
Abstract
Formalin-fixed paraffin-embedded archival tissues of seven adult cats of both sexes that died after being experimentally infected with Yersinia pestis were examined light microscopically to characterize the lesions. The cats were exposed in two groups using two routes of infection: ingestion of Y. pestis-infected rodent or a subcutaneous injection of Y. pestis to simulate a flea bite. Immunohistochemistry was performed on tissues from all organ systems from a representative cat from each group to determine the distribution of Y. pestis bacilli during infection. In all seven cats, bubonic plague lesions were seen. The lesions of pneumonic plague were present in two cats. Septicemic plague was confirmed in all seven cats by bacteriologic culture. Aggregations of bacteria were seen in lymphoid tissue in all cats and in lung tissues from the two cats with pneumonic plague. The most consistent histologic finding was necrosuppurative inflammation in the lymph nodes. Invariably, Y. pestis bacteria were present in large numbers at affected sites. Orally infected cats had more numerous lesions in the lymph nodes of the head and neck regions. These experimentally induced cases of feline plague document that cats are unique among carnivores in exhibiting bubonic, pneumonic, and septicemic plague following exposure to Y. pestis. The lesions of the orally infected cats were consistent with those previously described for naturally occurring Y. pestis infections in cats and corroborate the contention that cats most commonly contract plague by eating Y. pestis-infected rodents and not via flea bite. The histopathology of Y. pestis disease in these cats is comparable to that described for human plague.
Keywords
Yersinia pestis is the causative agent of plague, a gram-negative bacterial disease occurring principally in rodents and fleas and occasionally in nonrodent mammals including domestic cats and humans. 21 Y. pestis is established in certain ecological niches worldwide, including the southwestern United States. 16 Historically, the plague has been associated with large human epidemics, particularly in medieval Europe. Although there are historic suggestions of cats being susceptible to plague, 27 it was not until the latter part of the 1900s that plague was recognized as a significant disease in cats and that infected cats could serve as a threat to human health. 16 Cats are rare among carnivores in that they are unusually susceptible to disease after Y. pestis exposure. In 1999, a total of 42 plague cases in cats (Felis cattus) were confirmed from the three states where plague is most common: California (7), Colorado (9), and New Mexico (26) (C. Fritz, R. E. Enscore, and P. Reynolds, personal communication). Many cats may succumb to the disease without ever being identified, or examined, or counted, often dying in secluded areas. 10 The published mortality rates for plague in this species is approximately 33%. 9 15 26 Because of the number of cases of feline plague, this disease is becoming increasingly important as a veterinary medical concern and as a zoonotic link to humans. 12 14 In both cats and humans, clinical signs include fever, lethargy, myalgia, palpable lymphadenopathy, and anorexia with an onset 2–5 days following exposure. An average of 11.4 cases of human plague are reported each year in the United States, and about 2,000 cases/year are reported worldwide. 11 Several human plague cases in the southwestern USA reportedly originated in cats and were transferred via cat bite, scratch, or contact with infected bodily fluids, such as respiratory droplets. 8 30–32
In this study, archival tissues from seven cats with experimentally induced fatal plague were used to characterize the histopathology in affected organs. These seven cats were selected from 20 cats exposed to Y. pestis either orally (to simulate ingestion of infected prey) or subcutaneously (to simulate flea bite). In all cases, bubonic lesions were most prominent. Affected lymph nodes were characterized by necrosuppurative inflammation with an effacement of normal nodal architecture and abundant Y. pestis organisms, a characteristic hallmark of severe plague infection in humans and cats. Other tissues with consistent involvement included lung and perilymphnodal connective tissue.
