
Abstract
Glioblastoma multiforme is the most malignant astrocytic neoplasm and the most common brain neoplasm of humans. Spontaneous neoplasms of the brain are rare in nonhuman primates. This report describes three glioblastomas in adult captive-reared baboons. The animals exhibited a range of clinical signs, including depression, weight loss, weakness, and blindness. All three neoplasms were located in the cerebrum, with extension into the pons in one case. Histologically, the tumors were similar and were characterized by cellular pleomorphism, multinucleated cells, areas of necrosis, microvascular proliferation (glomeruloid bodies), and palisading of neoplastic cells around blood vessels and areas of necrosis. Two baboons exhibited gemistocytic differentiation, and in one baboon, the neoplastic cells were predominantly spindle shaped with a fascicular growth pattern. Immunohistochemical staining for glial fibrillary acidic protein, vimentin, and S-100 protein was positive, whereas immunostaining for synaptophysin and chromogranin A was negative. Positive staining for the cell proliferation marker Ki67 ranged from 8.2% to 13.9%. Terminal deoxynucleotidyl transferase mediated dVTPnick end labeling (TUNEL) staining ranged from 1.8% to 5.7%. These baboon glioblastomas share many features with those of humans.
Brain neoplasms have been rarely reported in nonhuman primates. Most publications describe tumors that were experimentally induced and of neuroepithelial origin. Both astrocytomas and oligodendrogliomas have been reported in macaques coinfected with simian virus 40 and simian immunodeficiency virus. 4,10 Astrocytomas have also occurred in owl monkeys inoculated with human JC virus. 15 Glioblastoma multiforme has been associated with experimental exposure to radiation of the brain 5,6,14,20 and spinal cord 16 of rhesus macaques. Spontaneous neuroepithelial tumors described in the literature include an astrocytoma in a cynomolgus macaque, 23 a neurohypophyseal astrocytoma in a rhesus macaque, 9 a medulloblastoma in a baboon, 1 and a glioblastoma multiforme in a baboon. 7 Here, we describe the pathologic features of three cases of spontaneous glioblastoma multiforme in baboons.
