
Abstract
Since 1982 Ehrlichia platys, now emended as Anaplasma platys, has been diagnosed in dogs from Maracaibo, Venezuela, using buffy coat smears stained with Dip Quick. Three dogs were inoculated with an A. platys strain. When parasitemia reached 60–97%, blood samples obtained from the inoculated dogs and from two naturally infected dogs were centrifuged to obtain platelet-rich plasma, which was mixed with 0.1% glutaraldehyde at 37 C for 10 minutes. Platelet pellets were fixed in 3% glutaraldehyde for 72 hours and processed for conventional transmission electron microscopy. Platelets contained pleomorphic organisms with a distinct double membrane that was not observed when the bodies were in a determinate developmental stage. There were 1–15 individual bodies included in a host cell vacuole. The organisms had an electron-lucent inner area, whereas the internal surface of their inner plasma membranes exhibited an electron-dense rough substance. In naturally infected dogs, organisms with different ultrastructural features were found inside the same platelet. Some organisms contained central dense material surrounded by a pale zone, which was in turn surrounded by a moderately dense peripheral area. Other organisms contained an eccentrically electron-dense material. The intravacuolar space appeared fully electron-lucent. Each organism usually exhibited inner fine strands. Empty structures displaying junctions with the vacuolar membrane were observed. Our results indicate that distinct ultrastructural characteristics are associated with different stages of A. platys development and may differ among A. platys strains.
Keywords
Introduction
Ehrlichia platys (now Anaplasma platys) is a small rickettsiae that appears in stained blood films as a blue intraplatelet organism within vacuoles.
ICCT can be transmitted by injection of blood containing E. platys obtained during the parasitemic period. 6 , 14–16 , 18–20 Under natural conditions, E. platys can be transmitted through the bite of an infected tick (Rhipicephalus sanguineus) or other arthropods. 4 , 18 , 20 In most previous reports, ICCT was considered a benign disease without obvious clinical signs. Therefore, E. platys is not considered a very pathogenic organism or an important disease-causing agent. 6 , 7 , 18 , 20 , 33 , 34 Nevertheless, other investigators have reported more severe clinical signs, similar in severity to those associated with E. canis infections. 2 , 15 , 35
In ICCT, cyclic parasitemia occurs followed by cyclic thrombocytopenia at 7- to 14-day intervals. 14 , 15 , 18–20 , 35 Due to cyclic parasitemia, detection of E. platys in blood smears can be difficult. E. platys inclusions or morulae are generally an incidental finding during routine blood smear examination. 8 In 1996, Bradfield et al. 7 identified E. platys morulae in only 3 of 50 blood samples that were obtained from dogs. In 1997, Harrus et al. 17 mentioned that the identification of E. platys morulae or inclusion bodies requires a systematic, careful, and time-consuming evaluation of the blood smear. In Venezuela, we routinely use this technique to detect E. platys infection. Between January 1998 and December 1999 we diagnosed 4,936 cases of E. platys infection by observing organisms in blood smears. 5 We also have reported 97% as the maximal percentage of E. platys–parasitized platelets. 4
In 1983, French and Harvey described an Indirect Fluorescent Antibody test for the detection of serum antibodies to E. platys. This test is extremely useful when parasitemia is absent or low. Arraga-Alvarado et al. 4 introduced a modification to French and Harvey's technique to prepare an E. platys antigen. Dogs from Maracaibo, Venezuela, served as donors of the infected, platelet-rich plasma (PRP). Identification of E. platys in the sample was by electron microscopy.
Transmission electron microscopy (TEM) can be used to study the ultrastructural features of this organism and to determine if organisms observed in blood smears are truly E. platys. 4 , 16 , 19 , 24 , 25 New E. platys isolates have been characterized using TEM, 24 but molecular characterization is necessary to verify the existence of new strains.
Recent genetic analyses of 16SrRNA gene, groEL, and surface protein genes have indicated that the existing taxonomic designation of some obligate intracellular bacteria that reside in vacuoles of eukaryotic cells was flawed. That is the case for E. platys that has recently been reclassified into the genus Anaplasma and renamed A. platys. 13 , 36
This article describes the ultrastructural features of E. platys (A. platys) in dogs from Maracaibo, Venezuela, emphasizing the process of binary division and morphologic differences among organisms found within the same platelet.
Materials and Methods
Since 1982, E. platys, now named A. platys, has been diagnosed in dog blood samples obtained from Maracaibo, Venezuela, that were referred to the Clinical Diagnosis Laboratory of the Faculty of Veterinary Sciences at the University of Zulia. Animal care and experimentation were carried out in accordance with all applicable institutional, local, and national guidelines.
A Doberman was taken to the Small Animal Clinic at the Faculty of Veterinary Sciences. This dog had 136,000 platelets/μl, and 98% of these platelets were infected with A. platys. Twenty milliliters of whole blood was collected in ethylenediamine-tetraacetic acid–coated tubes, and PRP was prepared and separated into five 1-ml vials, adding an equal amount of 10% glycerol (buffered with sodium phosphate, pH 7.4) to each vial. Vials were stored in liquid nitrogen.
Two Boxers and one mongrel were infected using one vial of PRP per dog. Blood parameters of these dogs were monitored for a period of 18 days in the boxers and 17 days in the mongrel. Blood counts were evaluated every other day after inoculation. Buffy coat smears were prepared using the microhematocrit method 30 to concentrate platelets and leukocytes. Smears were stained using Dip Quick Stain (Jorgensen Lab. Inc., Colorado), and platelets infected with A. platys were observed by light microscopy.
Platelet concentrates for electron microscopy were prepared on day 18 when the percentage of parasitemia reached 97 and 60% in the boxers; a platelet concentrate from the mongrel was obtained on day 17 when 63% parasitemia was observed. The percentage of parasitemia, platelet count, and other clinical observations in these dogs are included in Table 1.
Clinical and hematologic observations in dogs infected with Anaplasma platys.
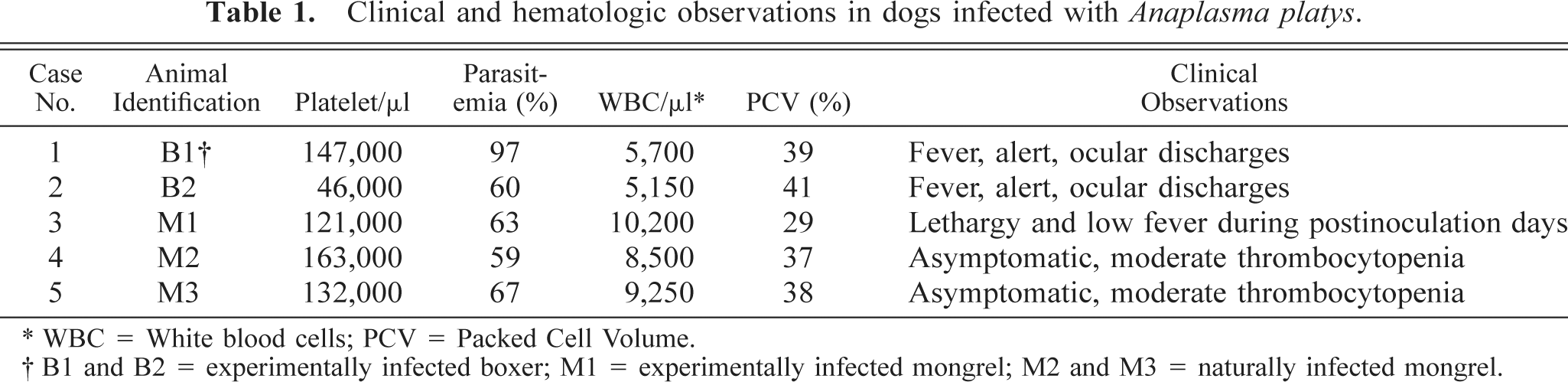
∗ WBC = White blood cells; PCV = Packed Cell Volume.
† B1 and B2 = experimentally infected boxer; M1 = experimentally infected mongrel; M2 and M3 = naturally infected mongrel.
Two other mongrels were taken to the Small Animal Clinic for a general examination. Blood counts showed high A. platys parasitemia (59–67%). These dogs represented a natural infection of A. platys. Platelet concentrates for electron microscopy were collected immediately.
All platelet concentrates were obtained from whole blood samples that were centrifuged at 103 × g for 10 minutes. The PRP was separated into several centrifuge tubes and fixed by gradually adding 0.1% glutaraldehyde solution (pH 7.3, buffered with sodium cacodylate) to a final concentration of 0.05%. The platelet suspension was incubated at 37 C for 10 minutes and then centrifuged at 1,315 × g for 10 minutes at room temperature. The supernatant was discarded and the sediment overlaid with 3% glutaraldehyde solution (pH 7.3, buffered as above) for 72 hours. The platelet concentrate was postfixed in 1% buffered osmium tetroxide for 1 hour. After being block-stained in 3% uranyl acetate, platelets were dehydrated through ascending grades of ethanol and embedded in a low-viscosity epoxy resin. Ultrathin sections were cut using a Sorvall MT-2B ultramicrotome, collected on 300-mesh copper grids, stained with lead citrate, and observed using a Hitachi H-7000 electron microscope at 80 kV. Approximately 300 blood platelets were examined from each dog.