Materials and Methods
Bacterial strain and exposure
All cats and mice were infected with virulent Y. pestis strain NM77-58. Two exposure routes were employed. Group I cats were fed a 6-week-old laboratory mouse that died of a Y. pestis infection. Rodent inoculations were used to simulate the route purported to be the most common form of natural infection in cats, which is ingestion of a dead, or moribund, infected rodent. Group II cats were given a subcutaneous injection of 3.78 × 105 Y. pestis bacteria in the right inguinal area. The dosage was calculated to mimic the amount of bacteria that would be delivered by multiple flea bites (one bite may deliver up to 15,000 bacteria). 3
Animals and tissues
In part of a previous series of experiments, 20 adult cats, both males and females, were divided into two experimental groups (P. W. Gasper, unpublished material). There were originally 16 cats in group I, of which six died (38% mortality), and four cats in group II, of which two died (50% mortality). Gross and histopathologic examination and bacteriology results revealed that all deaths were a result of the Y. pestis infection. All eight fatalities occurred between days 4 and 9 postexposure. Seven of the fatally infected cats were used for the present study.
The animals were obtained from the City of Denver, Animal Control Section, and were housed in the Laboratory Animal Research Center at Colorado State University. All animals were handled according to the guidelines and protocols established by the Colorado State University Animal Care and Use Committee. At the Center, cats were held for quarantine and conditioning according to standard operating procedures of the center. On receipt, any cat found to be palpably pregnant or to have a significant health problem was rejected. Cats that were not pregnant and that were healthy or had minor treatable problems were held in a limited access room in individual cages and treated appropriately. All cats retained were vaccinated with a feline viral rhinotracheitis–calici virus–feline panleukopenia vaccine (FVR-CP, Pitman-Moore, Lincoln, NE) according to standard protocol. Each cat was treated with niclosamide (for tapeworm), metronidazole (for Giardia), sulfadimethoxine (for Coccidia spp.), and pyrantel pamoate (for nematodes). Animals were observed daily. Each cat received a complete physical examination, including tests for parasites, blood and serum chemistry evaluations, and complete blood counts. Cats were also tested for the presence of feline leukemia virus and for antibodies to Y. pestis by passive hemagglutination inhibition assay. Healthy cats without antibodies to Y. pestis were released as experiment ready after a minimum of 2 weeks under quarantine and treatment. Complete necropsies were performed on each animal that died during the experiment. 15
Histopathology
Tissues were fixed in 10% neutral buffered formalin, processed routinely, and embedded in paraffin. Blocks were sectioned at 5 µm and routinely stained with hematoxylin and eosin (HE). One infected lymph node section from cat No. 4 was additionally stained with Brown and Hopps Gram stain. Samples from a complete set of tissues from each animal were examined by light microscopy.
Immunohistochemistry
Tissues from two representative cats (No. 2 from group I and No. 7 from group II) and infected lymph nodes from cat No. 4 were examined immunohistochemically. Five-micrometer-thick paraffin-embedded tissue sections were prepared on positively charged microscopic slides (Superfrost/Plus, Fisher Scientific, Pittsburgh, PA). The sections were deparaffinized and rehydrated. For tissues from cat Nos. 2 and 7, the staining series following deparaffinization was carried out using an automated cell staining machine (Version 2.0 P.D. Optimax Automated Cell Staining System Plus, Biogenex, San Ramon, CA). Tissues were washed between each step using phosphate-buffered saline and deionized water. All incubations were done at room temperature. Following incubation with a peroxidase inhibitor and a protein block (Biogenex), a polyclonal antibody raised against the fraction 1 capsular antigen of Y. pestis (obtained from Dr. J. Ellis) was incubated on the tissues for 1 hour at dilutions of 1:1,000 (cat No. 4 lymph nodes and cat No. 2 lung, spleen, and tonsils) or 1:3,000 (all other tissues). Tissues were then incubated with biotinylated goat anti-rabbit IgG link (Kit GP000-UR, Biogenex) for 20 minutes. Bacteria were visualized using a streptavadin–horseradish peroxidase-conjugated label (Biogenex) followed by the imaging substrate (AEC chromagen, Biogenex) and Mayer's hematoxylin (Biogenex) counterstain. A heat-fixed smear of strain KIM5 Y. pestis grown at 37 C (provided by Dr. L. Lindler, WRAIR, Washington, DC) was used as a positive control.