These cases involve baboons that were housed at the Southwest National Primate Research Center/Southwest Foundation for Biomedical Research (SNPRC/SFBR) in San Antonio, Texas. In all three cases, tissue samples were fixed in 10% neutral buffered formalin, processed routinely, embedded in paraffin, sectioned at 5 µm, and stained with hematoxylin and eosin. Selected sections were stained using the periodic acid–Schiff (PAS) method. Sections of brain were stained immunohistochemically using a streptavidin–biotin–peroxidase method according to the manufacturer's instructions (Histostain SP Kit; Zymed Laboratories, San Francisco, CA). The primary antibodies used are summarized in Table 1. For chromogranin A, Envision®+ Dual Link Peroxidase system (DAKO, Carpinteria, CA) was used according to the manufacturer's instructions. The chromagen was 3,3′-diaminobenzidine, and the slides were lightly counterstained with Gill's hematoxylin. TUNEL staining was performed using the DeadEnd Colorimetric TUNEL System (Promega Corp., Madison, WI). For both Ki67 and TUNEL-stained slides, 500 nuclei per case were counted and the numbers of positive and negative cells were recorded.
Primary antibodies used in evaluation.
∗ GFAP = glial fibrillary acidic protein.
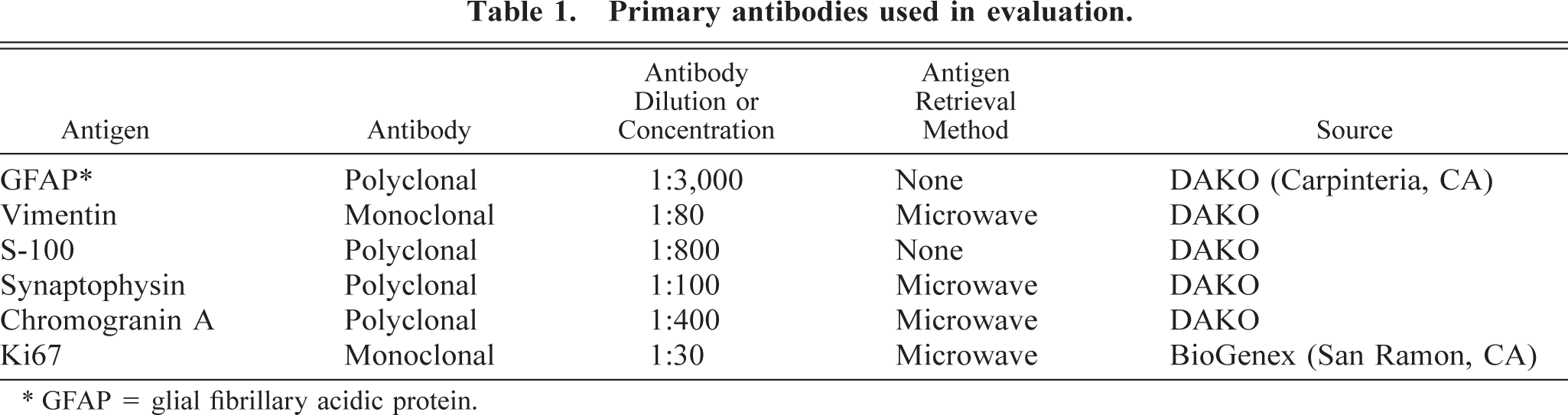
Baboon No. 1 was a 24-year-old female acquired at the age of eight from a primate importer and was a member of the breeding colony throughout her life. The animal was admitted to the hospital with a laceration on the face and was euthanatized after 11 days of worsening anorexia, weight loss, weakness, and depression. At necropsy, a soft, poorly defined mass was present in the right midcerebrum. Histologically, the neoplasm was highly infiltrative, affecting both white and gray matter. The neoplasm was composed of sheets of highly pleomorphic, round to polygonal glial cells that exhibited marked anisocytosis and anisokaryosis (Fig. 1). Tumor cells had a small to moderate amount of pale, eosinophilic, and frequently hyalinized cytoplasm. The nuclei were often located at the periphery of the cell and were irregularly ovoid, with coarsely stippled chromatin and one