
Abstract
Twenty-one 3-day-old turkey poults from British United Turkeys of America were orally inoculated with a recently characterized astrovirus, TAstV-2, isolated from turkeys with poult enteritis and mortality syndrome. At 1, 2, 3, 4, 5, 7, and 9 days postinfection (dpi), three inoculated birds were euthanatized, and tissues (intestines, spleen, bursa, and thymus) were collected immediately into 10% neutral buffered formalin. Inoculated birds were diarrheic by 3 dpi, and frothy feces persisted throughout the experimental period. Histologically, there was only slight evidence of enteric damage, which was characterized by mild epithelial necrosis, lamina propria infiltrates, minimal villus atrophy, and mild crypt hyperplasia. In situ hybridization, using a negative sense digoxigenin-labeled riboprobe to the capsid gene of TAstV-2, revealed viral RNA in intestinal epithelial cells at the basal margins of the villi, in distal small intestine, and in cecum at 2 dpi, with subsequent extension to epithelium of the large intestine and proximal small intestine (3–5 dpi). Minimal virus remained by 9 dpi.
Turkey astrovirus (TAstV) was first identified in 1980 in intestinal contents from poults with diarrhea,6 and subsequent experimental studies verified that this virus could cause diarrhea.7–11 Recently, a novel astrovirus was isolated from a group of turkeys with poult enteritis and mortality syndrome.3 This novel astrovirus, TAstV-2, is molecularly distinct from the earlier TAstV, sharing only 35% nucleotide sequence in the capsid gene. To determine the sites and extent of viral replication in turkeys infected with TAstV-2, turkey poults were experimentally inoculated with a defined inoculum containing TAstV-2 and monitored clinically with tissues harvested at various time points postinfection and examined by histopathology and in situ hybridization.
The virus was propagated and purified using specific pathogen-free (SPF) embryos as previously described.3 Intestinal tissue from poults infected with astrovirus was collected, homogenized in phosphate buffered saline (PBS), and clarified by low-speed centrifugation. Any endogenous reovirus was removed from the tissue filtrate by affinity purification using a rabbit polyclonal antibody against reovirus, then the supernatant was passed through a Whatman 0.2-μm filter (Fisher Scientific, Norcross, GA) before being inoculated into the yolk sac of 20-day-old turkey embryos from an SPF Small White flock maintained at the Southeast Poultry Research Laboratory. Following a 5-day incubation period, the embryo intestines were harvested, homogenized, clarified, and filtered in sequence for a total of three passages. Following the final passage, the intestinal fluid was centrifuged for 15 minutes at 2,000 rpm and then filtered through a Whatman 0.2-μm filter. The suspension was then analyzed using a reverse transcription polymerase chain reaction (RT-PCR) technique2 and immunoelectron microscopy to verify the presence of TAstV-2 and the absence of coronavirus and reovirus, as described previously.3
British United Turkeys of America poults from a source with no history of previous TastV-2 exposure were housed in conventional Horsfall isolation units and given unlimited access to water and standard University of Georgia unmedicated turkey starter ration. These poults were negative by RT-PCR for the presence of TAstV-2. Poults from this source have been used in numerous inoculation experiments, and the noninoculated controls have consistently remained free of TAstV-2 by RT-PCR. Twenty-one 3-day-old poults were orally inoculated with 0.2 ml of TAstV-2 inoculum, which consisted of 105 EID50/ml viral particles. Six age-matched noninoculated control poults were maintained in a separate Horsfall unit. Birds were monitored daily for clinical signs of disease. At 1, 2, 3, 4, 5, 7, and 9 days postinoculation (dpi), three inoculated birds were euthanatized by cervical dislocation, and the following tissues were collected in 10% neutral buffered formalin: proventriculus, duodenum, small intestine at site of Meckel's diverticulum, distal small intestine, cecal tonsils, large intestine, spleen, thymus, and bursa. After 24 hours in formalin, tissues were transferred to RNase-free PBS for several hours and then processed for paraffin embedment. Three-micrometer sections were cut and stained routinely with hematoxylin and eosin (HE). Adjacent unstained sections were used for in situ hybridization. Two age-matched noninoculated control birds were euthanatized and tissues collected at 1, 5, and 9 dpi.