Results
Table 1 provides a summary of the histologic findings of the seven cats that died of experimentally induced plague.
Summary of histopathologic findings∗ in seven cats with experimentally induced plague.
+ = present; 0 = not present; 0 = tissue not examined histologically.
Gross pathology
All orally challenged cats (group I) exhibited marked enlargement (approximately 7–10-fold increase in size) and abscessation of the medial retropharyngeal lymph node. Less consistently involved were the tonsils and submandibular and cranial thoracic lymph nodes. Cat No. 4 had similar changes in the ileocecal lymph node. There was dark reddish discoloration of tissues within 7–20 mm of affected nodes. Focal areas of petechiation were noted throughout all tissues, with larger (5–10 mm) ecchymoses present in the lungs and spleens of these cats.
Enlarged lymph nodes were not palpable in the animals that died following subcutaneous challenge (group II), although Y. pestis was cultured from several lymph nodes in the head and neck region. Large (12 × 5 × 3 cm) subcutaneous dissecting abscesses were noted in both of the group II animals that died. Extensive extravasation of blood and dark reddish black discoloration of the muscles of the thorax, abdomen, and proximal hind limbs were also noted. Cat Nos. 6 and 7 exhibited splenic enlargement and adrenal hemorrhages.
Histopathology
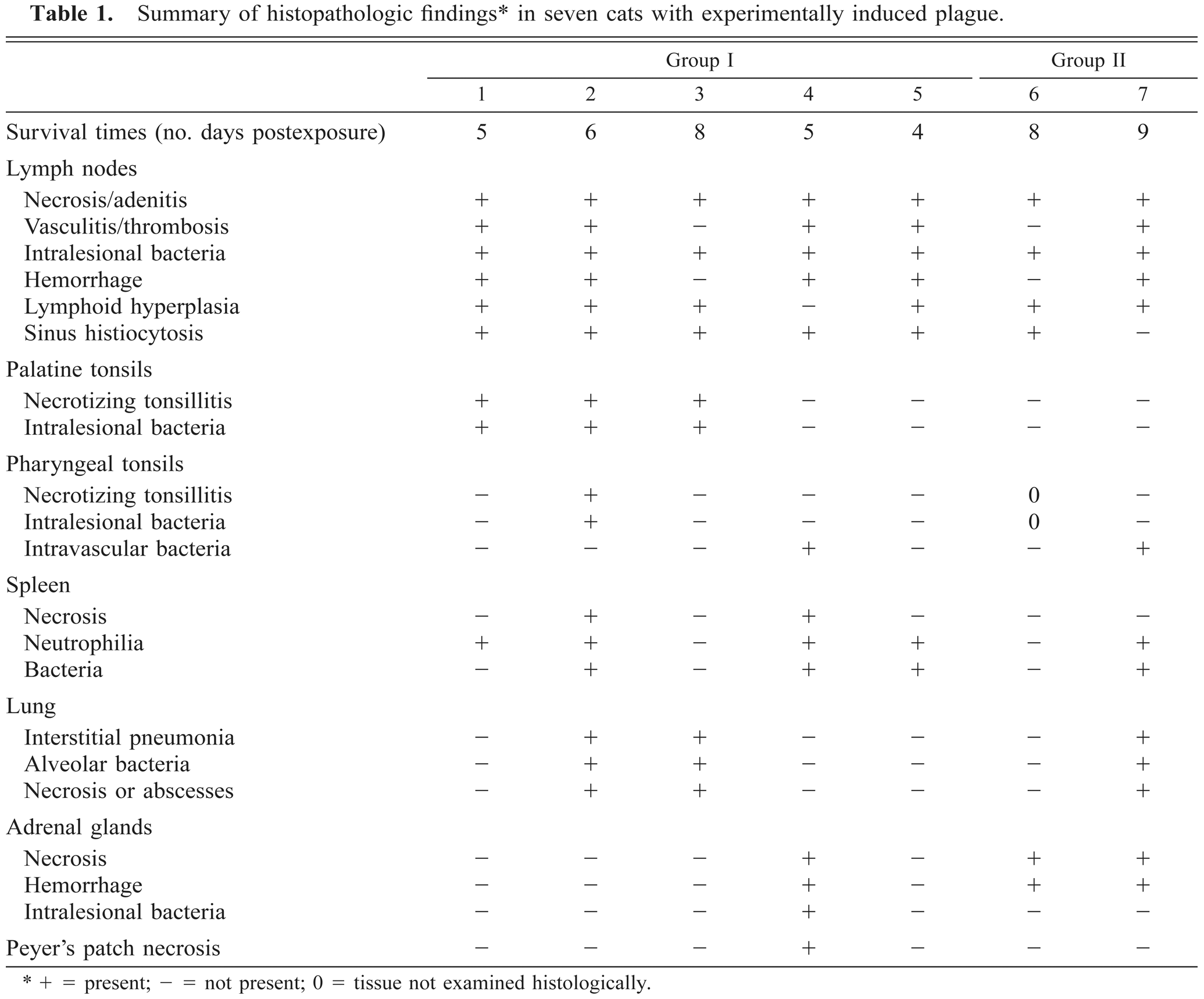
The most consistent changes were in the anterior cervical, medial retropharyngeal, submandibular, and cranial thoracic lymph nodes. All cats had lesions in at least one of these nodes. In most cats, lymph node architecture was markedly altered by acute necrotizing inflammation. Lymph nodes were variably effaced by hemorrhage, edema, and fibrin admixed with eosinophilic cellular and karyorrhectic debris. Numerous viable and degenerate neutrophils and large aggregates of 1–3-µm gram-negative bacilli were present (Figs. 1, 2). Hemorrhage, inflammatory debris, and bacteria often extended beyond the lymph node capsule and infiltrated adjacent fibroadipose tissue. The vascular media of intranodal arterioles frequently was infiltrated by neutrophils and less often was necrotic. There was widespread endothelial necrosis and loss within both venules and arterioles in these lymph nodes. Numerous small venules and fewer arterioles contained fibrinocellular thrombi.