Results
Electron microscopic examination showed the isolated platelet-rich fractions to be relatively devoid of red blood cells and leukocytes (Fig. 1). Such fractions allowed us to study the characteristics of the platelet inclusions. Platelets contained pleomorphic Anaplasma organisms that were 0.45–1.55 μm in diameter. Organisms ranged in shape from round to oval (Fig. 2) to bean shaped (Fig. 3). Each organism appeared to be bound by inner and outer plasma membranes (Fig. 4). The membranes were parallel to each other. This double membrane was not observed when the bodies were in an initial developmental stage. In this case, a single, thickened membrane was observed.
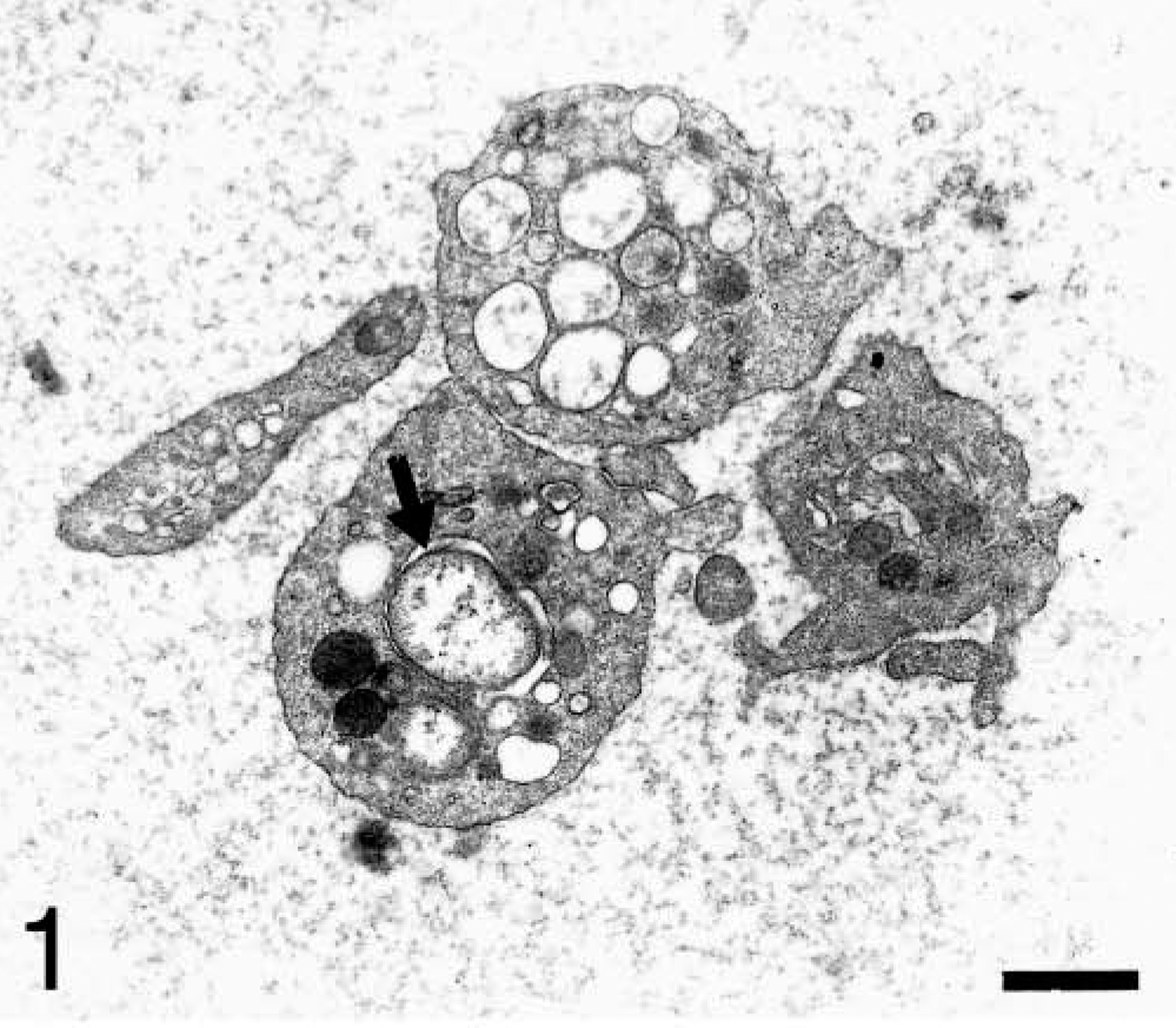
Blood platelet fraction from an experimentally infected dog; dog No. B1, case No. 1. Platelet contains an anaplasmal organism (arrow). Bar = 1 μm.
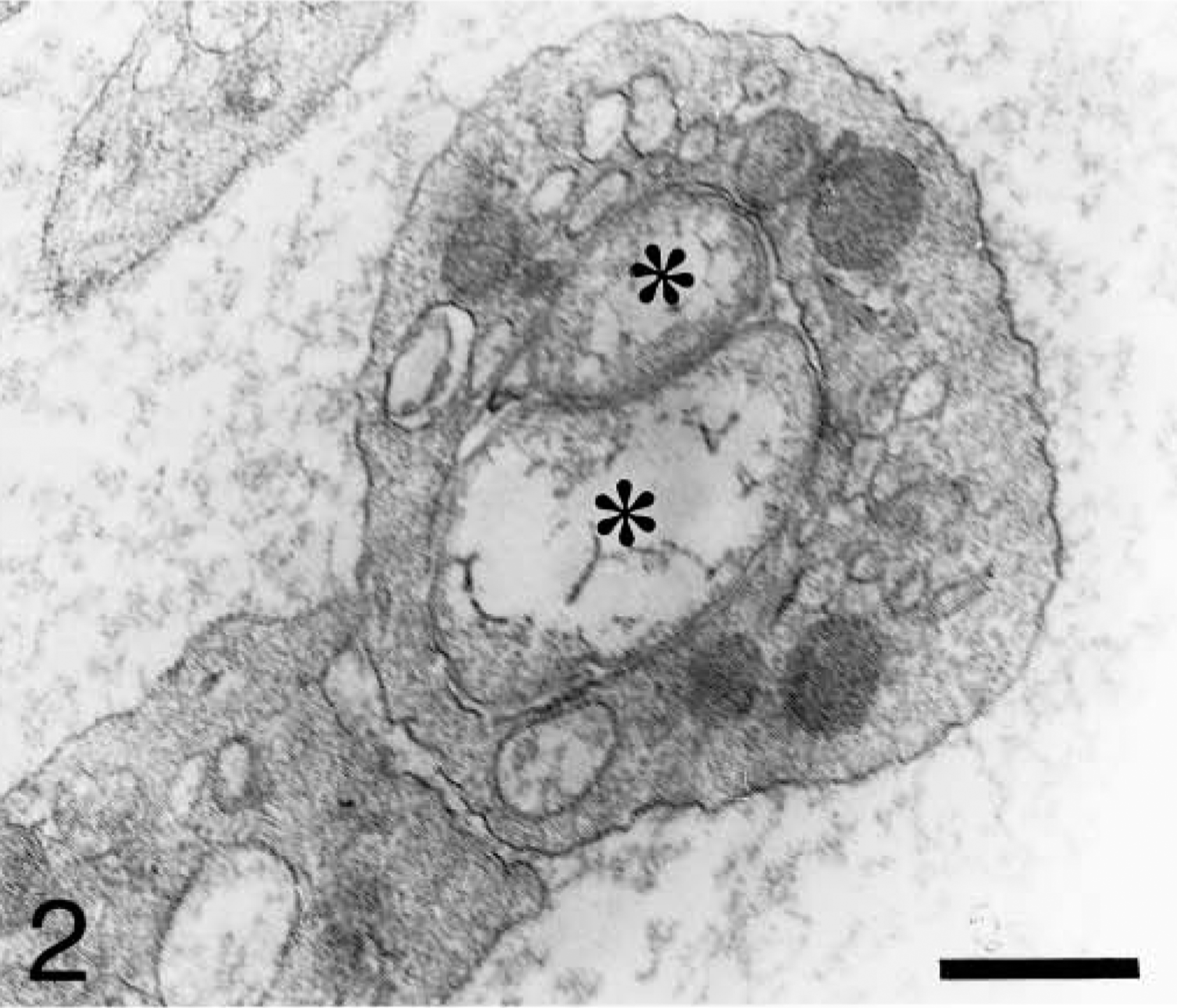
Blood platelet from an experimentally infected dog; dog No. M1, case No. 3. Two pleomorphic organisms (asterisks) are inside a vacuole. Bar = 0.5 μm.