A riboprobe was generated by digesting plasmid p25.5, which contains a 1.5-kilobase (kb) segment of the extreme 3′ end of the TAstV-2 genome, with BamHI.3 Subsequent transcription with T7 RNA polymerase and digoxigenin-labeled UTP (Roche Molecular, Indianapolis, IN) created an antisense riboprobe approximately 1.6 kb in length. Digoxigenin incorporation was verified by dot-blot.
Riboprobe in situ hybridization was performed according to previously described techniques.1 Tissue sections were deparaffinized with Hemo-De (Fisher Scientific) and digested with 35 µg/ml Proteinase K for 15 minutes at 37 C. Hybridization occurred overnight at 42 C, using approximately 35 ng of digoxigenin-labeled riboprobe per slide in 5× standard sodium citrate (SSC), 50% formamide, 5% modified milk protein (Roche Molecular), 1% N-lauroylsarcosine, and 0.02% sodium dodecyl sulfate (SDS). The following day, slides were washed in increasingly stringent solutions: 2× SSC with 1% SDS for 30 minutes at 50 C, 1× SSC with 0.1% SDS for 30 minutes at 50 C, 1× SSC for 15 minutes three times at room temperature, and 0.1× SSC for 15 minutes at room temperature. After the posthybridization washes, sections were incubated with anti-digoxigenin antibody conjugated to alkaline phosphatase (Roche Molecular) for 2 hours at 37 C. The chromogen was nitroblue tetrazolium and bromcresylindolyl phosphate. Development progressed for 1–3 hours. Sections were stained lightly with hematoxylin and coverslipped with Permount for a permanent record. Each group of slides was processed with a positive control section consisting of a section of positive embryo intestine and with negative control sections from noninoculated poults. Additional negative controls, i.e., omission of probe or use of a nonsense probe, were not processed because previous studies have indicated that results are consistently negative (probe omission) or spurious (nonsense probe).1
Inoculated birds were listless at 1 dpi and developed diarrhea at 3 dpi. Feces were fluid, frothy, and yellow. The diarrhea persisted to the end of the experiment. Birds were depressed through 9 dpi. No clinical signs of disease were observed in noninoculated control birds.
There were no abnormal morphologic findings at 1 or 2 dpi. At 3 dpi, intestines and ceca were dilated and thin walled, intestinal contents were abundant, watery, yellow, and frothy, and gallbladders were enlarged. At 5, 7, and 9 dpi, thymuses and bursae were smaller than those of noninoculated controls.
All histopathologic changes were relatively mild. At 2 dpi, there was increased cytoplasmic eosinophilia in infrequent clusters of epithelial cells in the small intestine, primarily at the basal part of the villi. At 3 and 4 dpi, there was degeneration and necrosis of individual cells within the epithelium of the small intestine (Fig. 1). These single cells were darkly stained and markedly narrowed, as if collapsed. At 5 dpi, infrequent necrosis affecting clusters of cells near the surface of the distal small intestine was noted. There was erosion, with adjacent enterocytes stretching to cover (Fig. 2). Some shortening of villi was noted at 5 dpi and persisted through 9 dpi. Intestinal crypt hyperplasia was mild, beginning at 4 dpi and progressing through 9 dpi. Lamina propria of the small intestine had increased numbers of cells beginning at 3 dpi, predominantly lymphocytes but a few macrophages. This infiltrate persisted, but by 9 dpi there also were notable clusters of heterophils within the lamina propria.
