
Abstract
Between August and September 2000, five 2–7-year-old cows in Korea exhibited neurologic signs and were diagnosed as infected with Akabane virus based on the results of histopathology, immunohistochemistry, serology, and reverse transcription polymerase chain reaction (RT-PCR) analysis. Immunohistochemistry and RT-PCR were equally effective and sensitive for diagnosing Akabane virus infection during the early stage of infection. Typical lymphohistiocytic inflammation characterized by perivascular mononuclear cell infiltration, gliosis, neuronophagia, and neuronal loss was noted in the brain and the ventral horn gray matter of the spinal cord. The lesions in the brain were most prominent in the pons and medulla oblongata. Akabane virus antigen was detected in the brain and spinal cord, mainly in degenerating neurons and glial cells. RTPCR analysis revealed a target band of expected size in four cows. This is the first report on an outbreak of natural Akabane virus infection in adult cattle.
Akabane disease (AD) is an arthropod-borne bunyaviral disease associated with abortion, stillbirth, and various congenital abnormalities in cattle, goats, and sheep. Akabane virus (AV) is a member of the Simbu group, genus Bunyavirus, family Bunyaviridae, and was first isolated from Aedes vexans and Culex triaeniorhynchus in 1960 and 1964, respectively, in Japan.4,11 Sporadic outbreaks of AD have been reported in Japan, Taiwan, Australia, and Israel.1,8,18 In Korea, the first AD outbreak was reported in 1980.2 The pathogenesis of AV infection in cattle is well understood. Pregnant cows infected by AV through vectors generally do not develop any apparent clinical signs. However, during the early and mid to late gestation periods infection results in abortion and congenital arthrogryposis-hydranencephaly syndrome (AHS), respectively.4,10
Although nonsuppurative encephalomyelitis may be observed in the late gestation infection in calves without AHS, encephalomyelitis induced by AV is unusual in cattle after birth. A few case of this condition caused by a highly virulent AV strain known as Iriki strain were reported in Japan and Taiwan, suggesting that AV may cause nonsuppurative inflammation in the brain and spinal cord of cows through horizontal rather than vertical transmission.12,15 In July 1998, a severe outbreak of congenital abnormalities in calves was reported in most areas of Japan; a 5-month-old bull calf and 27 calves ranging from 1 to 27 days of age were examined, and their cases were diagnosed as AV infection.19 Here, we describe an outbreak of natural AV infection in adult cows as revealed through histopathology, immunohistochemistry, neutralizing antibody assay, and reverse transcription polymerase chain reaction (RT-PCR) analysis.
Between August and September 2000, 5 adult cows in Wonju City ranging from 2 to 7 years of age developed neurologic signs, including hypersensitivity, tremor, locomotor ataxia, or lameness. The cows were from five different farms where the morbidity was about 30%. Abortions and congenital abnormalities were not detected in these farms. Affected cows generally had normal body temperature and appetite but were unresponsive to fluid, nutritional, and antibiotic therapies. Blood and serum chemistry panel results were within the normal range. Because of a poor prognosis, the cows were euthanatized and subjected to postmortem examinations to determine the etiology of this problem (Table 1).
Summary of histopathology, immunohistochemistry, and antibody titers for Akabane virus in cows.

+++ = severe; ++ = moderate; + = mild.
Akabane viral detection: + = detected; - = not found.
ND = not done.
Brain, spinal cord, and skeletal muscles collected from two of the slaughtered cows were submitted to the Department of Veterinary Pathology, Seoul National University, for histologic diagnosis. No significant gross abnormalities were found at slaughter. The submitted specimens were fixed in 10% phosphate-buffered neutral formalin, routinely processed, embedded in paraffin, and stained with hematoxylin and eosin (HE) for histopathologic examination. Brain sections were taken from the cerebral cortex, midbrain, cerebellum, pons, and medulla oblongata. Two sections were taken from cervical, thoracic, and lumbar spinal cord. The lesion severity was graded as mild, moderate, or severe based on the extent of the lesion.