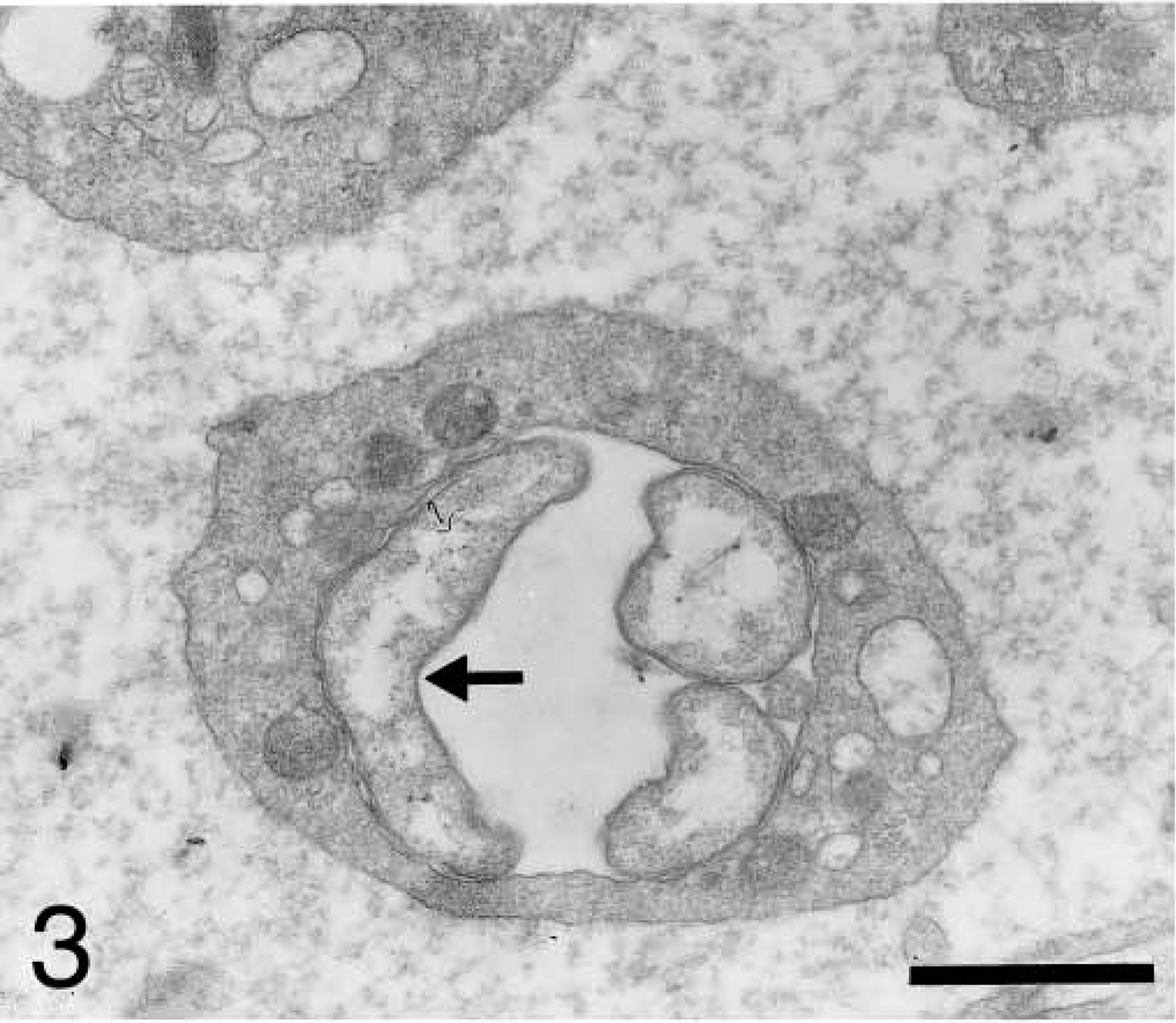
Blood platelet from an experimentally infected dog; dog No. B2, case No. 2. Platelet shows three anaplasmal bodies. Division transforms a single unit into two units and usually occurs in bean-shaped organisms (arrow). Bar = 1 μm.
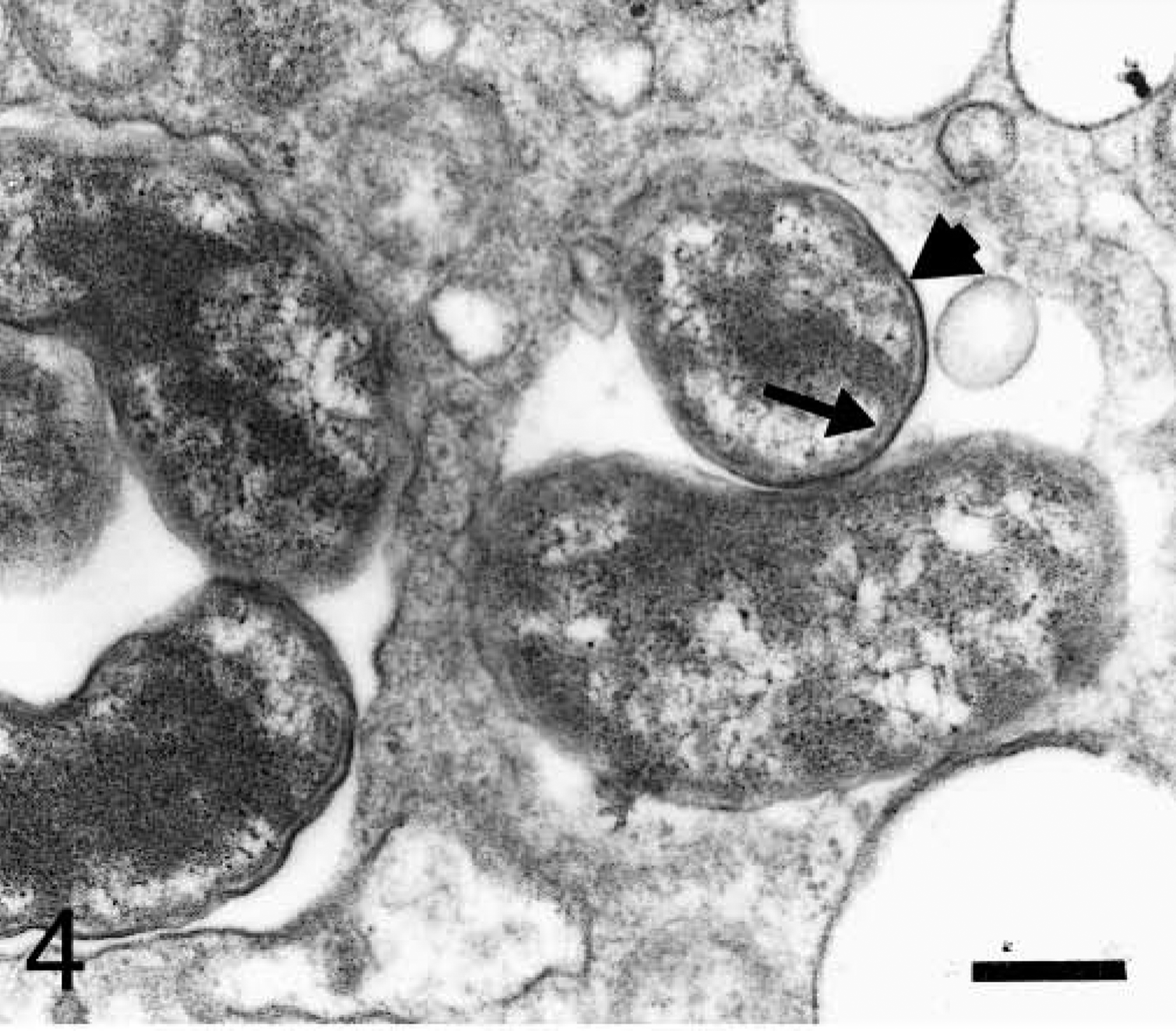
Section of blood platelet from a naturally infected dog; dog No. M3, case No. 5. Anaplasmal organisms are inside two lucent vacuoles. Observe the organisms bounded by inner (arrow) and outer (short arrow) plasma membranes. Bar = 0.25 μm.