to three small nucleoli. Multinucleated cells were common, and some nuclei were very large, bizarrely shaped, and contained round eosinophilic inclusions (Fig. 1). In some areas, a background of slightly mucinous stroma was present. The mitotic rate was approximately one to two mitoses per 40× field, and bizarre mitotic figures were common. Multifocal areas of necrosis, which often contained abundant mineral and were occasionally serpiginous, were present (Fig. 2). Neoplastic cells occasionally palisaded around necrotic areas and blood vessels. Prominent compact tufts of capillaries (glomeruloid bodies) were present (Fig. 3), especially on the margins of necrotic areas and at the margin between neoplastic and normal tissue. These vessels had thick walls composed of two to three layers of plump cells and generally lacked a discernible lumen. Some neoplastic cells stained strongly positively for glial fibrillary acidic protein (GFAP), which highlighted a few long cytoplasmic processes (Fig. 4). Many reactive astrocytes on the margins of the neoplasm stained strongly and uniformly for GFAP. Many neoplastic cells were positive with immunohistochemical staining for vimentin and S-100 protein, whereas stains for synaptophysin and chromogranin A were negative. With Ki67, 8.2% of neoplastic cells were positive and 5.7% of neoplastic cells were positive with TUNEL staining. Other histologic findings in baboon No. 1 included mild amyloidosis in pancreatic islets, mild multifocal neutrophilic and lymphoplasmacytic tubulointerstitial nephritis, and mild multifocal myocardial necrosis.
Cerebral cortex; baboon No. 1. The neoplasm is composed of sheets of highly pleomorphic cells. Multinucleated cells (closed arrow) are common. Note the bizarrely shaped nucleus (open arrow) and intranuclear inclusions (arrowhead). HE. Bar = 64 µm.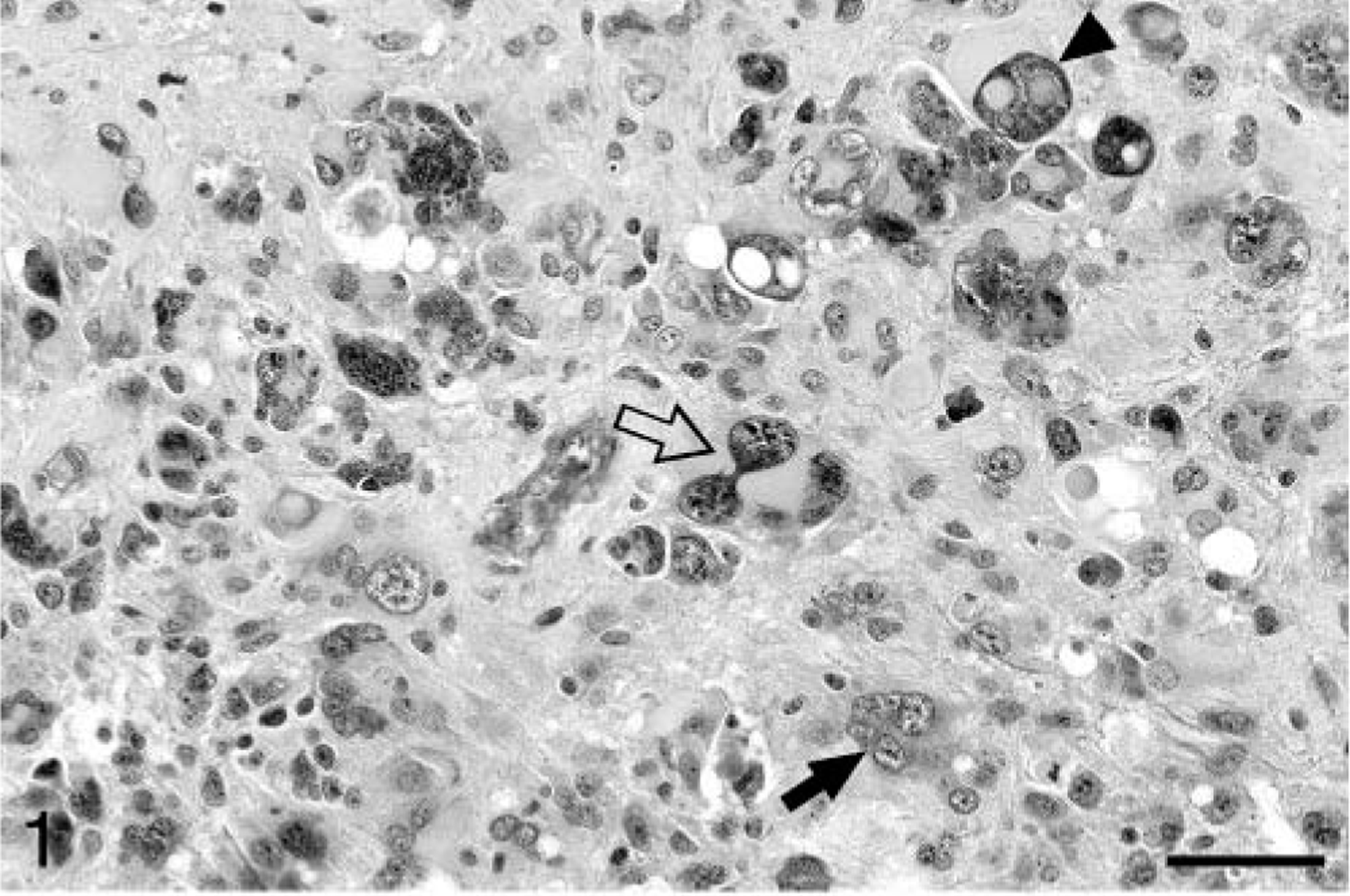
Cerebral cortex; baboon No. 1. Serpiginous areas of necrosis (asterisk) are present within the neoplasm. HE. Bar = 127 µm.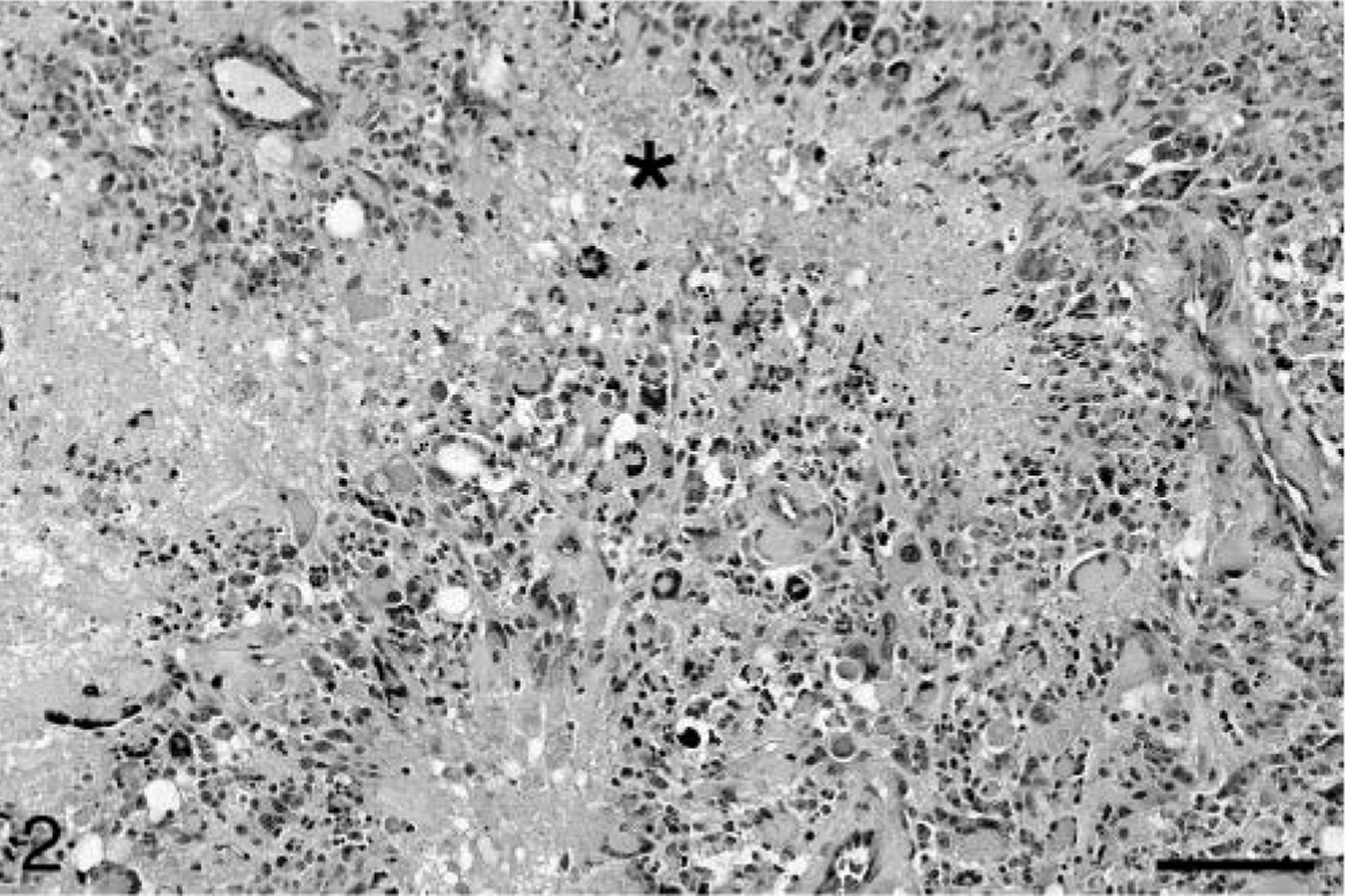
Cerebral cortex; baboon No. 1. Compact microvascular proliferation (glomeruloid bodies) are present within the neoplasm (arrows). HE. Bar = 64 µm.