Immunohistochemical identification of AV was performed on replicate paraffin-embedded sections of the brain and spinal cord as previously described.7 Sections were mounted on Probe-On slides (Fisher Scientific, Pittsburgh, PA), and unlabeled rabbit polyclonal antibody directed against the OBE-1 strain of AV was used as the primary antibody.7 The standard avidin–biotin–peroxidase complex (ABC) method was used according to the manufacturer's protocol (Zymed, South San Francisco, CA) to demonstrate the antigen; 3,3-diaminobenzidine (DAB) was used as the chromogen. Control procedures included omission of the primary antibody and substitution of an isotype-matched irrelevant antibody. Brain tissue of a mouse experimentally infected with the OBE-1 strain of AV was used as a positive control.7
Pooled brain and spinal cord specimens from each cow were ground with liquid nitrogen, and total RNA was isolated using TRIZOL (Gibco BRL, Grand Island, NY). Five micrograms of total RNA was reverse transcribed into the first strand cDNA at 37 C for 60 minutes using a cDNA synthesis kit (Takara, Shiga, Japan) and heated at 95 C for 5 minutes to terminate the reverse transcription reaction. The AV S gene was amplified from 3 µl of cDNA mixture in a final volume of 25 µl PCR mixture containing 10× PCR buffer (10 mM Tris-HCl, 50 mM KCl, 0.1% Triton X-100), 3.5 mM of MgCl2, 0.3 mM each of dNTP, 50 pmol each of forward primer F2 (5′-ACCAGAAGAAGGCCAAGATG-3′) and reverse primer R2 (5′-CACACGGTGCATGTCGATAA-3′), and 1 unit of Taq DNA polymerase. Primer sets for the PCR amplification of the AV S gene were selected based on published sequence.5 The PCRs were carried out using a Thermocycler 9600 (Perkin-Elmer, Norwalk, CT). Reaction mixtures were first denatured at 94 C for 5 minutes, and amplification was performed for 30 cycles at 94 C for 30 seconds, at 58 C for 30 seconds, and at 72 C for 30 seconds, followed by an extension at 72 C for 7 minutes. The brain from a noninfected cow was used as a negative control. The OBE-1 strain of AV was used as a positive control. The amplified PCR products were run on a 1.2% agarose gel. PCR for bovine ephemeral fever virus (BEFV), bovine immunodeficiency virus (BIV), infectious bovine rhinotracheitis virus (IBRV), bovine viral diarrbea virus (BVDV), bovine herpesvirus type 5 (BHV-5), and Chlamydia were also performed. Primer sequences and PCR conditions for each virus were selected based on published literature.6,13,14,16,17 DNA extracted from paraffin blocks of the brain and spinal cord were also amplified for the detection of Neospora caninum as previously described.3
Serum was collected from four adult cows at necropsy. After centrifugation at 2,000 rpm, the serum was stored at −20 C until use. The serum neutralization (SN) test was performed for the AV (OBE-1) and the Aino virus (JaNAr28 strain). The neutralization antibody titers represented the highest dilution rate of examined sera that inhibited a cytopathic effect for the cultured cells.19 The SN test for BEFV, Ibaraki disease virus, BVDV, and Japanese encephalitis virus, an indirect fluorescent antibody test for Chlamydia psittaci and N. caninum, and an enzyme-linked immunosorbent assay for Aujeszky's disease were also performed using standard methods.9,13 Direct fluorescent antibody (FA) tests for IBRV, BVDV, and rabies virus were conducted with specific conjugates on frozen sections of brain and spinal cord.13 Portions of brain and spinal cord from all five cows were collected aseptically, and routine aerobic and anaerobic bacterial cultures were performed.