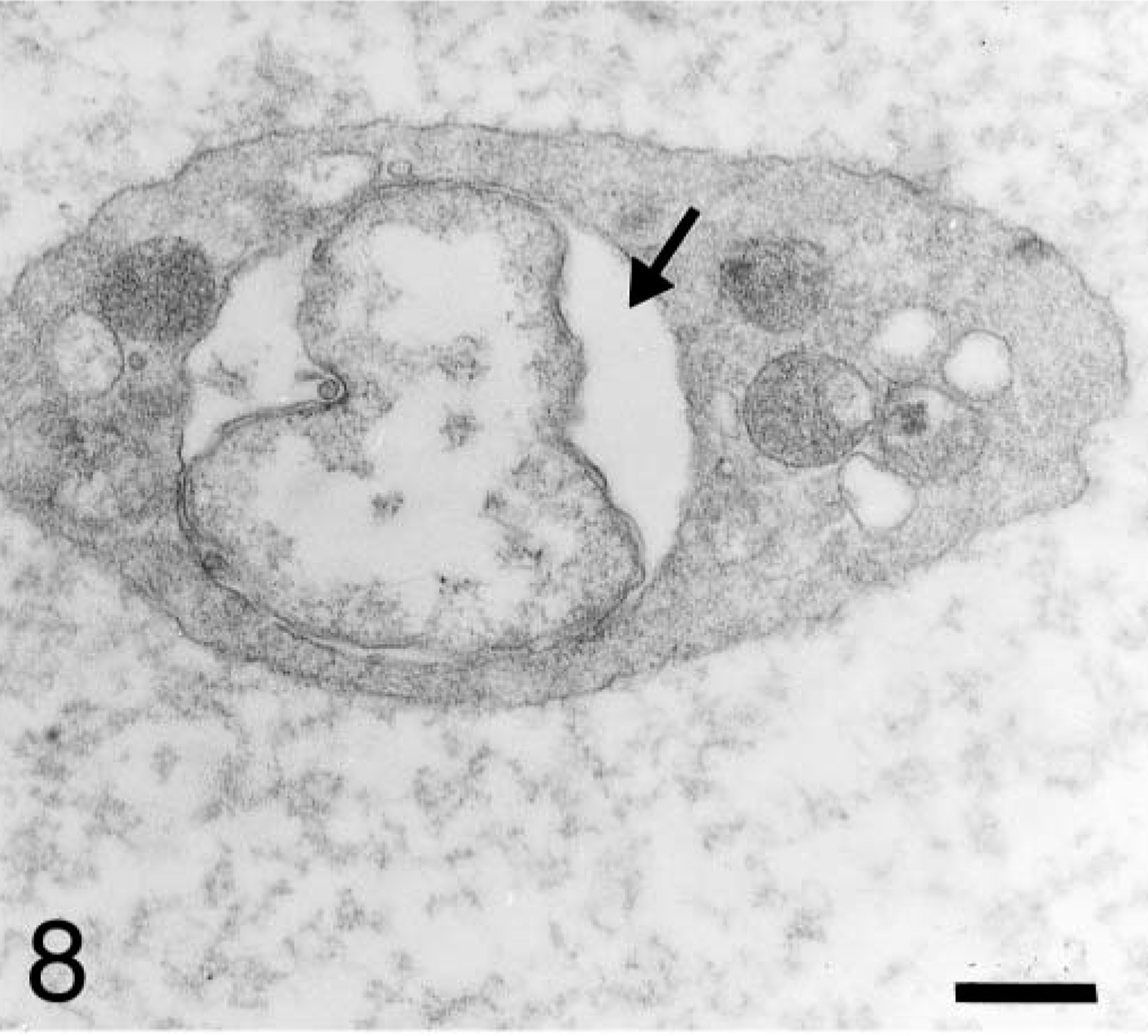
The number of organisms per host cell vacuole varied from 1 to 15. Electron-lucent, ring-shaped structures displaying electron-dense connections with the vacuole membrane were rarely observed outside the vacuole. Sites at which intravacuolar anaplasmal organisms made contact with platelet cytoplasm were observed (Fig. 5). Anaplasmal bodies, either alone or constituting a morulae, were included in a vacuole that occupied a large portion of the platelet cytoplasm (Fig. 6). Sometimes, more than one vacuole was observed within a platelet (Fig. 7). Vacuoles containing organisms were of different sizes (0.8–2.1 μm in diameter). Most of these vacuoles contained electron-lucent matrices (Fig. 8), but other vacuoles had an electron-dense fibrillar matrix between the anaplasmal organisms. Frequently, platelet mitochondria were concentrated around vacuoles containing ehrlichial organisms and formed contacts with vacuole membranes. Rarely, mitochondria were observed establishing connections with the organism itself (Fig. 9).
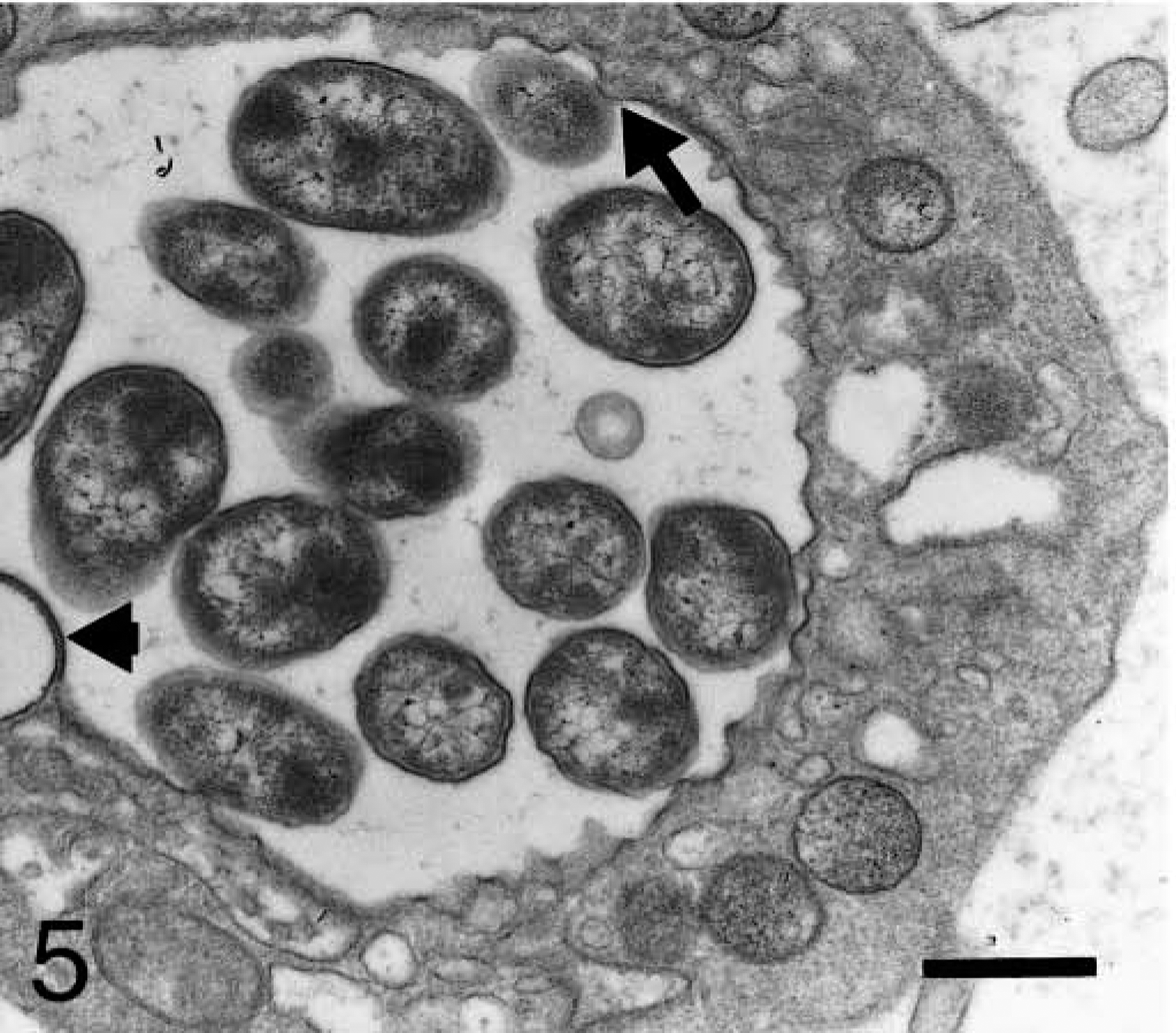
Blood platelet from a naturally infected dog; dog No. M2, case No. 4. Platelet displays a vacuole with numerous individual bodies; some of them are making contact with platelet cytoplasm (arrow). Note electron-lucent ring-shaped structures displaying electron-dense connection (short arrow) with the vacuole membrane. Bar = 0.5 μm.
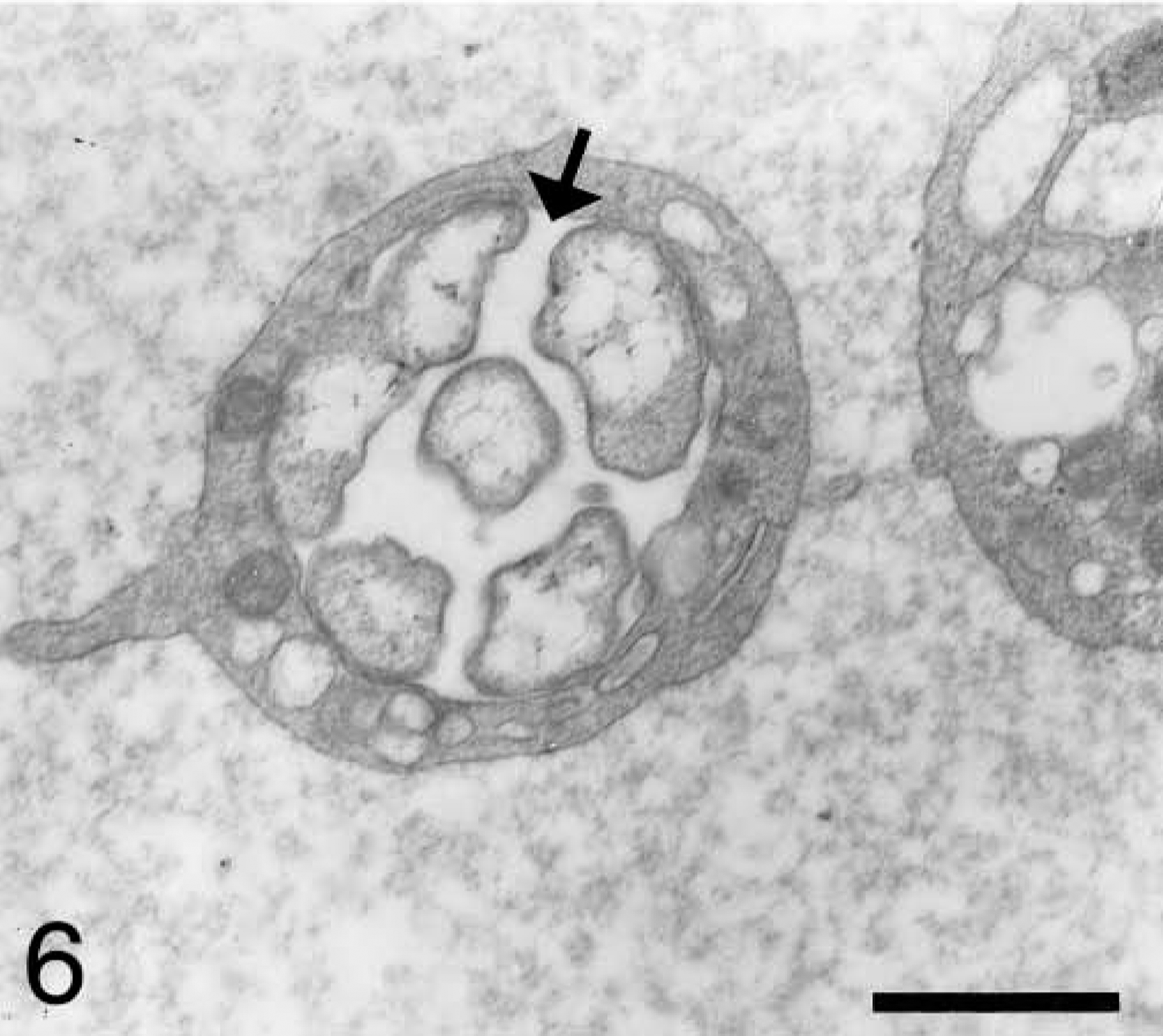
Blood platelet from an experimentally infected dog; dog No. B2, case No. 2. Platelet shows a morula (arrow) with several organisms, which is occupying a large surface of platelet. Bar = 1 μm.