Cerebral cortex; baboon No. 1. Some neoplastic cells are positive for GFAP (arrows). Cytoplasmic processes (arrowhead) are also stained. Streptavidin–biotin–peroxidase method, Gill's hematoxylin counterstain. Bar = 32 µm.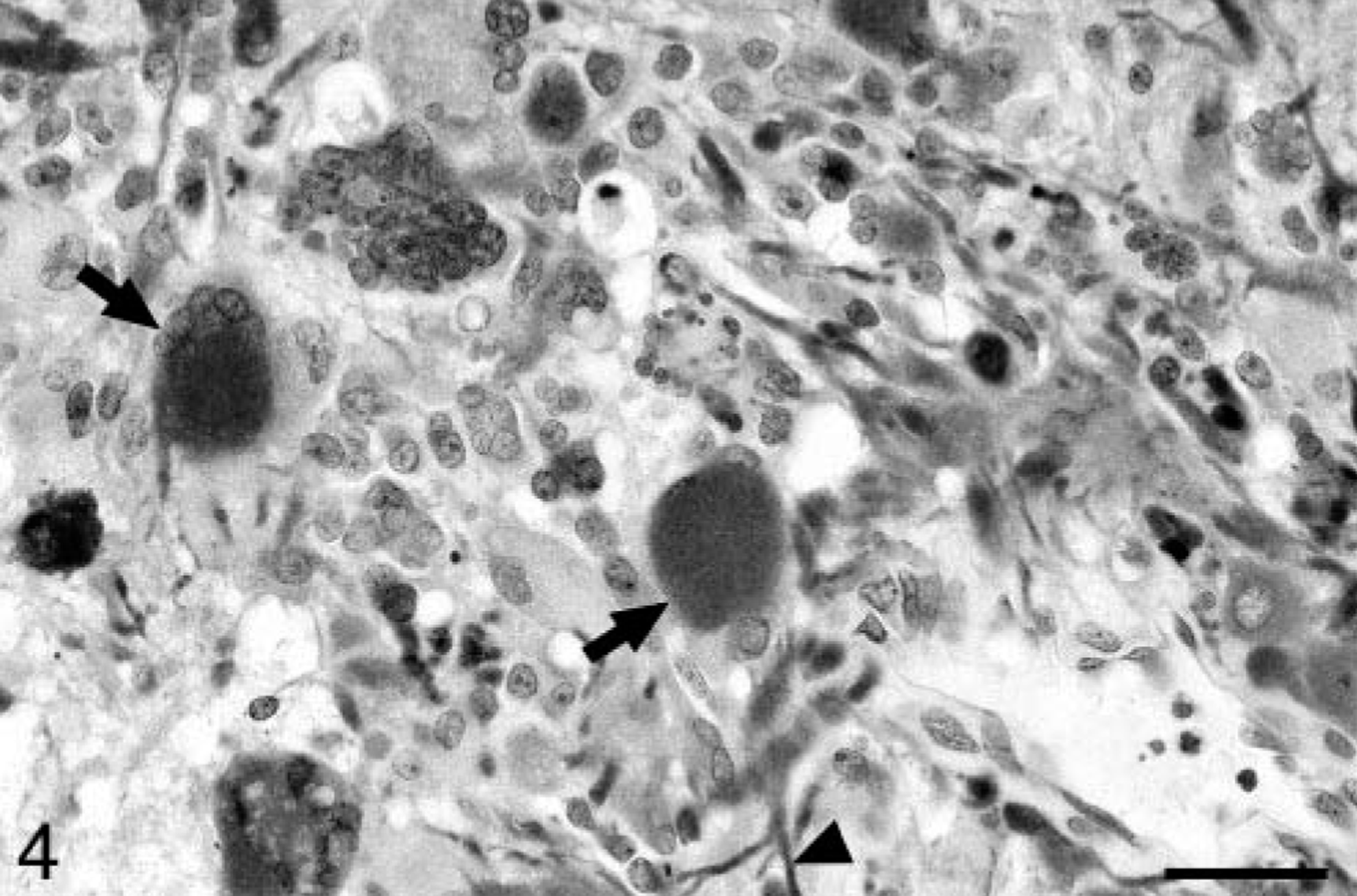
Baboon No. 2 was a 12-year-old male that was born at SNPRC/SFBR and was also a member of the breeding colony. The animal died one day after an acute onset of weakness. A complete blood count (CBC) and chemistry profile performed before death were unremarkable. At necropsy, a 2 × 3 × 2 cm soft tan area was present in the left anterior cerebrum, involving both the gray and white matter. Histologically, the brain mass was poorly demarcated and highly infiltrative, composed of irregular sheets of pleomorphic cells within a fine, fibrillary, richly vascular stroma. The majority of cells were fusiform, often forming interweaving fascicles (Fig. 5) in which long, delicate cytoplasmic processes were evident. In other areas, the cells varied from round to stellate to polygonal. The cells had a minimal to moderate amount of eosinophilic cytoplasm and ovoid to elongate nuclei, with finely stippled chromatin and occasional small single nucleoli. Marked anisocytosis and anisokaryosis were present, and multinucleated cells were common. Approximately one mitotic figure was present per 40× field, with occasional bizarre mitoses. Neoplastic cells frequently palisaded around blood vessels and small areas of necrosis (Fig. 6). Prominent reactive capillaries were present throughout the neoplasm, as well as occasional small areas of hemorrhage. GFAP immunostaining was patchy and often confined to neoplastic cells at the perimeter of the mass. Neoplastic cells stained positively for vimentin and S-100 and negatively for synaptophysin and chromogranin A. Of the neoplastic cells, 13.9% were positive with Ki67 and 1.8% were TUNEL positive. Significant lesions were not observed in the other tissues examined.