Although the severity and distribution of lesions were different among the five cows, all examined cows had typical lymphohistiocytic inflammation in the brain and spinal cord (Table 1). The lesion in the brain was mainly focused in the pons and medulla oblongata, with moderate to severe perivascular infiltration of primarily lymphocytes and histiocytes with fewer plasma cells, multifocal gliosis, neuronal necrosis, and occasional neuronophagia with microglial cells and neutrophils (Figs. 1, 2). Similar mild to minimal changes were observed in the midbrain but rarely in the cerebral and cerebellar cortices. Mild perivascular lymphohistiocytic cuffing was noted in scattered submeningeal areas.
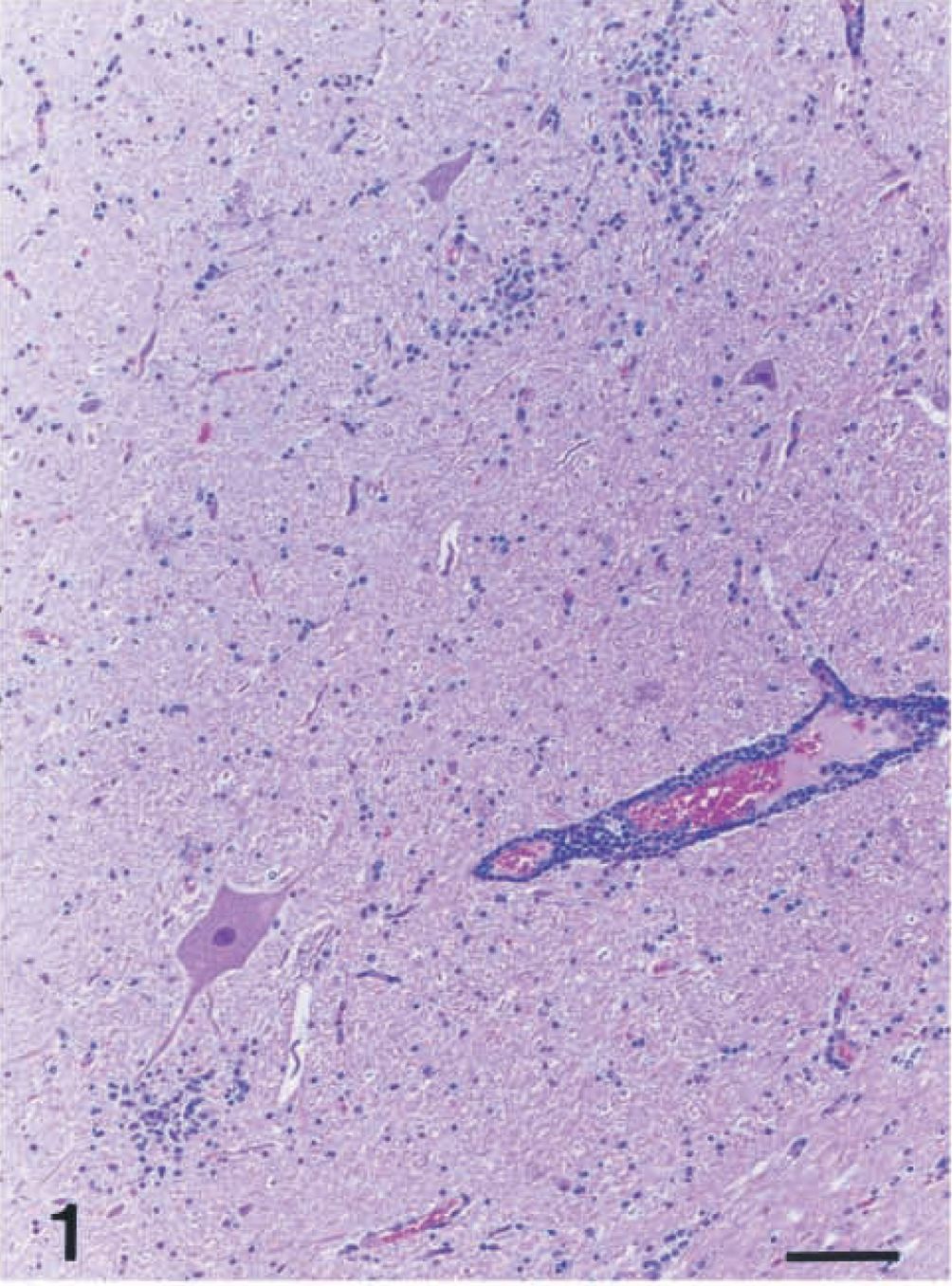
Medulla oblongata; cow No. 2. Note perivascular cuffing by mononuclear cells and multifocal gliosis. HE. Bar = 80 μm.
