. Only with close examination, when the deer were undisturbed and moving slowly, were the minimal intention tremors and mild proprioceptive deficits discernible. This is much different from the clinical course in other species. For example sheep, cattle, and horses develop noticeable neurologic deficits within 30 days of locoweed ingestion. 21 Cattle and sheep are most often depressed and reluctant to move. Horses are also reluctant to move, but often they react anxiously, and they can have uncontrolled fits of flight in which they can damage themselves and others. If swainsonine exposure is discontinued, many of the clinical signs can resolve; however, neurologic lesions are permanent and poisoned animals are often permanently affected and unsuitable for work. 24, 28
Within 60 days, the locoweed-poisoned deer developed a mild normocytic, normochromic, nonregenerative anemia (Table 1). Similar nonregenerative anemias have been identified in locoweed-poisoned sheep and cattle. 20, 22, 23 The other leukocytes and platelets were seemingly unchanged in number or morphology. This suggests that erythrocyte production is more easily disrupted by mannosidase-altered glycoprotein processing than the other hematopoietic cells. Lymphocyte vacuolation (Fig. 2) suggests that some immune cells are also compromised. Previous work in cattle has shown that swainsonine initially results in increased immunoglobulin production and lymphocyte blastogenesis. With prolonged intoxication, leukocyte functions, including blastogenesis, phagocytosis, and bacterial killing, are altered. 27 Lymphocyte vacuolation is commonly seen and has been reported in various species with several weeks of poisoning. 18, 27 This vacuolation was not associated with altered lymphocyte function, and it appears to persist for several weeks after poisoning is discontinued. 27
The locoweed-treated deer had significantly elevated ALP and AST activities on days 60, 120, and 180 (Table 2). On these days, the mean ALP activity in locoweed-treated deer was 131, 104, and 156. Although these activities were often different from those of control deer, they are not activities commonly associated with biliary disease (>200 U/liter in many species). In most large animals, ALP is variable and GGT is considered a better indicator of cholestatic disease. GGT was unchanged in the locoweed-treated deer. This suggests that locoweed poisoning is not cholestatic. Histologically, there was no evidence of cholestasis or biliary disease. The increased ALP activity could be a result of increased isoenzyme production or altered enzyme metabolism, including cell damage that impedes enzyme clearance. AST is a sensitive but nonspecific indicator of locoweed poisoning in many species. 20, 25, 26 Activities in these locoweed-poisoned deer was significantly higher than the activities of controls, but the activities were relatively low; they averaged about 225 U/liter. This is similar to changes we have reported in locoweed-poisoned sheep and well within the reference AST activities in our laboratory for most species. 20 In our experience, locoweed poisoning increases AST activity in rodents, cattle, and sheep. AST in locoweed-poisoned horses is variable, and often its activity is minimally increased. In deer, AST appears to be a good indicator of locoweed poisoning.
Mean serum swainsonine concentrations in the treated deer were 120, 132, and 125 ng/ml on days 60, 120, and 180, respectively (Table 2). These concentrations are slightly lower than those reported in sheep (320 ng/ml) and cattle (204 ng/ml) given similar locoweed doses. 20, 22 Serum swainsonine has been used to monitor poisoning in livestock. However, it is quickly cleared from the serum, and within several days after animals are excluded from locoweed exposure, concentrations are well below detectable concentrations. Interestingly, when animals are removed from locoweed exposure and brought into a corral, the neurologic signs become more severe and apparent. Because these animals had several days for swainsonine clearance, they often had slightly increased AST and ALP activities, but no detectable swainsonine in the serum. This limits the use of swainsonine as a diagnostic indicator of poisoning. Recent serum protein analysis has identified several proteins, including transferrin, that are abnormally glycosylated in poisoned animals. The clearance of these proteins closely reflects the clearance of transferrin, suggesting they are much longer lived and likely to be good biomarkers of poisoning. Efforts are underway to develop methods to monitor these proteins and correlate their glycosylation with poisoning.
Vacuolation of exocrine pancreas and salivary glands has been reported in both rodents and livestock. 20, 26 However, the vacuolation in these poisoned deer (Fig. 3) is remarkable. The endocrine pancreas is notably spared, with no histologically evident changes. Other glands, including the parietal glands, the esophageal glands, and the duodenal glands, have similar but less severe vacuolation. Thyroid follicular epithelium is also commonly vacuolated in locoweed poisoning. In other species, follicular vacuolation is prominent and easily distinguished from resorption vacuole hyperplasia of hyperthyroidism. The thyroid follicular vacuolation in deer is minimal and patchy and very similar to resorption vacuoles (Fig. 4). Lectin histochemistry (inset, Fig. 4) demonstrates that this vacuolation includes complex oligosaccharides.