Cerebral cortex; baboon No. 2. This neoplasm is composed of spindle-shaped cells with a fascicular growth pattern. HE. Bar = 64 µm.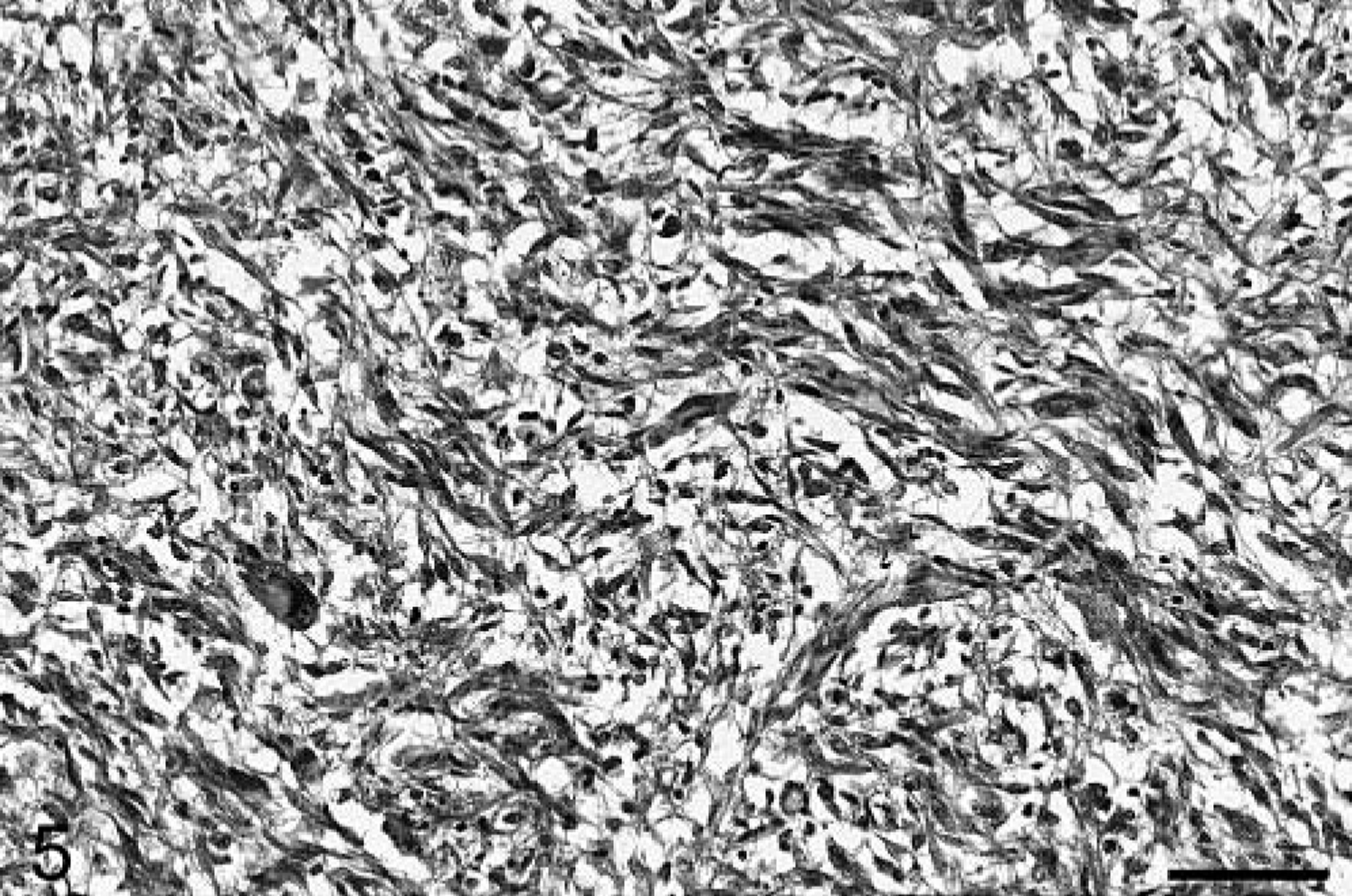
Cerebral cortex; baboon No. 2. Neoplastic cells palisade around a small area of necrosis (asterisk). HE. Bar = 64 µm.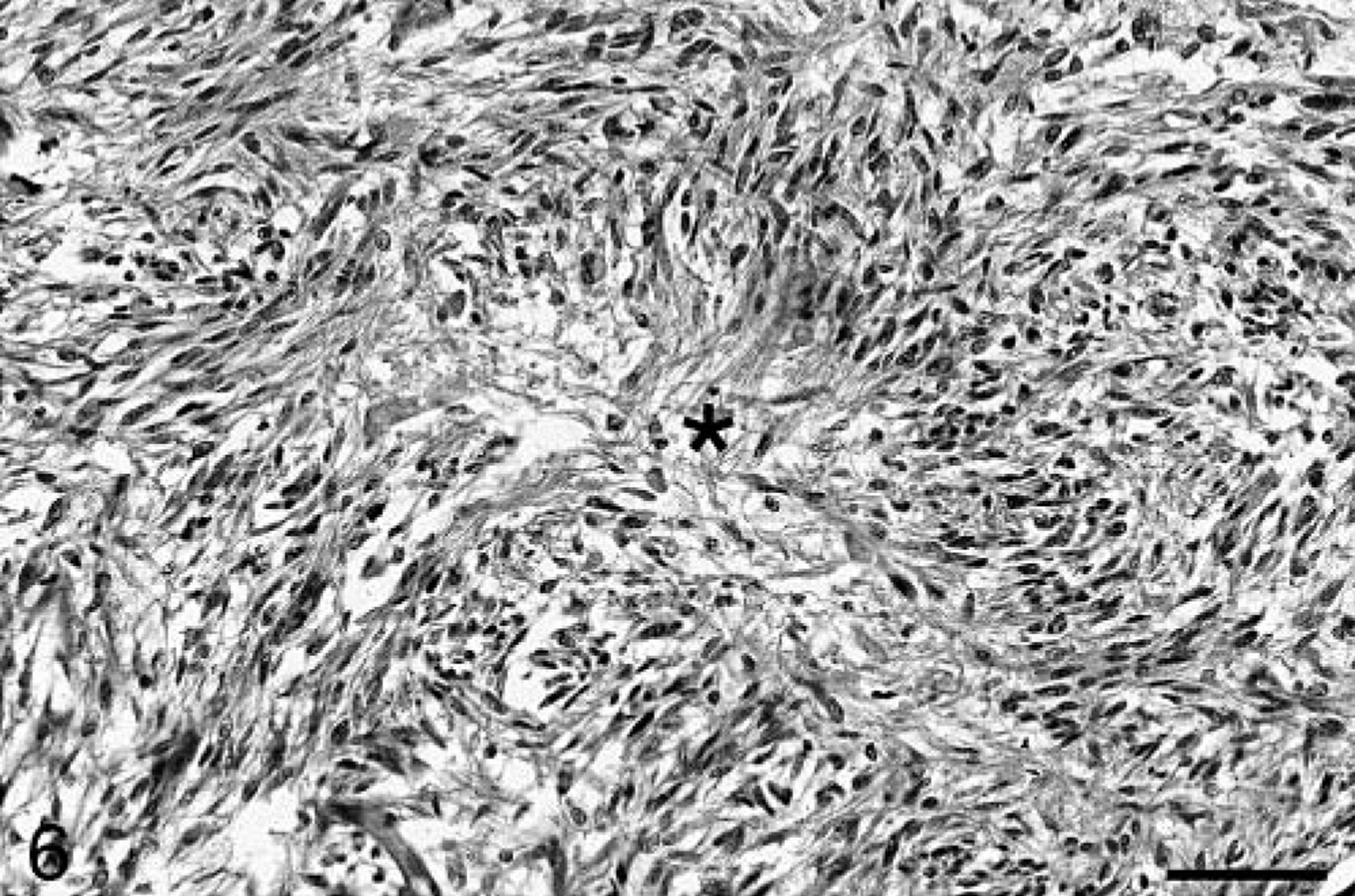
Baboon No. 3 was an 11-year-old male born at SNPRC/SFBR. The animal was euthanatized after a 4-month history of weight loss and 4 days of blindness and depression. CBC and chemistry profiles performed before death were unremarkable. At necropsy, a gelatinous, expansile, 2 cm in diameter mass was present in the midleft cerebral hemisphere, with extension into the left pons. Mild diffuse hydrocephalus was present. Histologically, the brain mass was a poorly demarcated, highly infiltrative neoplasm involving both white and gray matter of the cerebrum and extending into the lateral ventricle. The neoplasm was composed of irregular sheets of highly pleomorphic cells, with multifocal areas of slightly mucinous stroma. The cells were generally round to ovoid, with fewer fusiform cells. Most cells had a moderate to large amount of eosinophilic, hyalinized cytoplasm and irregularly ovoid, often eccentric nuclei. The nuclei had coarsely clumped chromatin and single nucleoli. Neoplastic cells exhibited marked anisocytosis and anisokaryosis, and multinucleated neoplastic cells were common, often with nuclei displaced to the margin of the cell (Fig. 7). Some nuclei contained eosinophilic inclusions. Two to three mitotic figures were present per 40× field, and abnormal mitoses were common. Small multifocal areas of hemorrhage and necrosis were present. The neoplastic cells occasionally palisaded around small blood vessels (Fig. 8). In some areas, the neoplasm had the appearance of a lower grade lesion, with cells more densely packed and having only a minimal to small amount of cytoplasm. In these areas, small pseudorosettes around blood vessels were evident. Neoplastic cells focally formed dense accumulations in the subpial area of the cerebrum and extended into the leptomeninges. Blood vessels within the tumor were often surrounded by small numbers of lymphocytes and fewer plasma cells. As with baboon No. 2, staining for GFAP was weak and multifocal. Vimentin and S-100 immunostaining were positive, and synaptophysin and chromogranin A immunostaining were negative. With Ki67 staining, 13.6% of neoplastic cells were positive, whereas 2.7% of neoplastic cells were positive with TUNEL staining. Other lesions in baboon No. 3 included minimal multifocal myocardial necrosis and mild diffuse hepatic lipidosis.
