
Abstract
Severe cutaneous hemorrhages with dermal and subcutaneous capillary angioplasia were seen in aborted and stillborn piglets, concurrently with an acute outbreak of porcine reproductive and respiratory syndrome virus (PRRSV) abortions. Histologically, the lesions consisted of angioblastic endothelial cells and immature capillary vascular structures coursing through the edematous myxomatous dermis and subcutis. Proliferating capillaries often were surrounded by large and foamy macrophages that stained positively for PRRSV by immunohistochemistry. The sudden appearance of these vascular lesions during the PRRSV outbreak and their abrupt disappearance after the abortion storm, along with the immunohistochemical localization of PRRSV-positive macrophages adjacent to the proliferating capillaries, suggest that PRRSV likely played a role in the development of these unusual lesions.
Proliferative lesions of the cutaneous microvasculature are rare in neonatal domestic animals. When they occur, they generally represent localized hyperplasia, dysplasia, or hamartomas. 9,15 This case report describes unusual widespread proliferation of capillary vascular structures and severe cutaneous hemorrhages in the skin of full-term stillborn and live-born piglets during an acute outbreak of porcine reproductive and respiratory syndrome virus (PRRSV).
PRRSV is a positive-strand RNA Arterivirus that causes a variety of clinical manifestations in swine, including acute febrile illness, abortions and stillbirths in adult sows, and histiocytic interstitial pneumonia in neonatal and postweaned pigs. 2,11 Transmission of PRRSV can occur through vertical spread from sow to fetal pigs and horizontally by aerosol, semen, saliva, or body fluids. Porcine