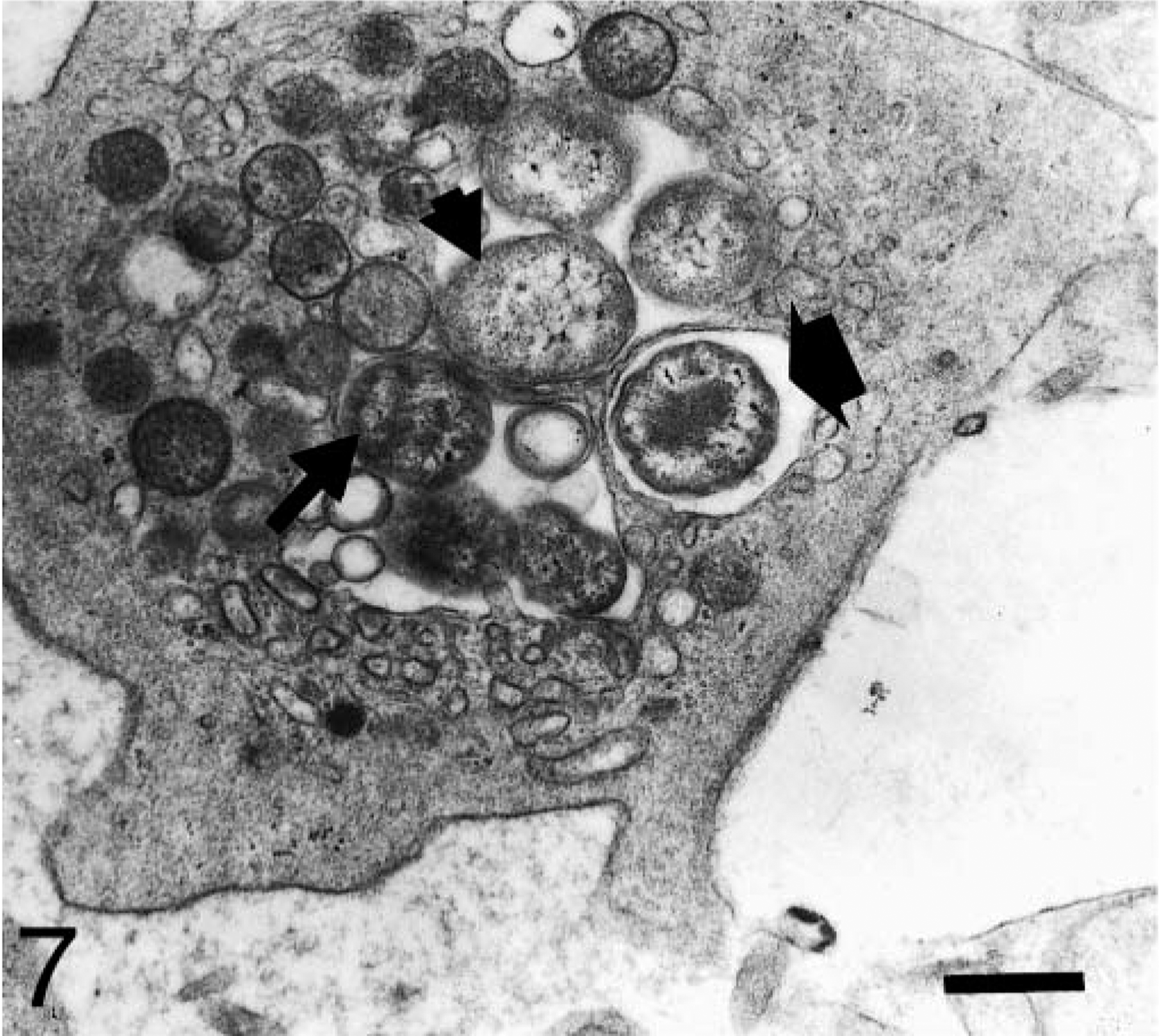
Blood platelet from a naturally infected dog; dog No. M3, case No. 5. Platelet shows several organisms with distinct ultrastructure. Observe initial bodies with a thickened membrane and a moderately electron-dense protoplasm (long arrow). Condensation of dense material in the central zone of other organisms is also observed (thick arrow). An electron-lucent central area and a peripheral electron-dense granular substance are observed in mature A. platys bodies (short arrow). Bar = 0.5 μm.
Blood platelet from an experimentally infected dog; dog No. B2, case No. 2. Platelet contains an organism inside a vacuole with electron-lucent matrix (arrow). Bar = 0.5 μm.
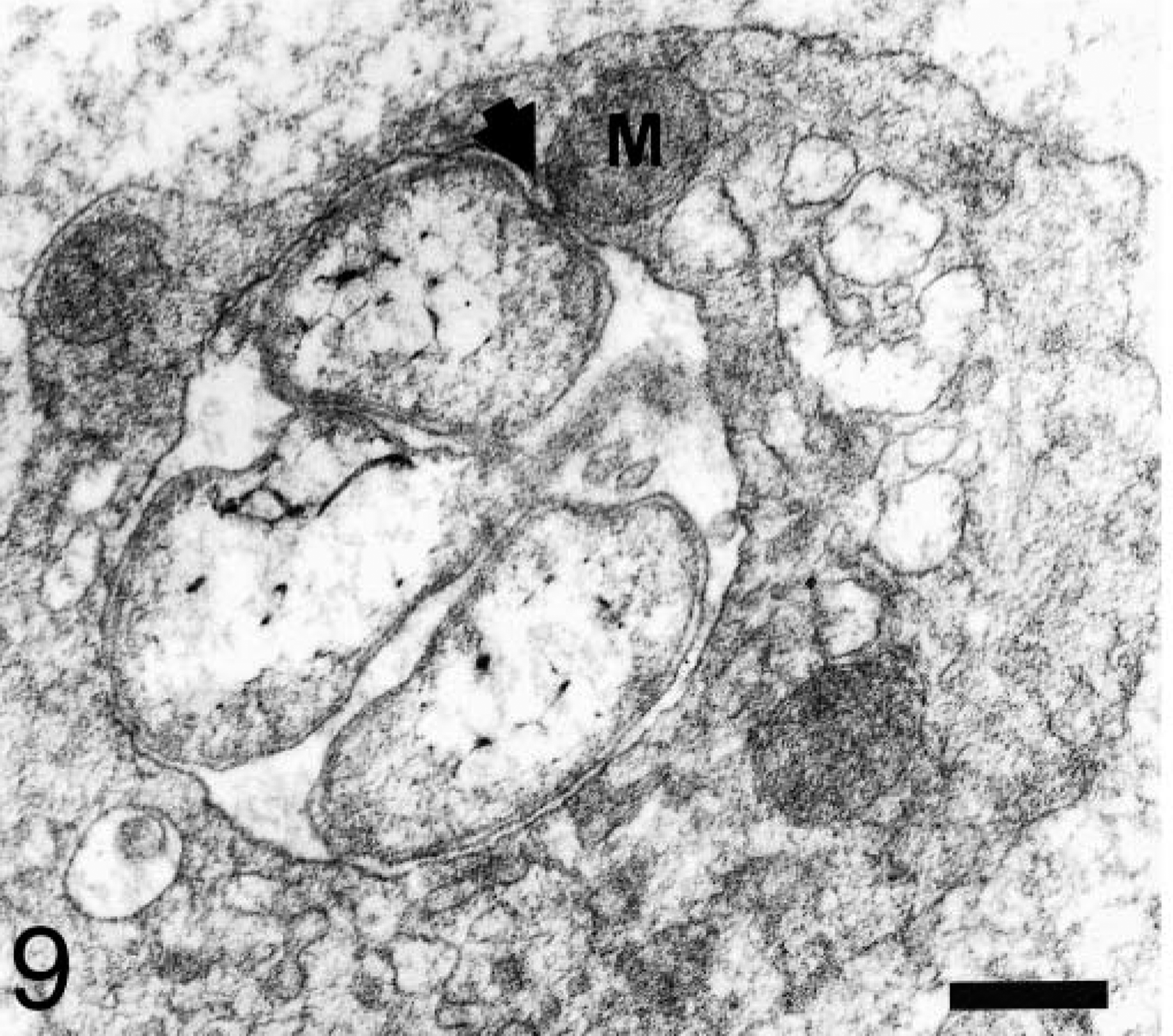
Blood platelet from an experimentally infected dog; dog No. M1, case No. 3. Platelet mitochondrion (M) is in contact (short arrow) with the vacuole membrane containing an anaplasmal organism. Bar = 0.2 μm.