Cerebral cortex; baboon No. 3. The neoplasm is composed of sheets of highly pleomorphic cells. A large multinucleated cell with peripheralized nuclei is present (arrow). An area of necrosis is present at the upper right (asterisk). HE. Bar = 127 µm.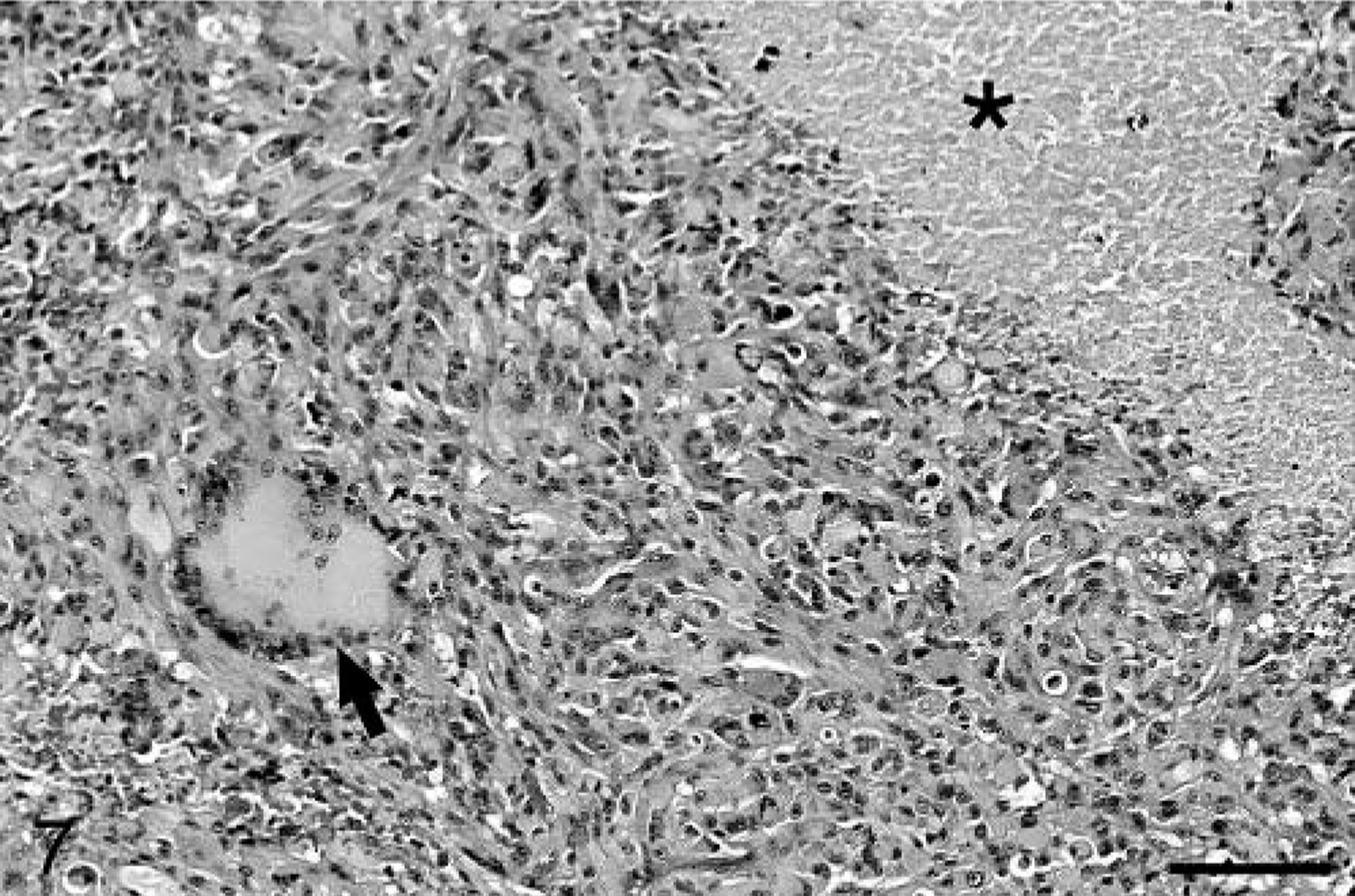
Cerebral cortex; baboon No. 3. Neoplastic cells palisade around a small blood vessel. HE. Bar = 64 µm.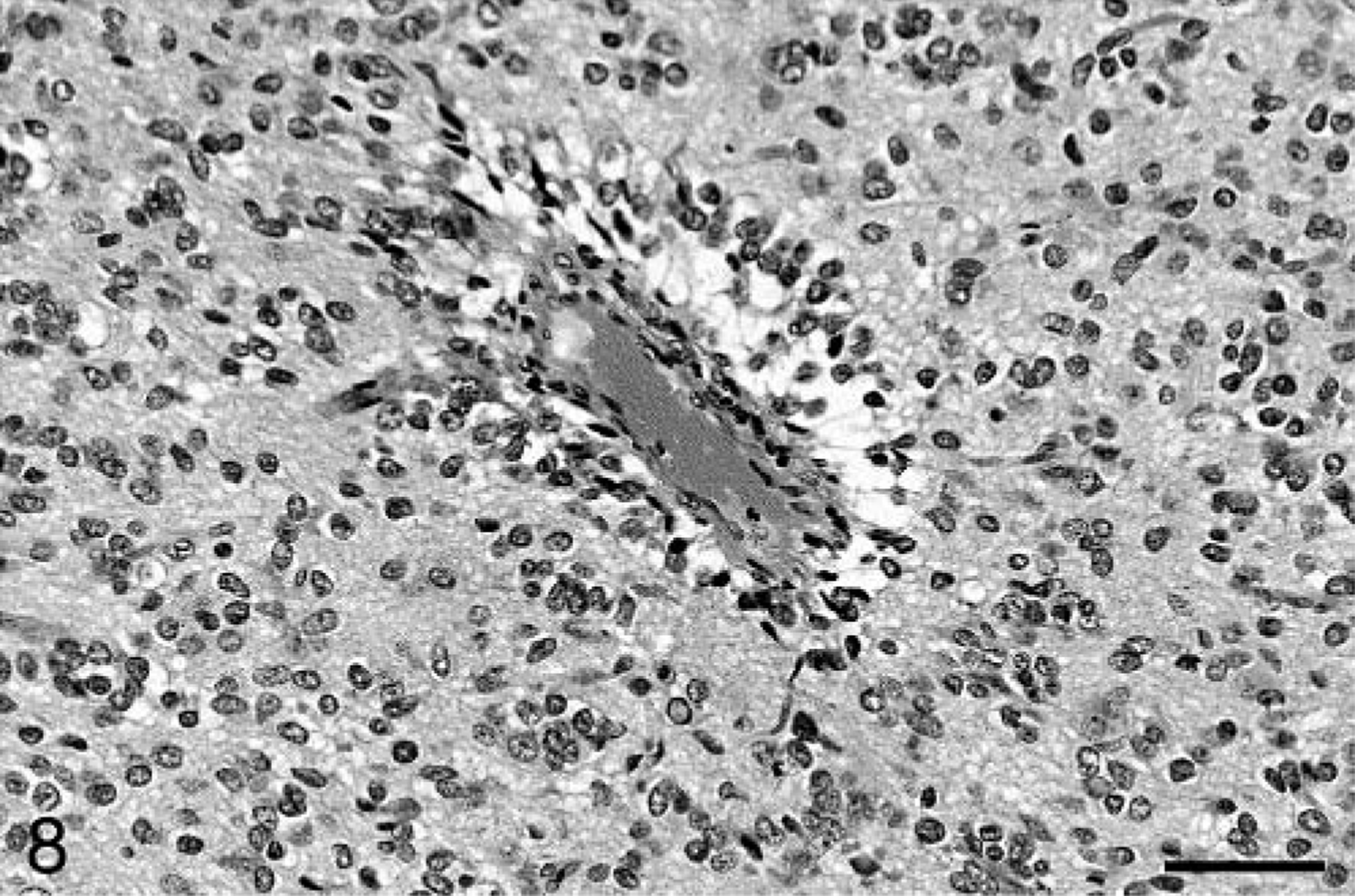
Glioblastoma multiforme is the most common brain neoplasm in humans, accounting for approximately 12–15% of all brain tumors. 3,11 The tumor is most common in adults, with a peak incidence between 45 and 70 years. 11 The neoplasm most commonly arises within the subcortical white matter of the cerebrum, and a slightly higher occurrence in males has been noted. 11 Glioblastomas are usually solitary, but multicentric tumors can occur. Glioblastomas in humans arise de novo (primary glioblastoma), or they can arise by mutation within a preexisting lower grade (less malignant) astrocytoma (secondary glioblastoma). 3 Primary glioblastomas arise rapidly in patients with a mean age of 55 years, whereas secondary glioblastomas develop more slowly in patients with a mean age of 40 years. 12
The baboon neoplasms described here shared many of the histologic features observed in human glioblastoma, including highly infiltrative growth with extension to the leptomeninges, marked cellular pleomorphism with characteristically bizarre nuclei, areas of necrosis and hemorrhage, microvascular proliferation, multinucleated cells, and pseudopalisading around blood vessels and areas of necrosis. The neoplastic cells in baboon Nos. 1 and 3 occasionally showed gemistocytic differentiation, a feature described in some human glioblastomas. Baboon No. 2 had a fascicular pattern, which is another well-recognized pattern in human glioblastomas. Characteristics occasionally seen in human glioblastomas that were not evident in the baboon tumors include lipidized tumor cells, PAS-positive granular cells, glandular or ribbonlike epithelial structures, and foci of squamous metaplasia. The intranuclear inclusions noted in baboon Nos. 1 and 3 were interpreted as cytoplasmic invaginations. The immunohistochemical findings in the baboon cases are similar to those described in human cases. 