dermatitis and nephropathy syndrome is a newly described systemic and dermal necrotizing vasculitis and fibrinous glomerulitis that has been associated with PRRSV and porcine circovirus type 2 (PCV-2) infection. 10 Recently, particularly virulent strains of PRRSV have caused severe abortion storms accompanied by high levels of adult mortality. PRRSV strains that are isolated from these severe outbreaks typically exhibit a 1-4-2 pattern by restriction fragment length polymorphism analysis of ORF5. 7 For a more complete description of lesions and epidemiology of PRRS virus the reader should consult one of the many review articles. 2,11 In the present report, we describe the gross and microscopic lesions (Figs. 1–5) of cutaneous hemorrhage and proliferative vasculopathy in porcine neonates infected with PRRSV.
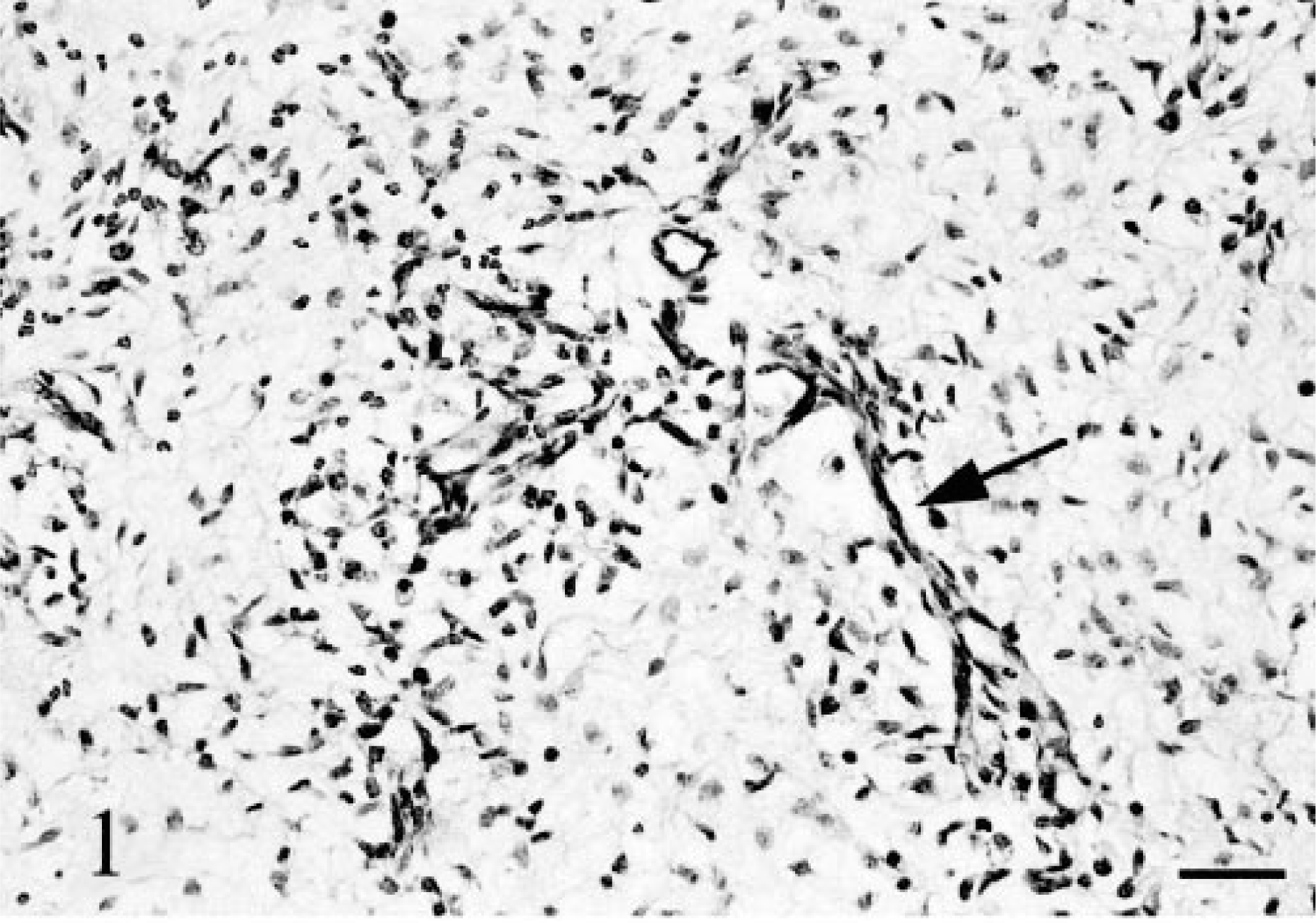