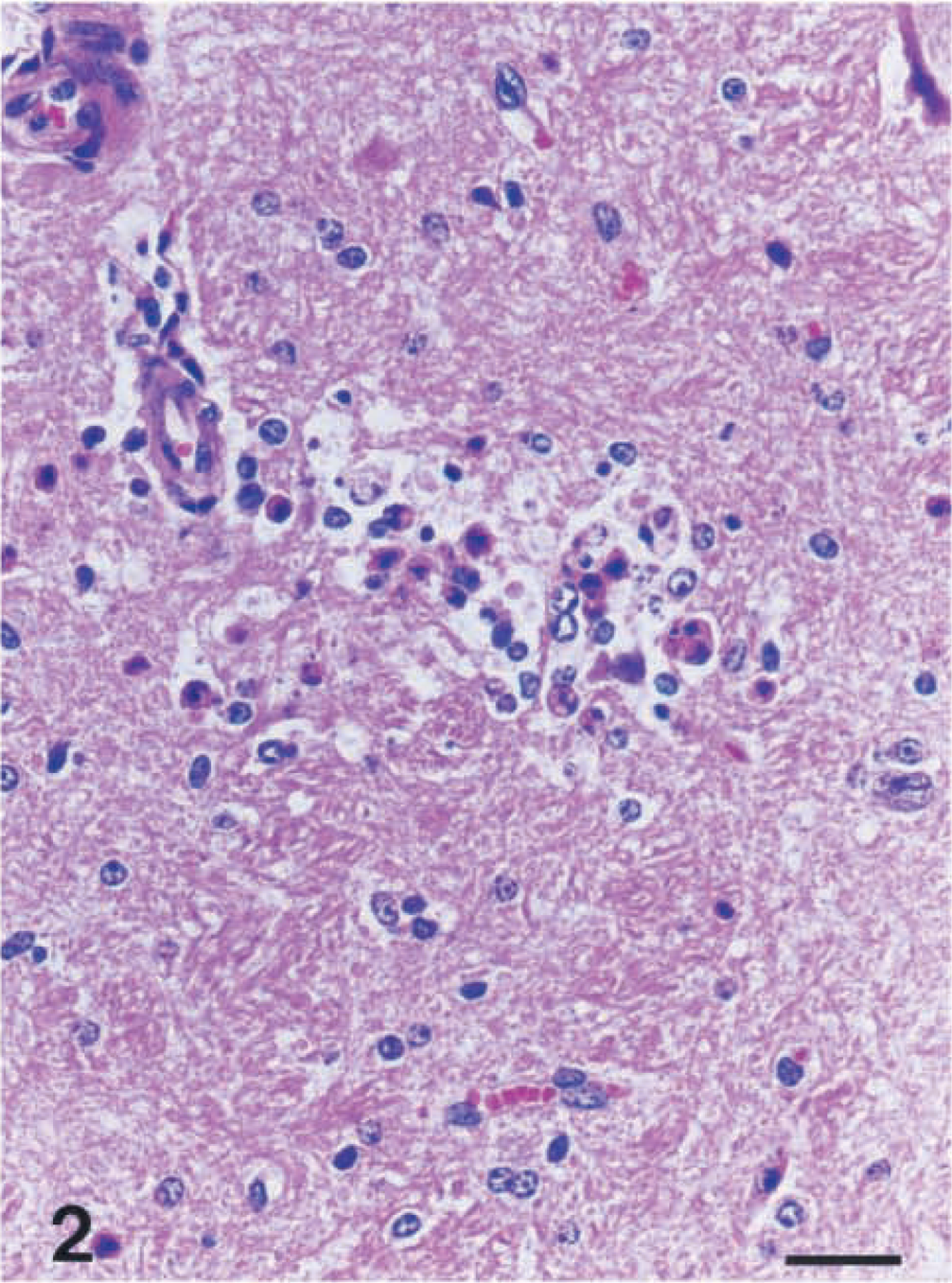
Medulla oblongata; cow No. 3. Note neuronophagia. HE. Bar = 55 μm.