3,11 Both proliferative activity and apoptosis have been evaluated in human gliomas as potential markers of tumor grade and prognosis, with varying results. 8,11,13,17,21 Recent studies have shown correlation between lower Ki67 cell proliferation rate 8 and higher apoptotic index 13 with longer survival times in human glioblastoma patients. Differences in methodology and the limited number of baboon cases make comparisons between this study and the reported human studies problematic. One notable parallel is that in both species, apoptotic cells tend to be concentrated around areas of palisading necrosis.
The three tumors described here share many features with previously described experimental and spontaneous cases of glioblastoma multiforme in nonhuman primates. 5–7,14,16,20 The majority of reported glioblastomas have been located in the cerebrum, with fewer tumors occurring in the cerebellum, brain stem, and cervical spinal cord. Histologic features have included cellular pleomorphism, multinucleated cells, microvascular proliferation, and palisading of neoplastic cells around blood vessels and areas of necrosis. Most cases of glioblastoma have been associated with experimental radiation of rhesus macaques, with the lone spontaneous case occurring in a 15-year-old female baboon. 7 The animals in this report had no known exposure to radiation. Astrocytomas have been associated with papovavirus infection in macaques 4,10 and owl monkeys; 15 the baboons in this report were not tested for papovaviruses. In the majority of previous cases stained with GFAP, neoplastic cells were positive. 7,14,16
The molecular pathogenesis of human glioblastoma multiforme has been studied extensively. Astrocytic neoplasms have been shown to progress from a sequential accumulation of genetic alterations. Primary and secondary glioblastoma result from different sets of mutations, but the loss of chromosome 10 is thought to be a crucial factor in the development of both types of glioblastoma. 11,18 Important genes that are affected by chromosome 10 deletions include PTEN (phosphatase and tensin homolog) and DMBT1 (deleted in malignant brain tumors 1). 