Dermis. Numerous anastomotic, immature capillaries are enmeshed in a loosely organized myxomatous dermal matrix. Arrow indicates one of many branching capillaries. HE stain. Bar = 75 µm.
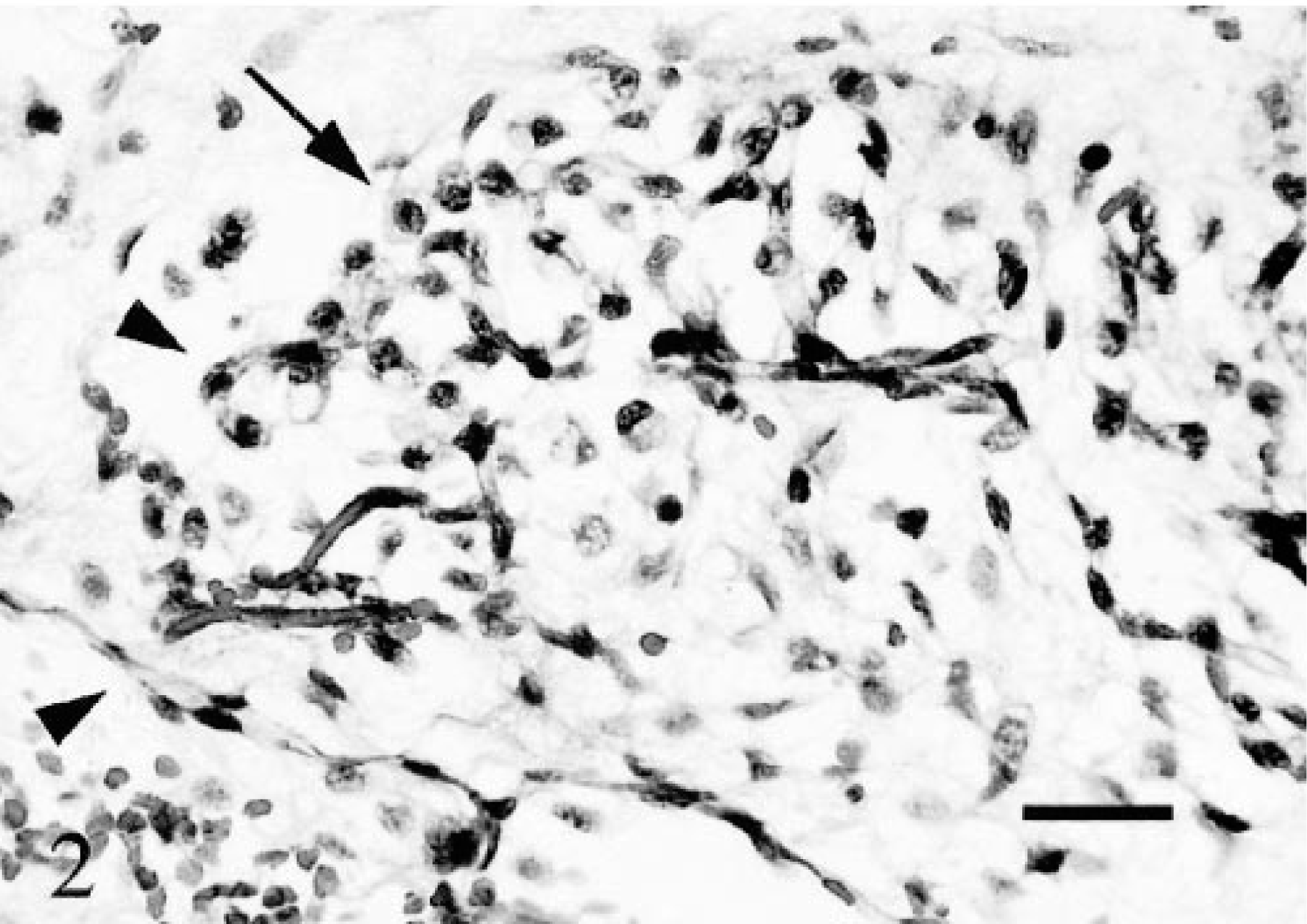
Dermis. Immature capillaries are encircled by loosely organized mesenchymal matrix containing large foamy macrophages (arrow). Note vascular structures, some of which lack clearly discernable lumina (arrowheads). HE stain. Bar = 25 µm.
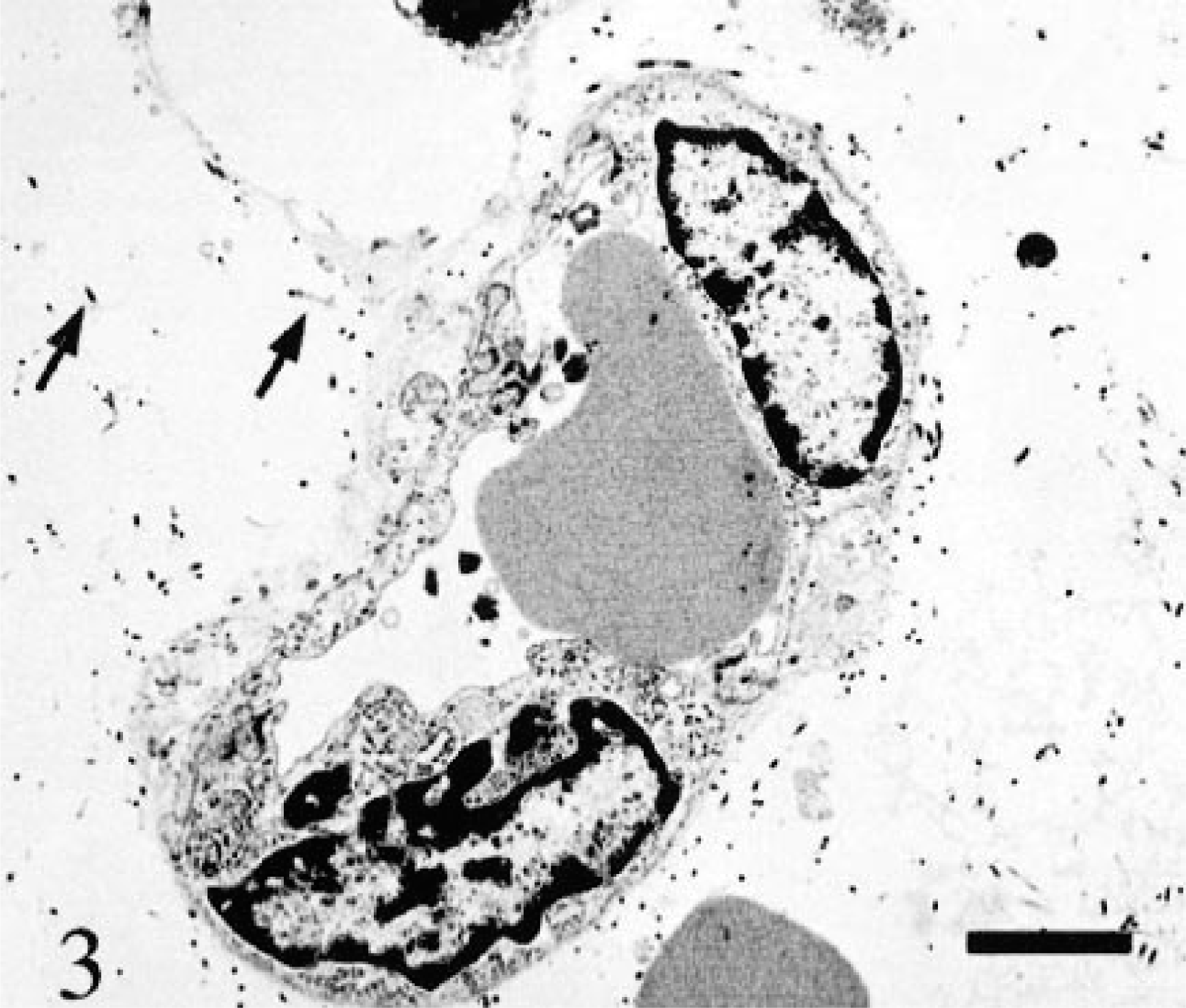
Skin. TEM. Immature dermal capillary. Note the lack of pericyte coating and widely spaced extracellular dermal collagen fibrils (arrows) in the edematous dermis. Bar = 2 µm.
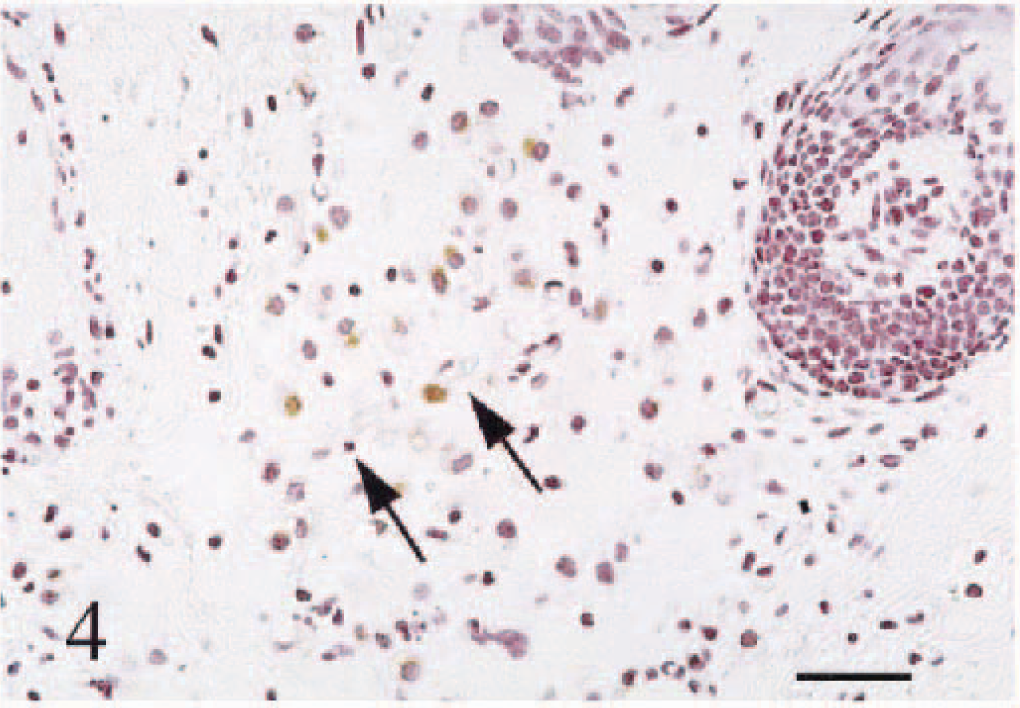
Skin. PRRS virus immunohistochemistry. Hematoxylin counter stain. Note PRRSV-positive macrophages adjacent to immature capillaries. Arrows indicate a dermal capillary. HE stain. Bar = 50 µm.