Lymph node; cat No. 4. Massive numbers of bacteria, admixed with macrophages and degenerate and necrotic neutrophils, expand the medullary sinus. HE. Bar = 25 µm.
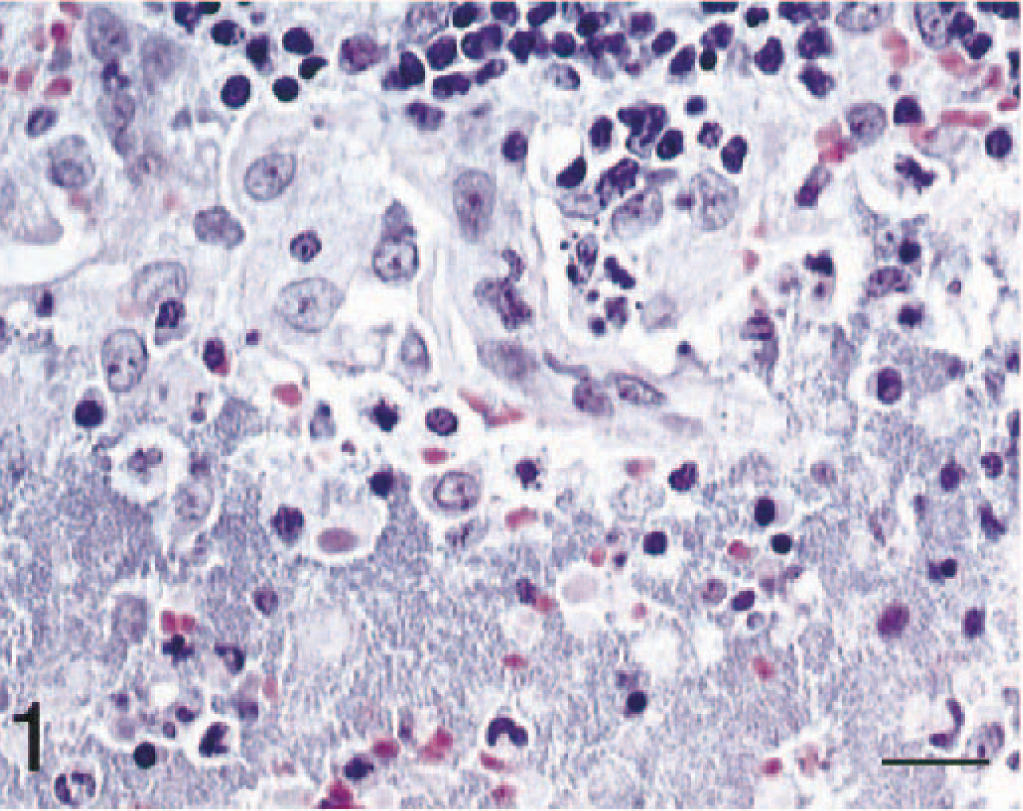
Lymph node; cat No. 4. Same lymph node as in Fig. 1. Numerous immunolabeled bacteria are present. Streptavidin–biotin–peroxidase complex method, Mayer's hematoxylin counterstain. Bar = 25 µm.
Mesenteric lymph nodes in two cats (Nos. 2 and 5) also were acutely inflamed but to a lesser degree than those in anterior cervical areas, and they generally contained fewer bacteria. Often lymph nodes exhibited moderate to marked lymphoid hyperplasia and varying degrees of sinus histiocytosis and draining hemorrhage.
Significant pulmonary lesions were found in three cats, two from group I and one from group II. In the most severely affected lung (cat No. 2), there was a diffuse interstitial pneumonia. Edema, strands of fibrin, and numerous neutrophils and macrophages expanded the alveolar septa, frequently filling alveolar lumina. Alveolar macrophages sometimes contained phagocytosed erythrocytes, necrotic cellular debris, and rarely bacteria. Large aggregates of bacilli were randomly distributed in interstitial