In both the left and right ventral horn gray matter of the spinal cord, lymphohistiocytic perivascular cuffing, gliosis, and neuronal necrosis and loss were observed (Fig. 3). Mild perivascular cuffing and occasional degenerating swollen axons were noted in the spinal cord white matter. However, the inflammatory changes were more severe in the spinal cord than in the brain, and no difference in lesion severity was observed among the spinal cord regions. Histologically, skeletal muscles of the two examined cows were severely atrophic, and mucinous change in the stroma was noted.
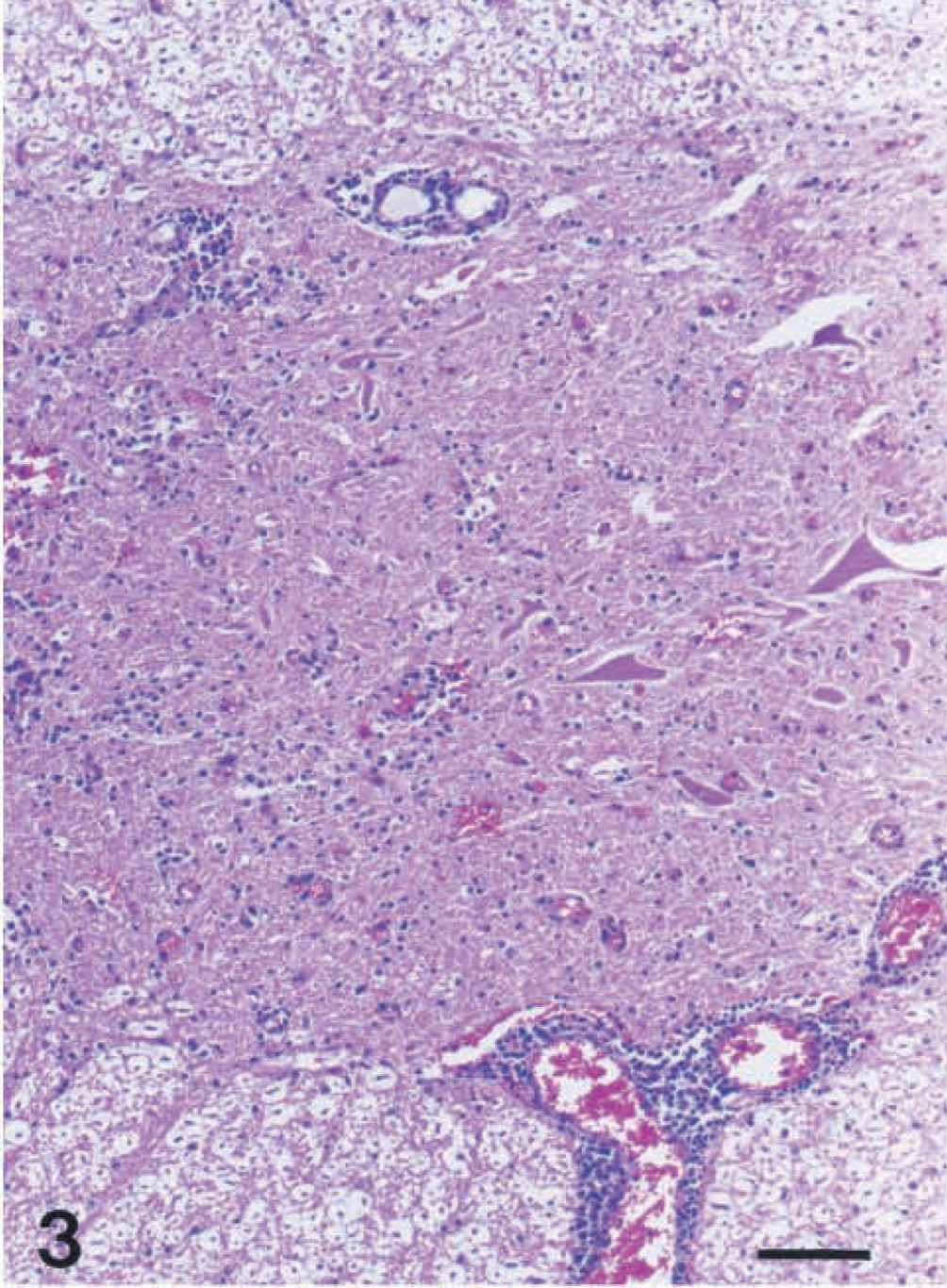
Spinal cord; cow No. 1. Gliosis and perivascular cuffing were noted in the ventral horn gray matter. HE. Bar = 80 μm.