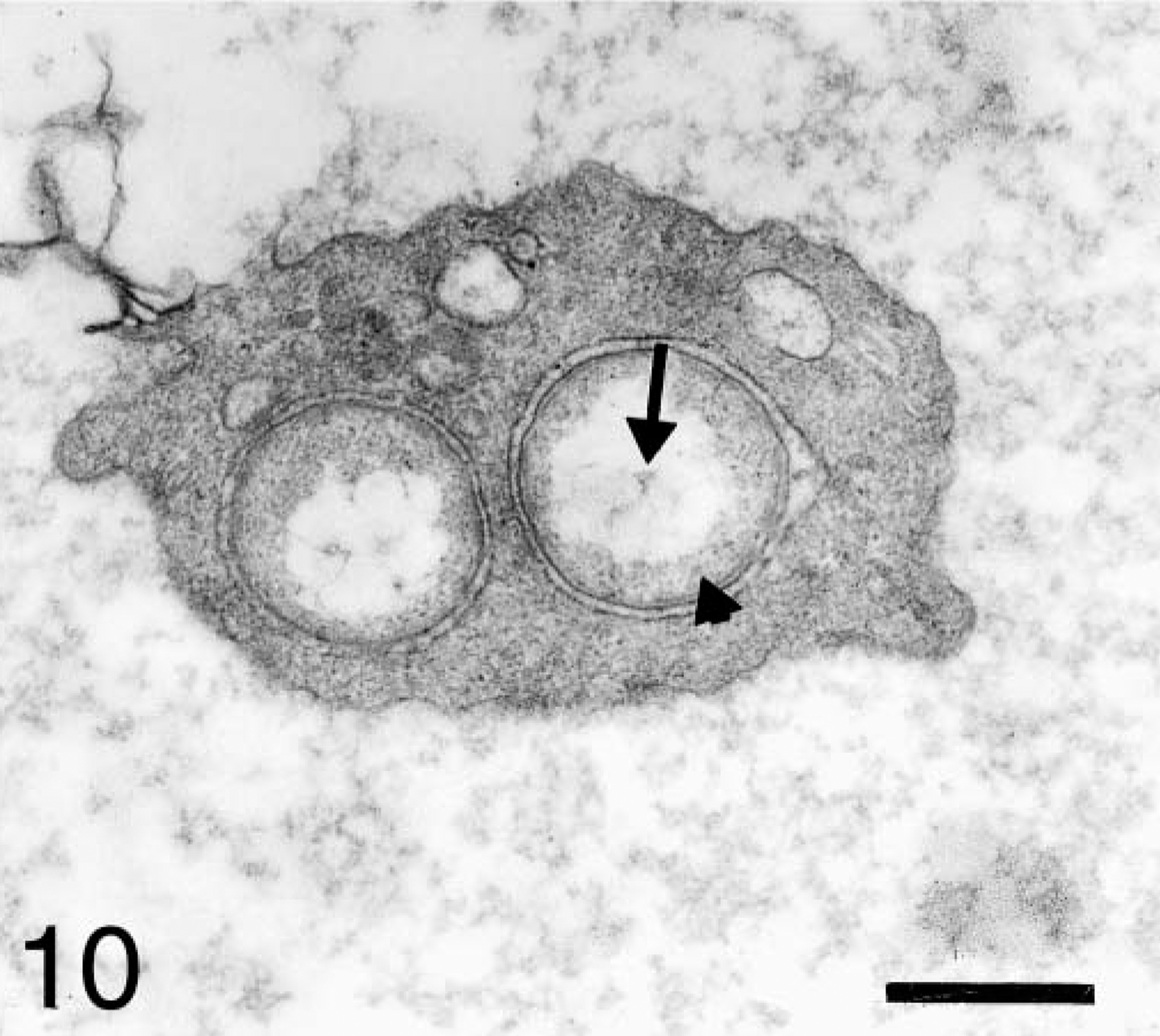
In experimentally infected dogs, all organisms had an electron-lucent central area, and the internal surface of their inner plasma membranes was coated with an electron-dense amorphous substance (Fig. 10). In one naturally infected dog, organisms within the same platelet had variable morphology. Some organisms were similar to those described above. Others had a central dense core surrounded by a pale zone, which was in turn surrounded by a moderately dense peripheral area (Fig. 11). In the other naturally infected dog, organisms had eccentrically located electron-dense material.
Blood platelet from an experimentally infected dog; dog No. B2, case No. 2. Platelet displays two organisms with electron-lucent central area (arrow) and a rough electron-dense substance in their periphery (short arrow). Bar = 0.5 μm.
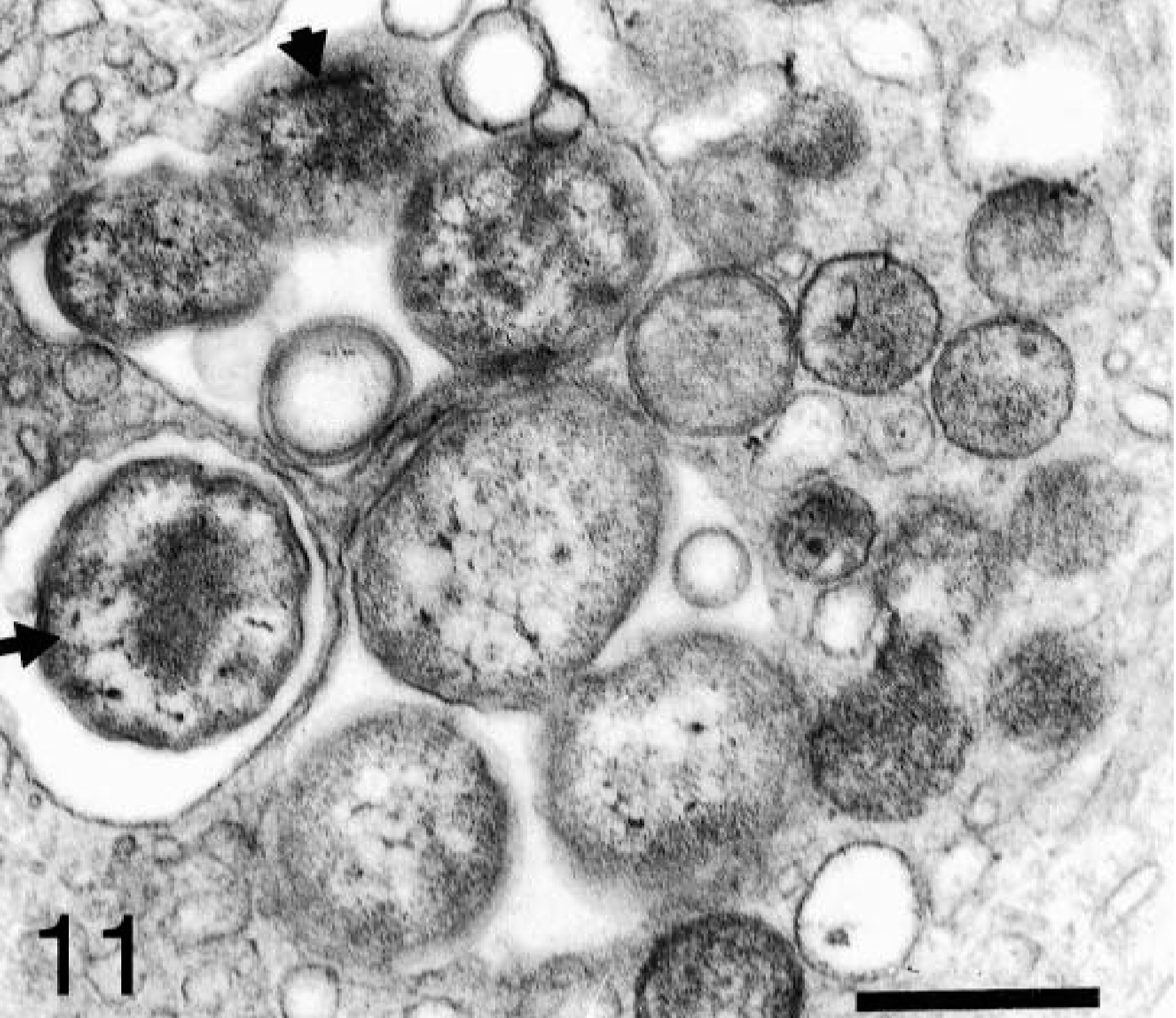
Section of blood platelet from a naturally infected dog; dog No. M3, case No. 5. Anaplasmal organisms with different ultrastructural features are observed inside the same platelet. Some organisms have a central dense material (arrow) surrounded by a pale zone; others exhibit dispersed electron-dense material (short arrow). Bar = 0.5 μm.