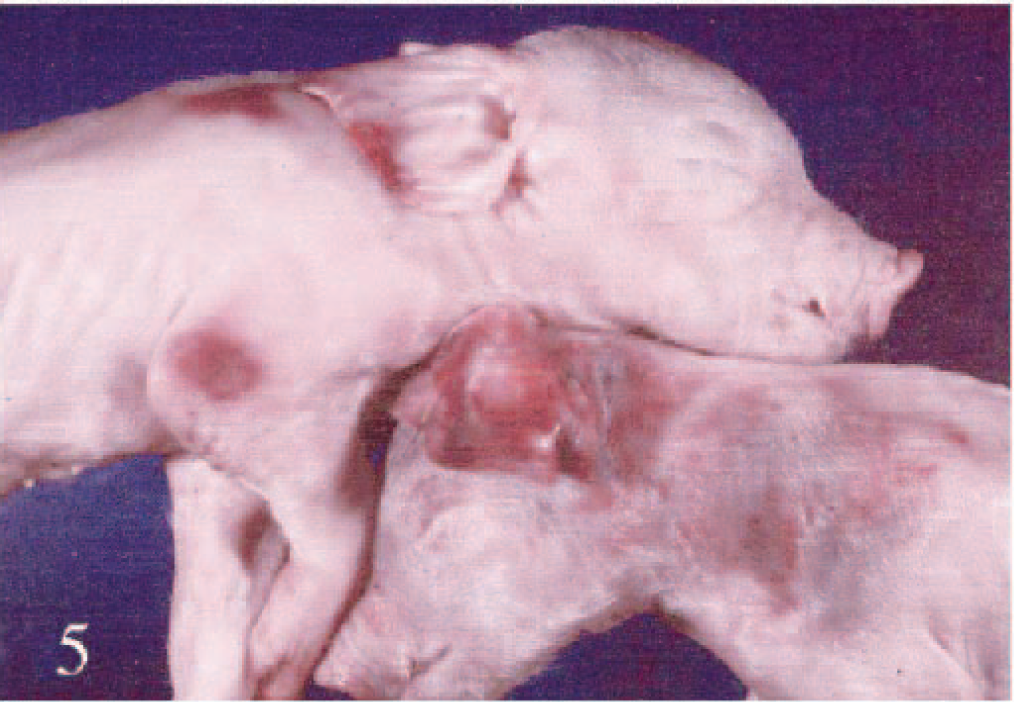
Stillborn pigs exhibiting focally extensive cutaneous hemorrhages.
A 2,500-sow PRRSV-positive breeding-farrowing unit experienced an acute outbreak of abortions and death in adult sows. Historically, this farm had experienced periodic spikes in PRRSV abortion, generally affecting 30 to 90 sows with little or no associated adult mortality. In this outbreak, sows initially developed anorexia with marked pyrexia, followed 24–48 hours later by abortion. The sows often exhibited marked lethargy, red-blue discoloration of the skin, and purulent vulvar discharge 12–24 hours before aborting. More than 45 sows died during or soon after the onset of the acute febrile illness. The majority of sow deaths occurred 24–72 hours after the abortion. More than 400 of the 2,500 sows aborted over a 4-week period. During this period, neonatal pigs were often born weak and lethargic; there was also a corresponding increase in neonatal and preweaning deaths. Full-term stillborn piglets and live-born, weak piglets occasionally had single or multiple coalescent dark reddish-blue cutaneous hemorrhages, which were most notable in the pinnae, cranium, and lateral cervical and shoulder areas (Fig. 5). Fewer than 10% of the aborted and live-born litters were affected with these hemorrhages. Affected litters, however, tended to have multiple (2–5) pigs affected with these hemorrhages. Less severely affected pigs had many small (2–10 mm) hemorrhages evident on the skin of the lateral neck and hind limbs. Cutaneous hemorrhages in less severely affected surviving piglets were spontaneously resolved within a week of birth. Narrow annular hemorrhages occasionally were visible on the tails of affected piglets. The skin overlying the large cutaneous hemorrhages was often fragile and easily removed.