Poisoned deer had neuronal perikaryonic swelling and cytoplasmic vacuolation typical of locoweed poisoning in other species. Vacuolation was apparent in the Purkinje cells of the cerebellum (Fig. 5). Other large neurons do not appear to be vacuolated. Interestingly, this relative lack of response is similar to that seen in poisoned rats and hamsters, which are also quite resistant to the neurologic effects of swainsonine intoxication. 26 In sheep, cattle, pigs, and horses the large neurons embedded in the cerebellar granular layers are severely vacuolated and they are probably among the first neurons lost in locoweed poisoning. 20 In these species the large neurons in the basal ganglia and the many nuclei in the medulla are mildly vacuolated. The white tracts of the cerebellar peduncles contain small numbers of swollen axons (spheroids). Locoweed poisoned deer have mild neuronal vacuolation with minimal changes in the white tracts. However there is severe axonal swelling and dystrophy in the gracilis and cuneate fasciculi and nuclei (Fig. 7). The neurons of the gracilis and cuneate nuclei are also degenerative and several have severe cytoplasmic vacuolation. This localized change appears to be unique to locoweed poisoning in deer as similar lesions have not been seen in other species, even in severe poisoning with durations of several years. The axons in the gracilis and cuneate fasciculi can be very long as most begin as distal dorsal root neurons that travel in the ascending spinal cord to the gracilis fasciculus to reach gracilis nucleus in the caudal medulla. Processes of more proximal dorsal root ganglion cells (T6 and rostral) ascend to the cuneatus fasciculus and end in the cuneatus nucleus. Axons from cells in the gracilis and the cuneatus nuclei pass ventrally as internal arcuate fibers, cross the midline and form the medial lemniscus and ultimately end in the somatosensory cortex and cerebellum. Afferent fibers from the lateral cuneate nucleus in the medulla enter the cerebellum through the inferior cerebellar peduncle and terminate in the ipsilateral cerebellar hemisphere. Most studies suggest that lesions in the gracilis fasciculus, the gracilis nucleus, the cuneatus fasciculus and the cuneatus nucleus result in proprioceptive deficits and loss of fine motor control. In these poisoned deer, the lesions could explain the intention tremors and mild proprioception deficits that we detected clinically.
Silver stains demonstrated that Purkinje cells in poisoned deer had fewer dendritic/axon cell body interactions and increased numbers of abnormal, swollen axons (torpedos and axon hillocks). The many empty Purkinje cell baskets and morphometric studies demonstrated the loss of cells (Figs. 9, 10). Both have been described in other species that are poisoned with locoweed or purified swainsonine. 20, 24, 28
Similar to other studies, lectin histochemistry demonstrated accumulation of oligosaccharide material in the cytoplasmic vacuoles (Fig. 11). 26 WGA and PWM react with terminalN-acetylglucosamine residues on oligosaccharides. Con-A binds to high-mannose polysaccharides. Positive staining with these lectins suggests that locoweed poisoning causes accumulations of incompletely metabolized oligosaccharides rich both in N-acetylglucosamine and mannose residues. As also described in other species, these locoweed vacuoles are membrane-bound cytoplasmic vacuoles (Fig. 12).
Interestingly, the distribution and severity of locoweed-induced lesions in deer is different from most livestock and more similar to rodents. Vacuolation of epithelial cells of the kidney and thyroid are minimal in deer and rodents. 26 These tissues are affected early and severely in livestock, rodents, and probably elk. 1, 20, 22 The most remarkable lesion in deer is the extensive pancreatic vacuolation. Pancreatic vacuolation is also severe in rodents. In other species, the exocrine pancreas is vacuolated, but not to this extent. The neurologic lesions in the cerebellum and basal ganglia are minimal in deer and rodents, and most neurologic lesions in these species require relatively high doses over long durations to develop clinical signs. Because locoweed populations can be cyclic and they periodically become a dominant proportion of the available plant community, it is likely that locoweed poisoning in deer is underdiagnosed. Because there are no reports of spontaneous locoweed poisoning in deer, perhaps this work will inspire wildlife pathologists to more closely examine deer from these endemic areas. Although locoweed-poisoned deer might not demonstrate the obvious neurologic signs common in other species, poisoning probably affects animal condition, reproduction, and well-being. Poisoned deer are probably reluctant to move and will congregate near food and water, allowing increased exposure to other infectious diseases.
Although no instances of locoweed poisoning in mule deer have been reported, there are reports of deer consuming locoweed. 9, 29 Although consumption is probably closely related to locoweed population cycles, one study reported that deer find locoweed highly palatable. 29 When locoweed populations are high, it is not uncommon for grazing livestock to consume locoweed at rates of 70–80% of their diets. 13, 15 This suggests that the exposure rates of 15% of the diet being composed of locoweed or 1.5 mg swainsonine/kg bw/day are similar to what free-ranging deer might ingest.
CWD is a transmissible spongiform encephalopathy of mule deer and elk. Similar to locoweed poisoning, the clinical signs include emaciation; weakness; excessive salivation, drinking, and urination; listlessness; hyperexcitability; and ultimately separating from the herd. Differing from locoweed poisoning, the histologic lesions include spongiform transformation of gray matter, intracytoplasmic vacuolation of neurons, neuronal degeneration and loss, astrocytic hypertrophy and hyperplasia, occurrence of amyloid plaques, and absence of significant inflammatory response. 19, 30 These histologic lesions are very different from the storage disease–like accumulation of oligosaccharides, cellular swelling, and degeneration of locoweed poisoning. None of the tissues from the deer in this study demonstrated immunoreactivity with antibodies developed to synthetic peptides that bind to cervid prion proteins. Although swainsonine-altered proteins and glycoproteins have altered function and might have a higher propensity to vary from their natural conformation, there is no evidence that locoweed alters CWD transmission or pathogenesis.
Conclusions
These findings indicate that deer are susceptible to locoweed poisoning and develop lesions similar to those seen in other species with differing severity and distribution. The histologic changes of locoweed poisoning are distinctly different from those of CWD in deer. However, because the principle clinical changes of locoweed poisoning are wasting and emaciation with minimal neurologic deficits, the clinical presentation of locoweed poisoning in deer is very similar to CWD. This suggests a difficulty in clinically differentiating the diseases, but the diagnosis is easily made with histologic and immunohistochemical studies.
Footnotes
Acknowledgements
This research was performed under the supervision and approval of the Utah State University Animal Care and Use Committee. The authors thank Ed Knoppel, Brianna Johnson, Miranda Gordon, Joyce Johnson, and Rex Probst for their expert technical assistance in these studies. We also thank Drs. Jeffery O. Hall, Anthony Knight, and Dan Gould for their constructive criticism and review of this manuscript.