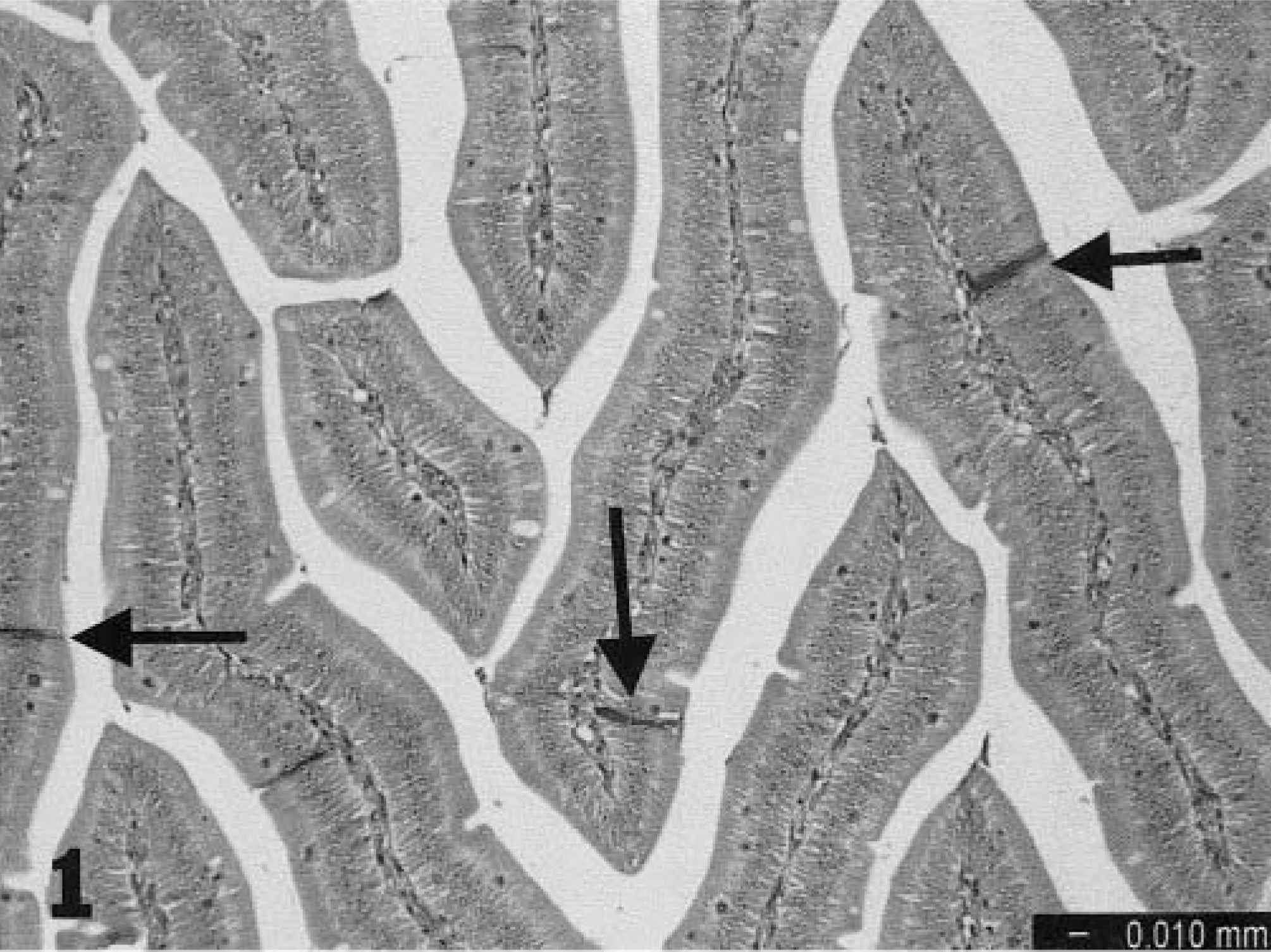
Epithelial cells, small intestine; turkey poult. Individual cell necrosis (arrows) is evident 4 days after oral inoculation with TAstV-2; HE.