Most A. platys organisms contained fine fibrils near the central region (Fig. 12). In some platelets, binary fission of the anaplasmal organisms was observed. Large anaplasmal organisms were found dividing unequally (Fig. 13). Tubular and vesicular structures of small size (0.1 and 0.06 μm, respectively), and an electron-dense fibrillar matrix and a thickened membrane were observed inside the host vacuoles (Fig. 14).
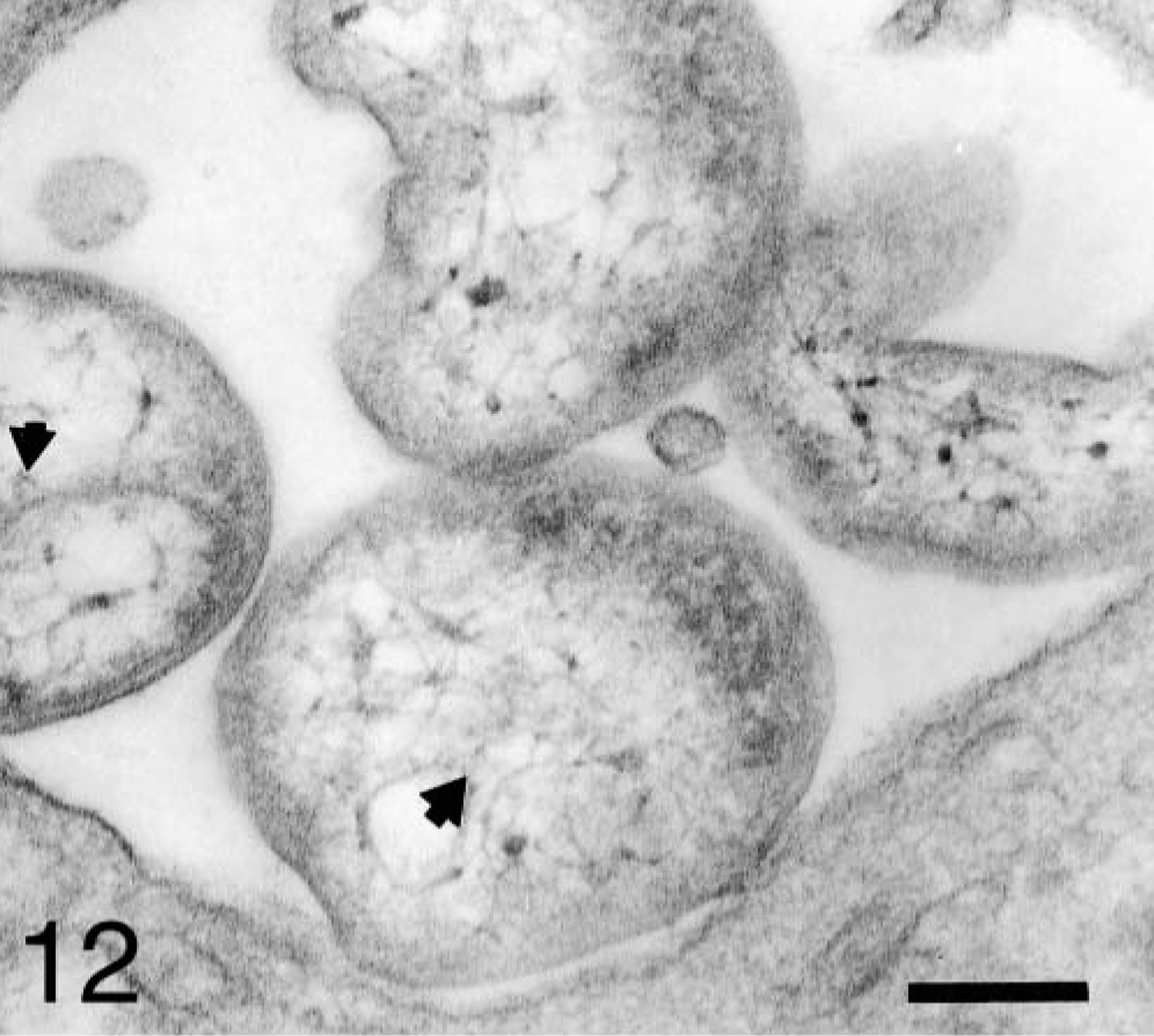
Platelet vacuole from a naturally infected dog; dog No. M2, case No. 4. Four A. platys organisms exhibit a reticular structure in their central region (short arrows). Bar = 0.2 μm.
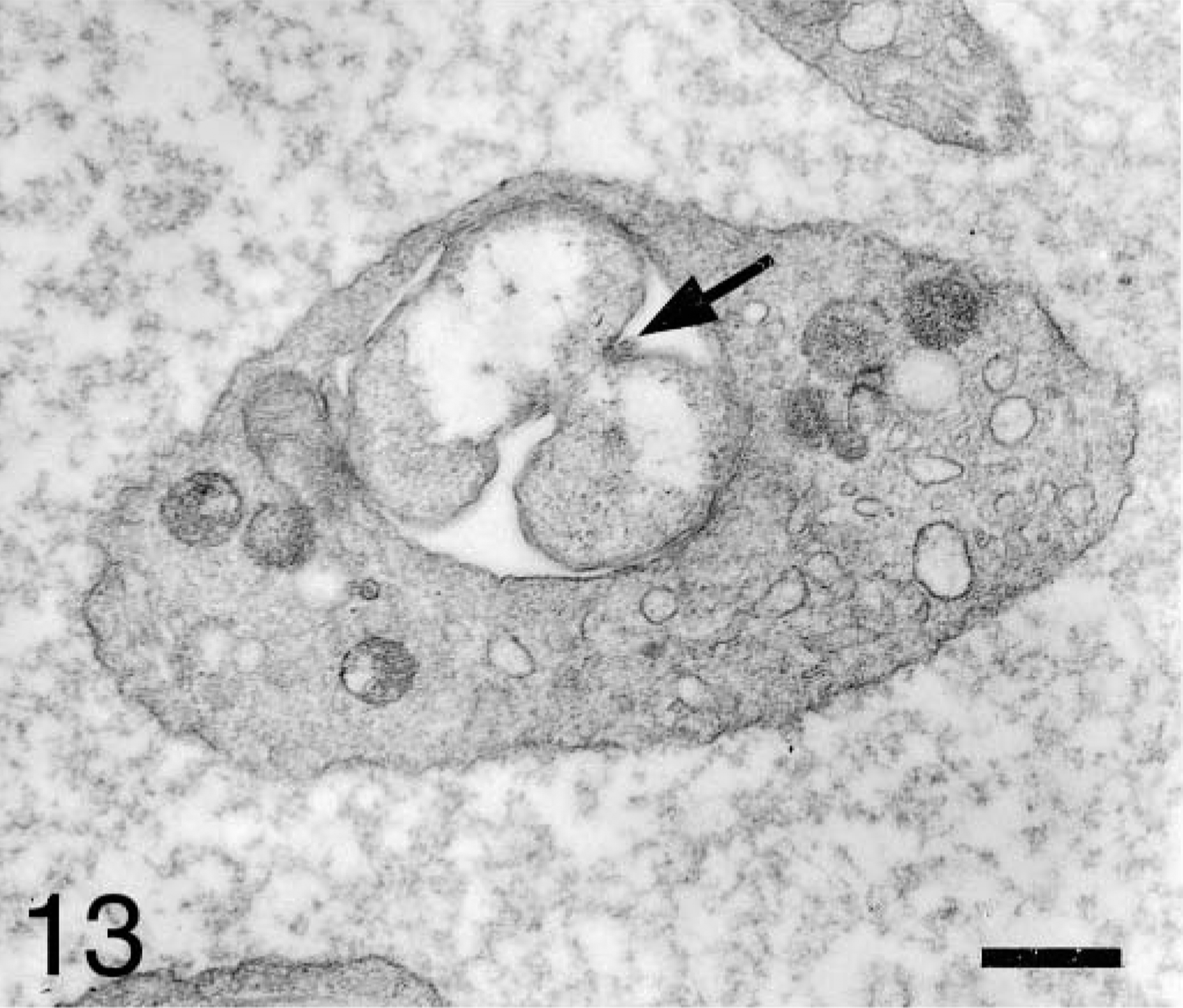
Blood platelet from an experimentally infected dog; dog No. B2, case No. 2. Platelet shows unequal division (arrow) of an anaplasmal body. Bar = 0.5 μm.