spaces and within alveoli (Figs. 3, 4). Multifocally within the lung parenchyma there were coalescing areas of necrosis that obliterated the normal architecture and in some areas formed large abscesses. Abscesses were characterized by a central area of lytic necrosis and degenerate neutrophils, which blended peripherally with numerous bacteria entwined in fibrin and hemorrhage. Within the more severely affected portions of lung, bronchiolar walls were necrotic, and bronchiolar lumina contained hemorrhage, fibrin, necrotic cellular debris, and bacteria. A section of trachea contained hemorrhage and numerous bacteria. Lungs of less severely affected cats had a few scattered focal necrotic areas containing degenerate neutrophils, fewer macrophages, and few bacteria.
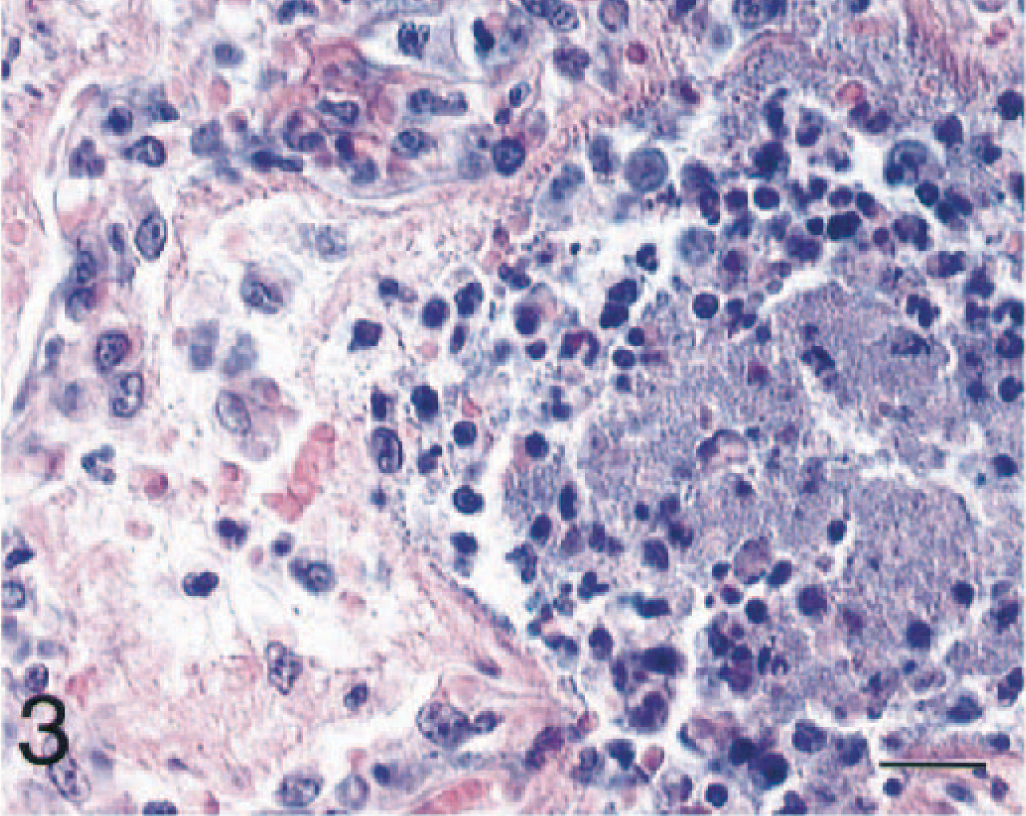
Lung; cat No. 2. The alveolus is filled with numerous bacteria, degenerate and necrotic neutrophils, fewer macrophages, and fibrin. Note expanded alveolar wall (upper left). HE. Bar = 25 µm.
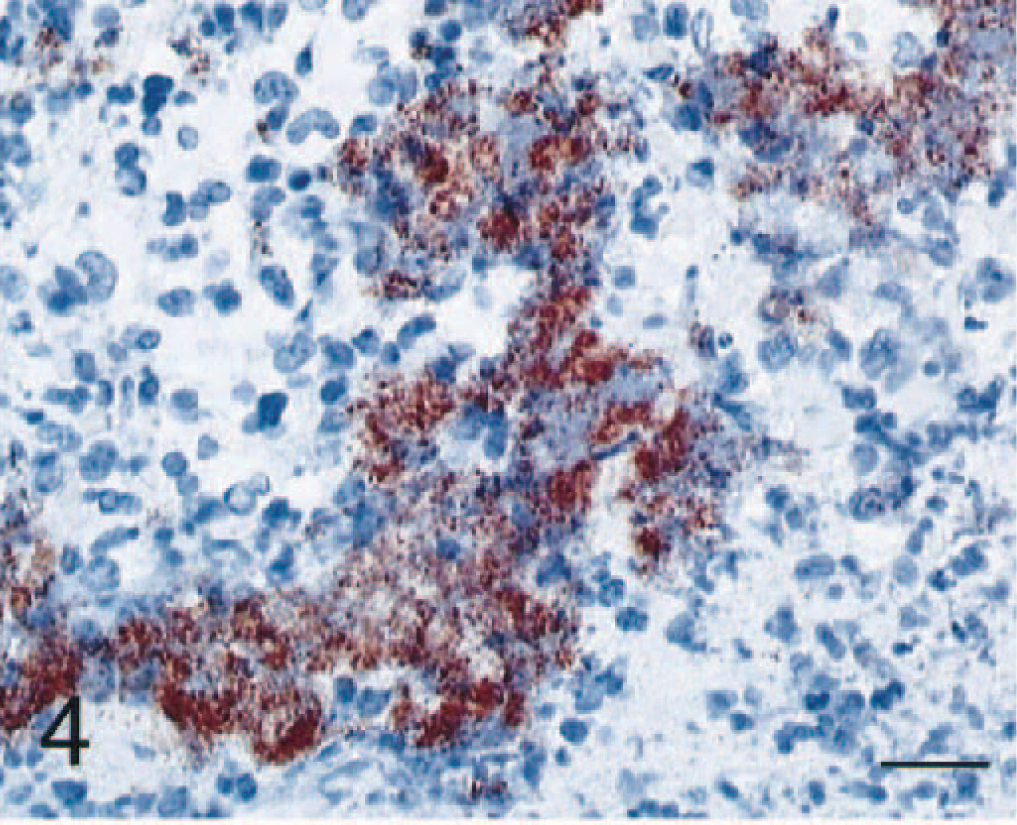
Lung; cat No. 2. Note aggregates of immunolabeled bacteria. Streptavidin–biotin–peroxidase complex method, Mayer's hematoxylin counterstain. Bar = 25 µm.