Immunostaining revealed AV antigen in the brain and spinal cord in 2/5 and 4/5 cows, respectively (Table 1). Both in the brain and the spinal cord, the presence of viral antigen was generally colocalized with the inflammatory lesion, although the staining density or the number of positive cells was not correlated with lesion severity. Viral antigens were mainly present in the cytoplasm of the degenerating neurons and glial cells (Fig. 4). Muscle tissue samples from two cows tested were negative for AV antigen by immunohistochemistry. Cow No. 4, which was negative for AV antigen by immunohistochemistry, tested positive for AV by RT-PCR analysis.
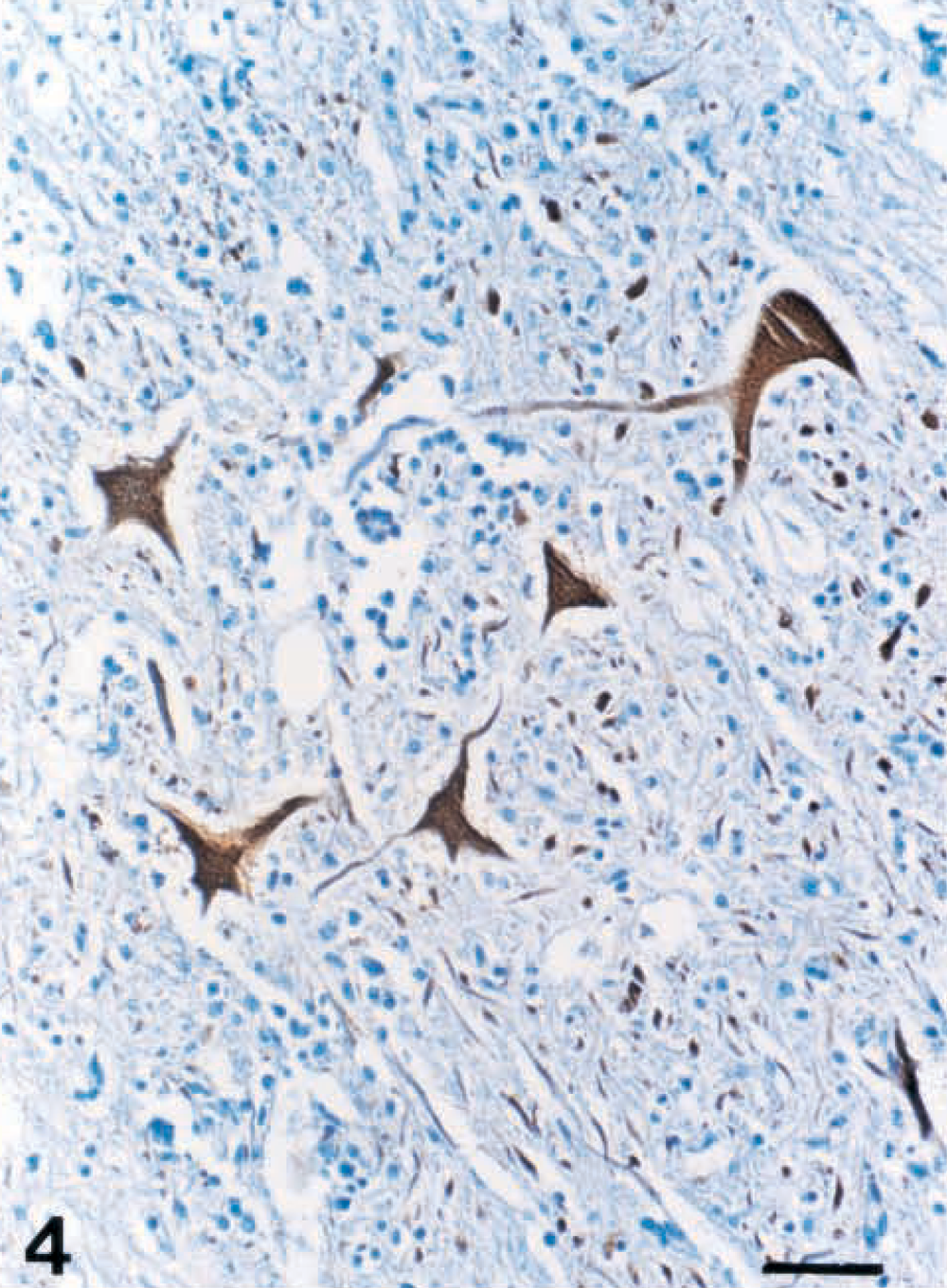
Spinal cord; cow No. 1. Akabane viral antigens were present in the degenerating neurons and glial cells. ABC immunohistochemistry, DAB with Mayer's hematoxylin counterstain. Bar = 80 μm.
According to results of the SN test for AV, 4 cows had antibody titers ranging from 1:64 to 1:256. In all cows, antibody titers to Aino virus were <1:4. All cows tested showed no significant antibody titers to BEFV, Ibaraki disease virus, BVDV, and Japanese encephalitis virus with the SN test. Antibody titers to Chlamydia spp., N. caninum, and Aujesky's disease virus were also negative.
In RT-PCR analyses of the brain and spinal cord samples using primers specific for the AV S gene, PCR products of approximately 347 base pairs were detected in Cow Nos. 1–4 and in the positive control (Fig. 5). No PCR product was produced when cDNA from Cow No. 5 and noninfected negative control were used for the template (Fig. 5). However, all cows tested were negative by PCR for BEFV, BIV, BVDV, IBRV, BHV-5, Chlamydia spp., and N. caninum.