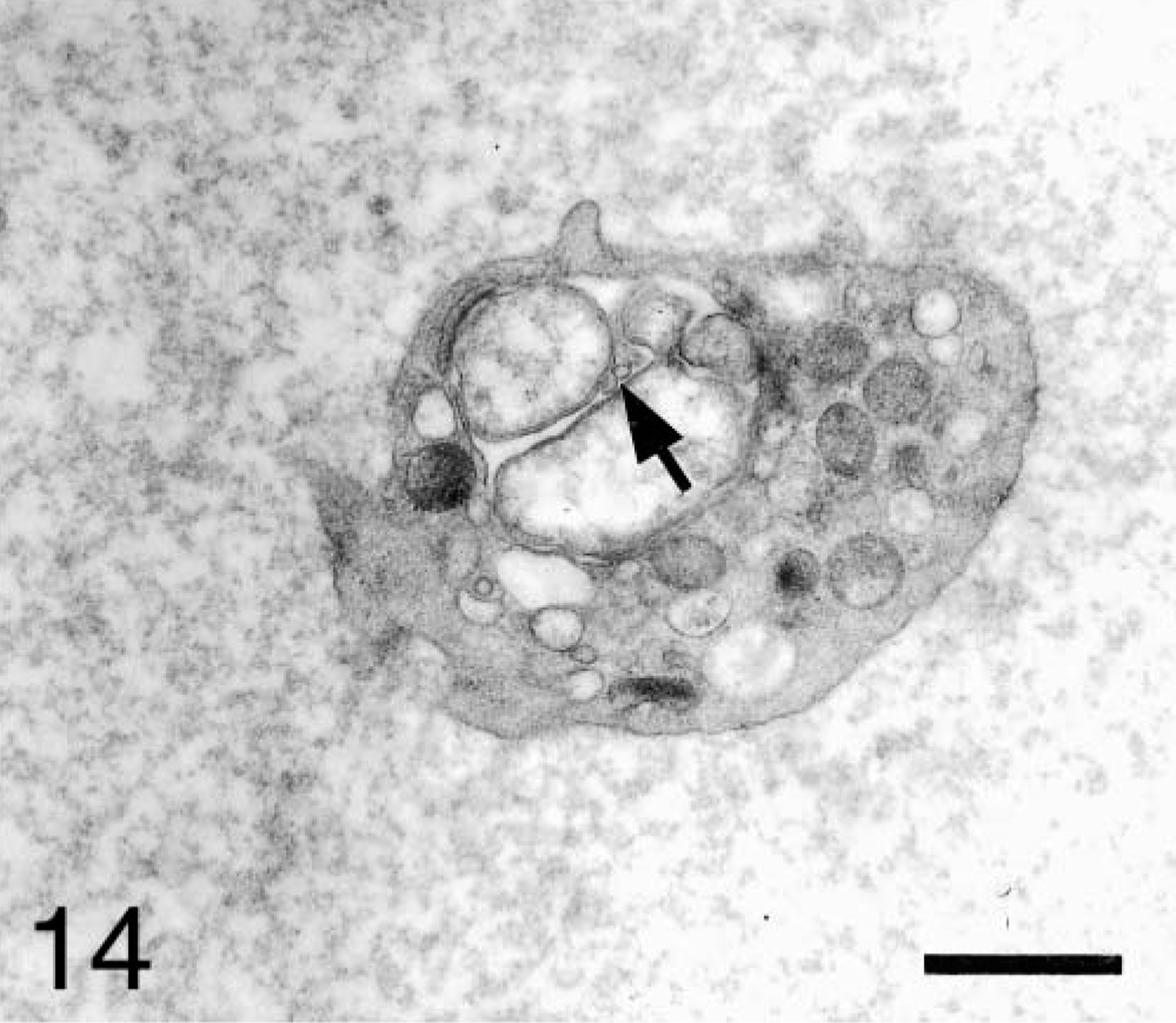
Blood platelet from an experimentally infected dog; dog No. B1, case No. 1. Small electron-dense vesicular structures (arrow) are observed inside the host vacuole. Bar = 1 μm.
Discussion
The microorganisms reported in this study in blood platelets were structurally similar to E. platys strains described by various authors. 14 , 18 , 19 , 24 Most of the organisms were surrounded by inner and outer membranes as described in previous studies of the genus Ehrlichia 11 , 19 , 21 , 24 , 25 , 27 , 28 , 31 and Anaplasma. 22 , 26
The outer membrane was always parallel to the inner membrane, an observation that is in contrast to earlier ultrastructural studies of E. platys 19 , 24 in which a rippled cell wall was reported. Perhaps the sample fixation method used by previous authors caused ripples to form at the level of the cell wall. Rather than a single step of glutaraldehyde fixation, we used a double glutaraldehyde fixation method to obtain good fixation of all platelet structures. The thickened membrane we observed in some organisms is similar to the membrane seen in rickettsial bodies in human platelets. 3
In our study, we observed 1–15 microorganisms per host vacuole, 8 is the maximum number of organisms previously reported in E. platys. 19 Nevertheless, Sells et al. 31 reported 1–33 thin-section profiles of E. equi in a study of horses experimentally inoculated with E. equi–infected blood, and Popov et al. 25 reported 30–60 profiles in an ultrathin section of E. canis in the cytoplasm of host macrophages. Ribeiro et al. 26 reported two to eight organisms by inclusion in Anaplasma marginale. Frequently, we observed that anaplasmal organisms lined the periphery of the host vacuoles, as observed for E. equi. 31
The electron-lucent central zone and the electron-dense granular substance in the periphery of organisms were similar to those reported by Buoro et al. 9 in ehrlichial organisms within feline mononuclear cells. The granular substance may represent clusters of ribosomes as described for E. equi and E. canis. 11 , 27 , 28 , 31 Fine fibrils near the central region of the organisms have been previously reported in ehrlichial species to be deoxyribonucleic acid (DNA). 11 , 19 , 21 , 27 , 28 , 31
Empty vesicles such as those observed outside the vacuole have been implicated in the transfer of host cellular material that is associated with the destruction of the constituents of the vacuole. 31 It is important to note that in our study these empty vesicles were observed in platelets with the highest number of organisms.
Tubular and vesicular structures observed inside the vacuoles containing an electron-dense fibrillar matrix and a thickened membrane have not been reported in previous studies on E. platys. However, similar tubules were described in E. canis by Popov et al., 25 and small organisms were observed in granulocytes after E. equi infection. 31
Mitochondria were in contact with the vacuole membrane and sometimes established connections with anaplasmal organisms, as also has been seen with E. canis and E. chaffeensis. 25 In the present study, platelet lysosomes were also located around the vacuole but were never observed in close contact with the vacuole membrane. Possibly A. platys infection of platelets impedes the fusion of the vacuole membrane with the lysosomes.
The distinct ultrastructural differences observed among various organisms inside the same platelet could correspond to different developmental stages of Anaplasma. Anderson et al. 1 have described the ultrastructural characteristics of the various stages in the reproductive cycle of the Ornithosis virus. Some ultrastructural features of A. platys described in this study were similar to those reported in the Ornithosis study.
The initial stage of A. platys development appears to be represented by small bodies with a thickened membrane and a moderately electron-dense protoplasm. During this stage, it seems probable that the inner plasma membrane of the parasite remains in close contact with the outer membrane. The intermediate stage of development appears to be characterized by the condensation of dense material associated with DNA in the central zone of the organism. This is similar to the initial bodies of anaplasmal inclusions within erythrocytes. 22 A pale area containing DNA fibrils surrounds the central area. Electron-dense granules or ribosomes may form the peripheral zone of the anaplasmal organism.
Finally, the mature A. platys bodies are characterized by an electron-lucent central area (with DNA fibrils) and a peripheral electron-dense granular substance (composed of ribosomes). These ultrastructural features are similar to those reported in initial bodies of A. marginale 26 and Paranaplasma caudata. 32 Some of these individual organisms were observed when they were undergoing binary fission. The bean shape is usually associated with the division that transforms a single organism into two organisms. 19 , 26
The organisms described in this study were previously classified as bacteria of the genus Ehrlichia, species platys. This classification of ehrlichial organisms was based on morphology and host cell tropism rather than on genetic phylogeny. 36 Recent studies of these organisms suggest that members of the E. platys group should be reclassified in the genus Anaplasma. 12 The morphologic characteristics of organisms in this study and those of bacterial species of the Anaplasma group, as reported previously by other authors, are similar. However, the morphologic characteristics observed in the initial stage of development of the organisms described in this study have not been reported previously in other anaplasmal organisms. Nevertheless, genetic data indicate a sufficient genotypic and phenotypic relationship between the genera Anaplasma and Ehrlichia 12 to warrant their inclusion in a single genus.
This study is the first detailed description of the ultrastructural development of A. platys. The identification of structural differences between experimentally infected and naturally infected dogs suggests that we may have evaluated two different strains of A. platys. To confirm this observation, a comparative serologic and molecular characterization of the organisms is required.
Footnotes
Acknowledgements
We express our gratitude to Marisel Peña, Jhonny Yanez, and Nerelis Ramos for their useful technical assistance, to Dr. Juan Mirt for permission to use the electron microscope, and to Dr. Edward Breitschwerdt for his advice and critical review of the manuscript. This research was partially supported by CONDES-LUZ.