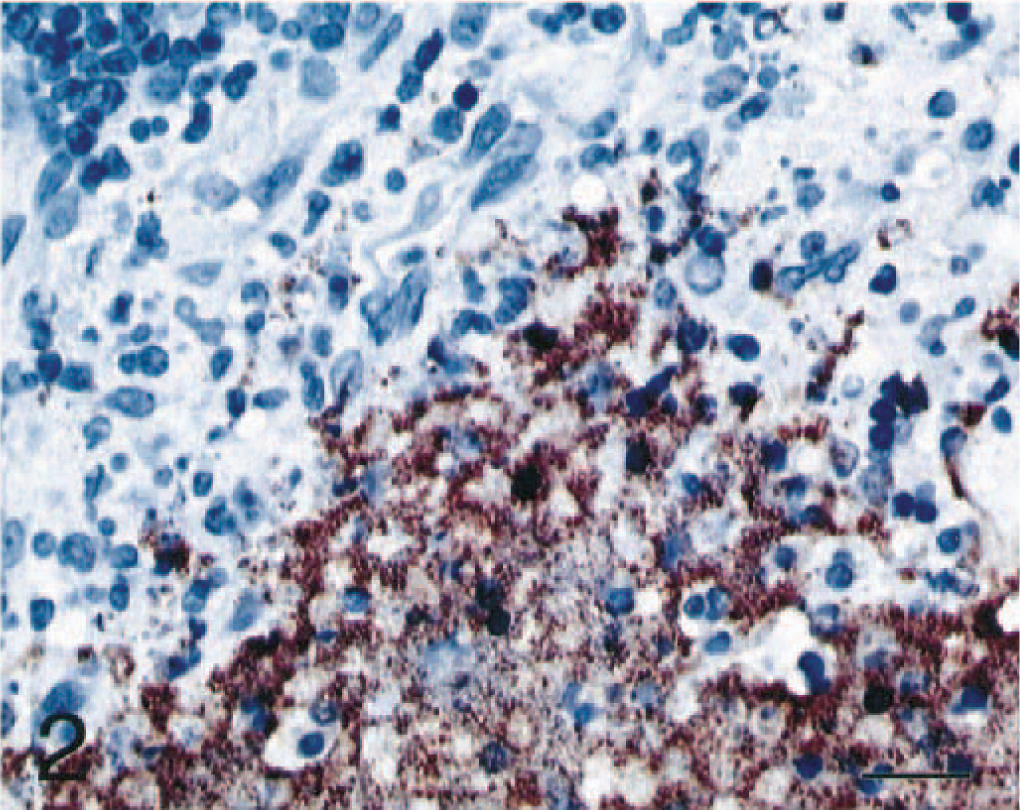
In three cats (Nos. 1–3), severe acute inflammation, widespread necrosis, and large aggregates of bacteria were present in palatine tonsils and, in one cat (cat No. 2), in the pharyngeal tonsils (Figs. 5, 6). Peritonsillar soft tissue was expanded by edema, fibrin, and hemorrhage, and bacteria were present within the perimyseal fascia of adjacent facial skeletal muscle bundles.