Serum was submitted from eight recently aborting sows and six neonatal pigs, 1–4 days old, for PRRSV isolation. Three full-term stillborn pigs were submitted for necropsy. PRRSV was isolated from the serum of five of eight acutely febrile aborting sows and from pooled serum collected from weak neonates. The PRRSV isolate exhibited a 1-4-2 pattern of restriction fragment length polymorphism of ORF5, consistent with other strains of PRRSV associated with acute severe PRRS outbreaks (Mengeling and Lager, personal communication). 6
Microscopic examination of skin lesions from the piglets depicted in Fig. 5 revealed extensive areas with acute subcutaneous and dermal hemorrhage, as well as an underlying network of tortuous capillary structures lined by plump fusiform cells. Dermal and subcutaeous extracellular matrix was edematous to myxomatous and faintly basophilic, and it showed pericapillary and perivenular clear zones (Fig. 1). Perivascular clear zones often contained loosely arranged infiltrates of large, foamy macrophages (Fig. 2). Similar, but less extensive, angioproliferative changes were seen in the perinodal connective tissue of the mesentery and in perirenal fat and pericardial fat. No significant lesions were observed in other tissues. Immunohistochemistry revealed PRRSV antigen in most of the perivascular macrophages (Fig. 4). Immunohistochemistry and in situ hybridization for PCV-2 was negative in the skin, lymph node, and spleen from the two affected pigs examined.
Transmission electron microscopy revealed that immature capillaries were lined by markedly hypertrophic endothelium with a lack of discernible basal lamina or encircling pericytes (Fig. 3). Collagen fibrils within the adjacent, markedly edematous dermis were widely spaced and haphazardly arranged. Larger vessels were lined by similar appearing endothelial cells, which were encircled by a layer of swollen elongate mesenchymal cells, suggestive of smooth muscle cells; these were in turn surrounded by widely spaced collagen fibrils. Perivascular macrophages had one or two large cytoplasmic vacuoles containing granular amorphous debris or membrane-bound organelle fragments. The cytoplasm was organelle rich with abundant, rough endoplasmic reticulum. Nuclei were peripherally located, with marked convolution and indentation of the nuclear membrane and abundant marginated heterochromatin. Small clusters of individual collagen fibrils were adjacent to the plasma membrane.