Distal small intestine; turkey poult. Foci of degenerating epithelial cells (arrows) are visible 5 days after oral inoculation with TAstV-2, HE.
Only three nonintestinal tissues were examined: spleen, thymus, and bursa. At 2 dpi, there was increased prominence of splenic Schweigger-Seidel sheaths. Development of selected splenic lymphoid follicles began at 3 dpi, and germinal centers were visible at 5, 7, and 9 dpi. Changes in bursae consisted of some follicular edema at 1 dpi and nuclear fragmentation of lymphocytes beginning at 2 dpi and progressing in severity through 9 dpi. Thymic changes were minimal, and this organ was not noticeably different from that of controls.
The in situ hybridization data are summarized in Table 1. Specificity was confirmed by consistent staining of positive control embryo intestines and total lack of staining in tissues of noninoculated control birds. All positive hybridization signals were restricted to epithelial cells of the gut in inoculated birds. At 1 and 2 dpi, scattered positive intestinal epithelial cells were seen in the distal small intestine and cecal tonsils (Fig. 3). In general, positive cells were found at the deep edges of the villi and rarely in crypts. At 3–5 dpi, scattered positivity was seen in the same areas but was more likely to be present as clusters of positive cells (Fig. 4). Staining extended to include single cells or groups of cells in the colon, small intestine at Meckel's diverticulum, duodenum, and large intestine. At 7 dpi, staining was less extensive except for some areas of large intestine. By 9 dpi, staining was relatively infrequent. No staining was seen in any of the lymphoid tissues examined.