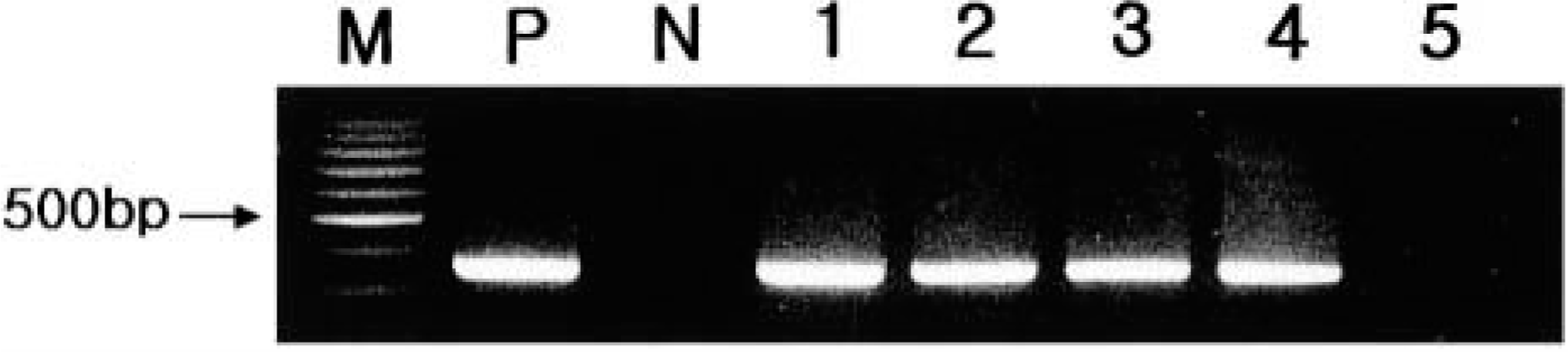
Detection of Akabane virus S gene by the RT-PCR method using the Akabane virus consensus primers. Lanes 1–4 show amplification of a 347-base pair (bp) Akabane virus DNA fragment. M = 100-bp scale molecular weight marker; P = positive control (OBE-1); N = negative control; lane 1 = cow No. 1; lane 2 = cow No. 2; lane 3 = cow No. 3; lane 4 = cow No. 4; lane 5 = cow No. 5.
The FA tests on frozen tissues from all five cows were negative for IBRV, BVDV, Chlamydia spp., and rabies virus antigens. Bacterial cultures failed to isolate any pathogen responsible for the brain and spinal cord lesions.
Based on the results of histopathology, immunohistochemistry, RT-PCR, and serology, the five cows were determined to be infected by AV. Other possible causes of encephalitis in cattle were all ruled out using appropriate antigen and antibody detection methods. The pattern and distribution of histologic lesions observed in the five cows were very similar to those in previous reports on AV infection.19 AV has a tropism for motor neurons in the spinal cord, as detected through immunohistochemistry, and causes neurogenic skeletal muscle atrophy or dysplasia that is believed to be the cause of the lameness and locomotor problem exhibited by these cows.4 Severe muscular atrophies were observed in two cows examined.
AV is transmitted by the biting midge Culicoides brevitarisis in Korea, and disease occurs primarily during the summer rainy season, when the vector becomes active.5 Although this disease outbreak subsided by the end of October 2000, there was still a marked increase in the incidences of bovine abortion and AHS due to AV infection in fall 2000 and winter 2001, indicating the hematogenous spread of virus from the infected dam to the fetuses.
Virus isolation is not an easy procedure for routine diagnostic work and is not always successful in detecting AV infection. The present study revealed that both immunohistochemical and RT-PCR methods were useful for confirming diagnosis of AV infection in adult cows. However, when infection progresses to a chronic phase, immunohistochemistry was not sensitive enough for detecting small amounts of antigen.19 Therefore, when adult cows showing neurologic signs are detected in the countries where AD is endemic, RT-PCR analysis of cerebrospinal fluid or buffy coat for monitoring possible involvement of AV is highly recommended.
Recently, AD was reported in a 5-month-old bull calf and in 1–27-day-old calves in Japan, suggesting that AV is a potential cause of encephalitis in young cattle outside the neonatal period.19 Our study demonstrated that adult cows could also develop brain and spinal cord lesions associated with AV infection. Therefore, AV should always be considered as a possible etiology for lymphohistiocytic encephalomyelitis in adult cattle, particularly in countries where the vectors are present.
A highly virulent Iriki strain of AV has the potential to cause encephalitis in cattle through natural or experimental postnatal infection.12,15 Isolation and characterization of the virus, including its pathogenecity in the infected cows, is in progress. Because AD outbreaks can recur, carefully controlled vaccination against AD is highly recommended once the diagnosis is confirmed.
Footnotes
Acknowledgements
This study was supported by a research grant from the Ministry of Agriculture and Forestry (399002-3) and the Brain Korea 21 Project. We extend our appreciation to Dr. Keun-Ha Park for submission of the cases.