19 Other genes altered in primary glioblastoma include EGFR (epidermal growth factor receptor), MDM2 (human homolog of mouse double minute 2), CDKN2A (cyclin-dependent kinase inhibitor 2A), Rb (retinoblastoma), and p16. 11,18 Secondary glioblastomas often have mutations in p53, PDGF (platelet-derived growth factor), and PDGFR (platelet-derived growth factor receptor). 11
Invasion of neoplastic glial cells involves complex interactions of extracellular matrix (ECM) components, proteolytic enzymes, and tumor cell surface molecules. 22
ECM components, such as laminin, fibronectin, tenascin, collagen IV, and vitronectin, may act as substrates for neoplastic glial cell migration. 22 Invasive neoplastic glial cells may use preexisting brain ECM or secrete their own matrix for subsequent invasion.
Neoplastic cells adhere to ECM components by means of membrane receptors such as α3β1 integrin and CD44. 22 Glioblastoma cells have been shown to degrade ECM barriers by secreting enzymes such as matrix metalloproteinases 2 and 9, urokinase-type plasminogen activator, and cathepsin B. 22 Rapid growth by the neoplastic cells results in brain swelling, leading to herniation, hypoxia, and the characteristic areas of ischemic necrosis. Increased secretion of vascular endothelial growth factor by hypoxic tumor cells is the likely stimulus for the endothelial cell proliferation that forms glomeruloid bodies. 2
Almost the full range of astrocytic tumors recognized in humans has also been observed in animals. At one end of this spectrum is glioblastoma multiforme, the most malignant of glial neoplasms. Glioblastoma multiforme should be included in the differential diagnosis for cerebral hemispheric neoplasms in the adult baboon. Whether these are primary or secondary glioblastomas remains to be established; this will be facilitated in future cases by contemporary imaging modalities, such as computed tomography and magnetic resonance imaging, allowing tumor biopsy and analysis of chromosomal aberrations.
Footnotes
Acknowledgements
We thank Alexis Wenski-Roberts of the Cornell University Image Laboratory for assistance with photography, Joy Cramer and Pat Fisher for technical assistance with immunohistochemistry, and Marie Silva and Antonio Perez for anatomic pathology support.