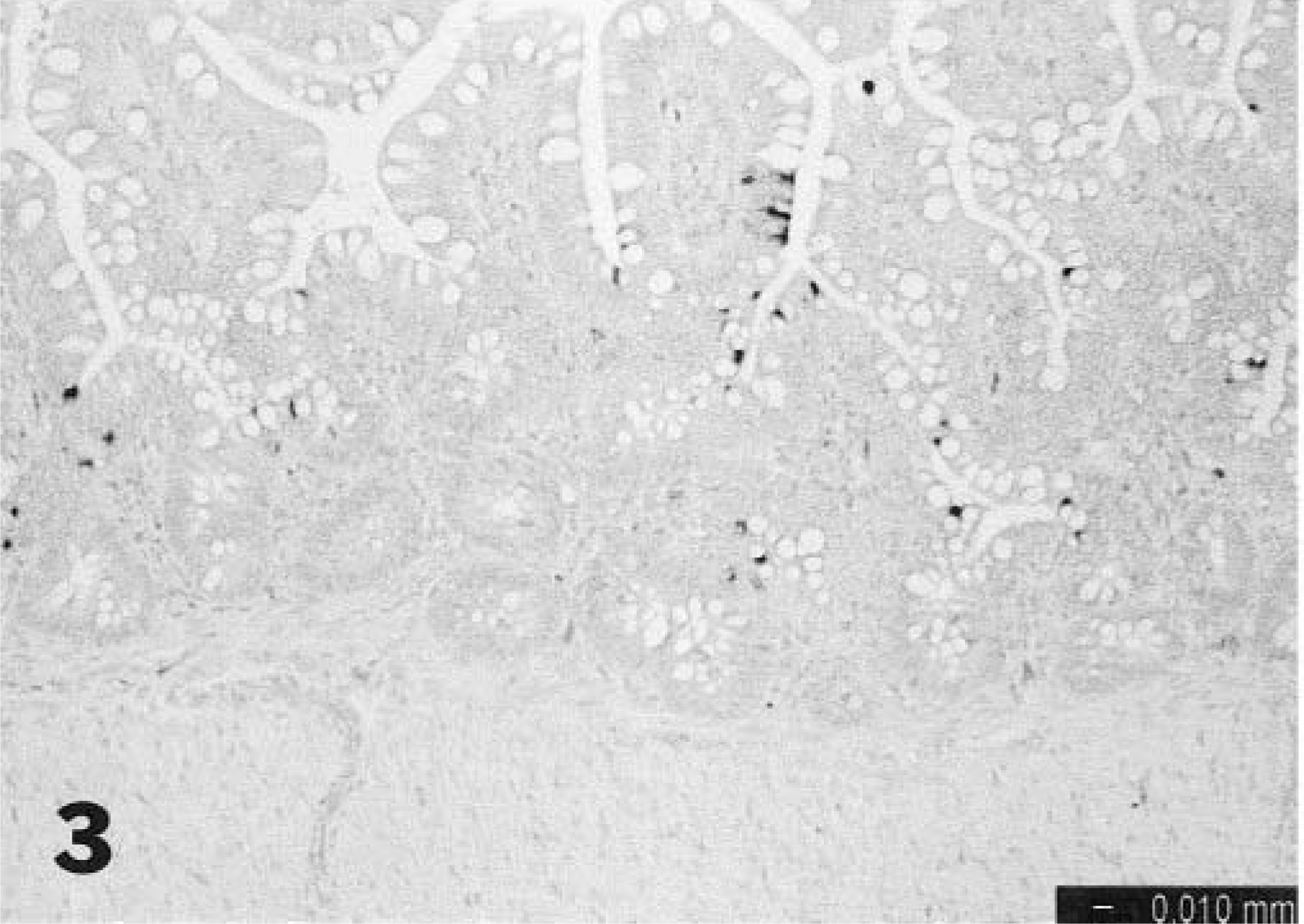
Distal small intestine; turkey poult. Scattered positive intestinal epithelial cells are visible 1 day after oral inoculation with TAstV-2, In situ hybridization.
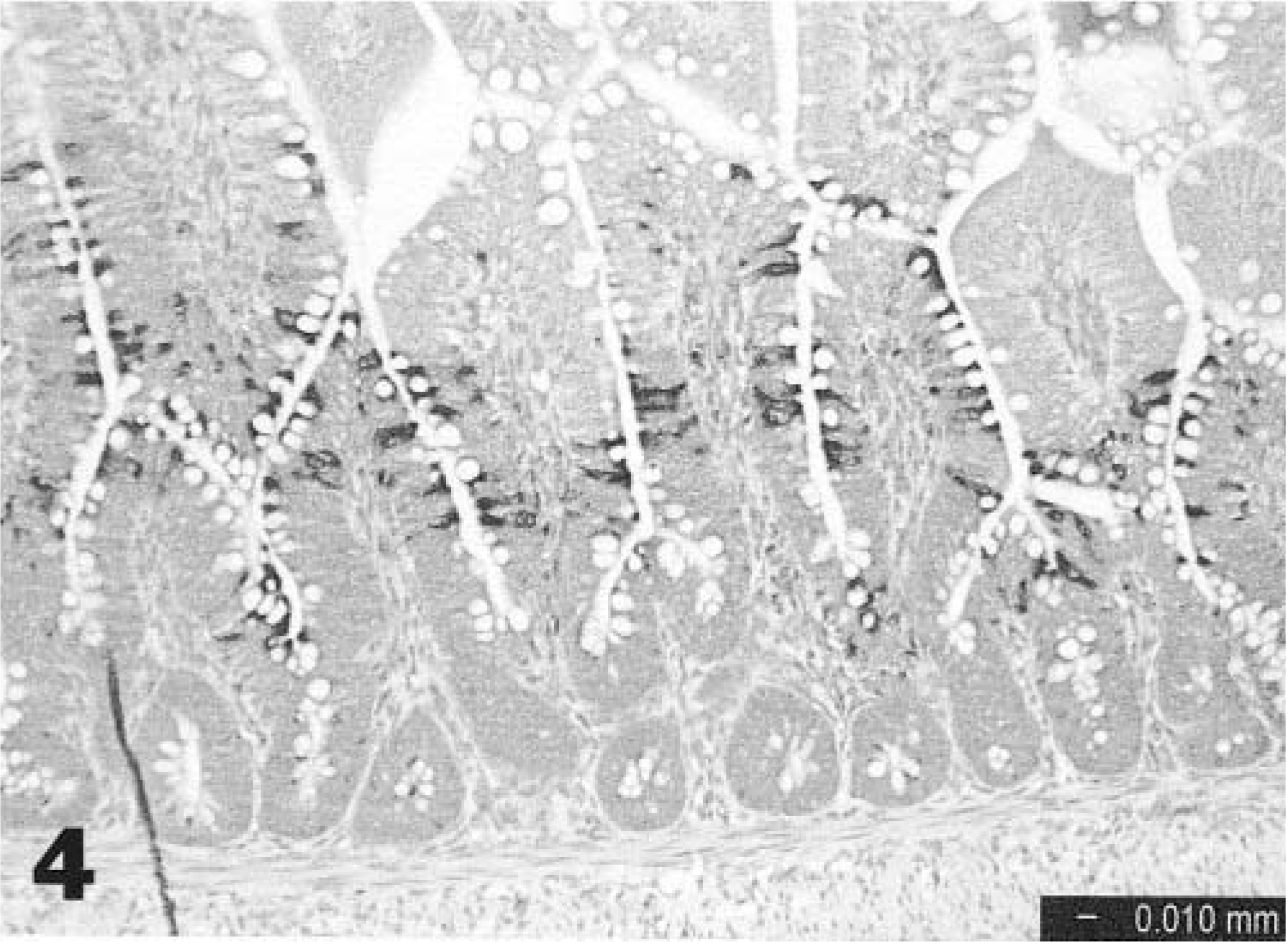
Distal small intestine; turkey poult. Extensive positive staining is visible in intestinal epithelial cells 3 days after oral inoculation with TAstV-2, In situ hybridization.
Replication of TAstV-2 in turkey poults as detected by in situ hybridization.∗

− = no staining; + = <5 positive cells/high-power field; ++ = 5–15 positive cells/high-power field; +++ = >15 positive cells/high-power field (all based on an average of 10 randomly selected high-power fields).
Studies with turkey astrovirus isolated in the early 1980s involved both commercial and SPF birds. The SPF poults were inoculated with intestinal homogenate that had been filtered, sonicated, and shown by immunoelectron microscopy to contain astrovirus, and these birds developed dilated ceca with yellowish frothy contents and thin intestinal walls.7 Subsequently, commercial poults were inoculated with a similar preparation. These commercial poults developed diarrhea, and astrovirus was detectable by electron microscopy in enterocytes on the sides and base of villi in small intestine.11 Measurement of intestinal disaccharidase activity in these infected poults indicated that malabsorption was playing a role in causing the diarrhea.12
Members of the astrovirus family have been implicated in enteric disease in some mammalian species, including humans.4,5 Experimental infection of lambs and calves with their specific astroviruses demonstrated that these viruses also have a tropism for small intestinal epithelium. In lambs, the mature apical villus cells were specifically targeted,10 whereas in calves astrovirus was found predominantly in dome epithelium of the ileum.13
Oral inoculation of commercial turkey poults with TAstV-2 resulted in clinical depression and mild to moderate diarrhea. Clinical signs of enteritis were evident as early as 3 dpi. At postmortem examination, the main findings were dilated intestines filled with frothy yellow feces and mild atrophy of thymus and bursa. Histologically, there was only slight evidence of enteric damage, characterized by mild epithelial necrosis, lamina propria infiltrates, minimal villous atrophy, and mild crypt hyperplasia. In situ hybridization revealed the presence of viral RNA in intestinal epithelial cells. Visualization of viral RNA was confined to cells along the base and sides of villi, predominantly in distal small intestine and cecum, but with extension both proximally and distally. The mechanism by which TAstV-2 infection results in diarrhea remains uncertain. The presence of intestinal epithelial degeneration and necrosis points to malabsorption as one mechanism. There was no viral RNA detected by in situ hybridization in extraintestinal tissues.