The lesions in this case of PRRSV abortions are atypical because gross lesions usually are absent from fetuses, and vascular proliferative lesions have not been reported previously. Gross fetal lesions that have been reported with PRRSV infection include umbilical cord edema and segmental hemorrhage, as well as edema of the eyelids and neck. 5 Microscopic examination of affected umbilical cords can reveal lymphohistiocytic or necrotizing arteritis, or both. Segmental lymphohistiocytic arteritis also has been described in multiple visceral organs, including the heart, lung, and kidney. In some cases, perirenal edema, mesenteric edema, and abdominal and thoracic effusions have been associated with vasculitis. 5 Uteroplacental lesions described in experimentally inoculated sows include endometrial lymphohistiocytic perivasculitis or vasculitis, placental microseparation, and edema fluid accumulation in the intervening chorionic–endometrial space. 5
Angiogenesis involves growth of primitive endothelium from preexisting vessels under the influence of multiple stimulatory growth factors. 3 Proliferation and maturation steps are mediated by a combination of soluble factors that ultimately lead to development of microvascular structures encircled by perithelial and smooth muscle cells. Initially, angiogenesis involves marked increases in vascular diameter and endothelial permeability, which allows exudation of fibrin to provide a matrix for support and attraction of endothelial cells. 3 Basic fibroblastic growth factor (bFGF) and vascular endothelial growth factor (VEGF) have been shown to mediate endothelial chemotaxis and migration. 13 Endothelial proliferation begins at the tips of the migratory endothelial strands, followed by development of lumina by endothelial covering of preexisting fluid-filled spaces or by coalescence of intracytoplasmic lumina within the endothelial cells. 13 Final events in vascular maturation include development of a basal lamina and acquisition of pericyte and smooth muscle coats; these provide increased vascular resistance to mechanical injury. 1 Recent findings suggest that VEGF and bFGF induce formation of the pericyte coating as well as protecting against vascular regression, by inducing production of the apoptosis inhibitor survivin. 3,8
Macrophages play an important role in mediating vascular proliferation through production of endothelial migration and proliferation factors, secretion of protease to degrade the extracellular matrix, and secretion of monokines that stimulate endothelial protease secretion for extracellular matrix degradation. 12 Macrophages are capable of producing a wide variety of angiogenic growth factors and monokines, such as bFGF, transforming growth factor α and β, platelet-derived growth factor, and VEGF, along with interleukin-1 (IL-1), interleukin-6 (IL-6) and interleukin-8 (IL-8). 12 Angiogenic macrophage activation is different from inflammatory activation; hence, standard markers for macrophage activation may not reflect their true angiogenic status. Macrophages become angiogenic during hypoxia; elevated lactate, pyruvate, or hydrogen ion concentration; or under cytokine profiles seen under healing wound conditions or chronic inflammation. 4,12 Alternatively activated macrophages (IL-4/glucocorticoid-induced macrophages) show a threefold increase in angiogenic activity above that demonstrated by classically activated (interferon-γ–induced) macrophages. 4 Likewise, lesions with higher levels of alternatively activated macrophages tend to be better vascularized. The difference in angiogenic reaction of these two subsets of macrophages is presumably the result of differences in angiogenic factor production between the classically and alternatively activated macrophages. 4 Macrophages are also capable of producing antiangiogenic factors, such as thrombospondin-1. 12
Histologic and ultrastructural vascular changes in the pigs described in this report are consistent with a multifocal angiogenic response in the skin and serosal connective tissue of the abdominal and thoracic cavity. The focally extensive cutaneous hemorrhages are likely a consequence of birth-associated trauma and rupture of these immature and fragile vessels. The marked edema, extracellular matrix alterations, and endothelial morphology are consistent with an ongoing vasculoproliferative event. The spatial location of these macrophages and their number and altered morphology suggest that they may modulate this unusual vascular proliferation. Although the cause of these vasculoproliferative lesions is uncertain, the temporal association of the lesions with the PRRSV outbreak and the localization of PRRSV antigen within macrophages adjacent to the proliferating capillaries suggest that PRRSV played a role in the development of these unusual lesions. Activated and immature porcine macrophages are preferred cells for PRRSV infections; thus, it is plausible that PRRSV infection altered the secretion of angiogenic cytokines in these perivascular macrophages, thereby stimulating angiogenesis. 11 Alternatively, PRRSV-induced umbilical vasculitis, placentitis, or premature placental separation may have induced sublethal fetal hypoxia, resulting in an induction of a cascade of angiogenic factors leading to this particular vasoproliferative response. Inhibition of the normal regression of proangiogenic stimuli, seen in the perinatal and postnatal period, may also be a plausible explanation for the development of these vascular lesions. 6
Porcine dermatitis and nephropathy syndrome (PDNS) is a systemic necrotizing vasculitis seen in weaned and finisher pigs that is associated with both PRRSV and PCV-2 infection. 10,14 The cutaneous and multisystemic leukoclastic vasculitis and fibrinous glomerulonephritis typically reported with PNDS is substantially different from the lesions seen in this case. 10,14 The negative in situ hybridization and immunohistochemistry results in these cases suggest that PCV-2 was not present in these pigs at the time of abortion. To our knowledge, this is the first report of these proliferative vascular lesions associated with PRRSV abortion in pigs.
Footnotes
Acknowledgements
This manuscript represents MAFES publication J9784.