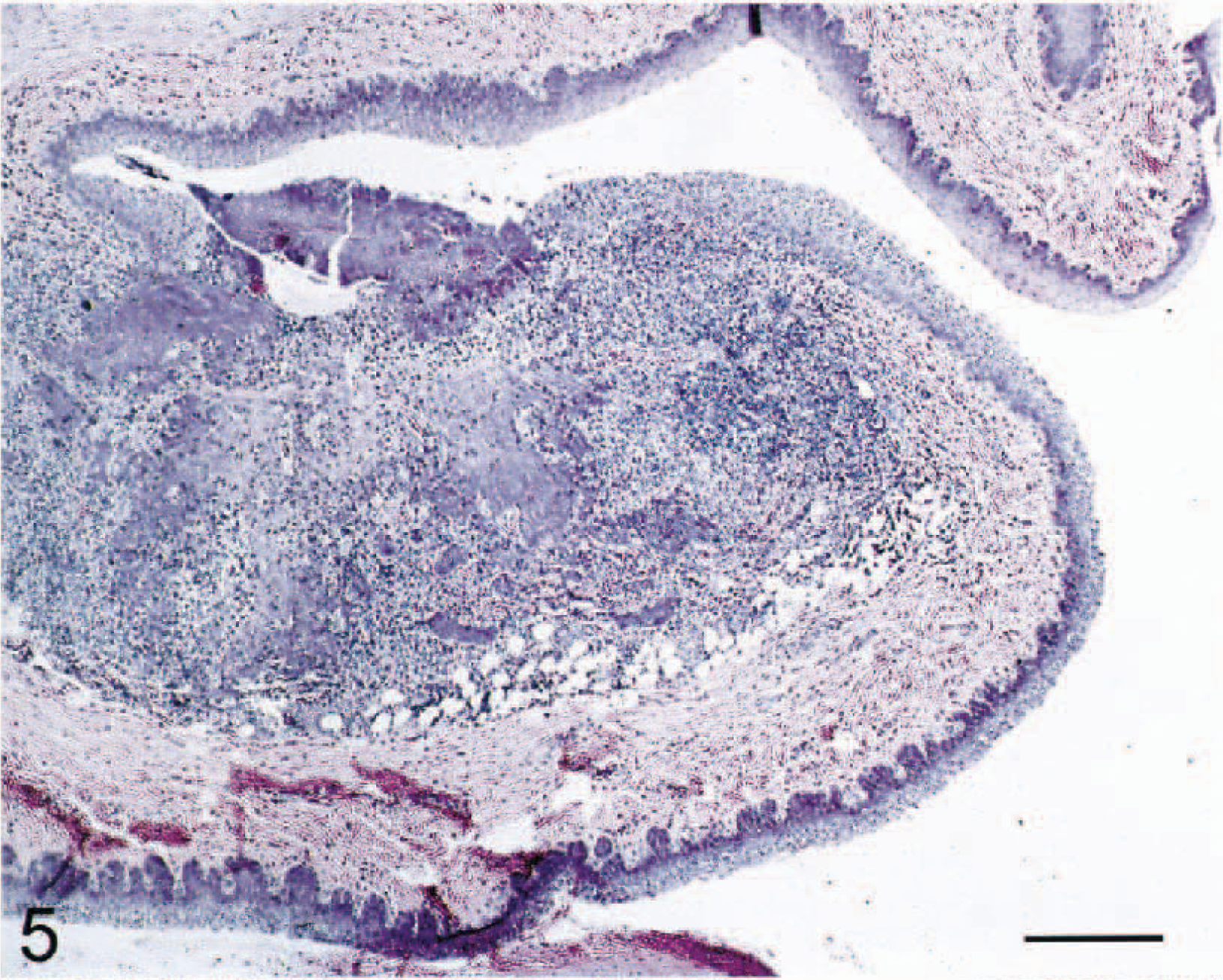
Pharyngeal tonsil; cat No. 2. There is necrosis of lymphoid tissue and disruption of the overlying tonsillar epithelium. HE. Bar = 210 µm.
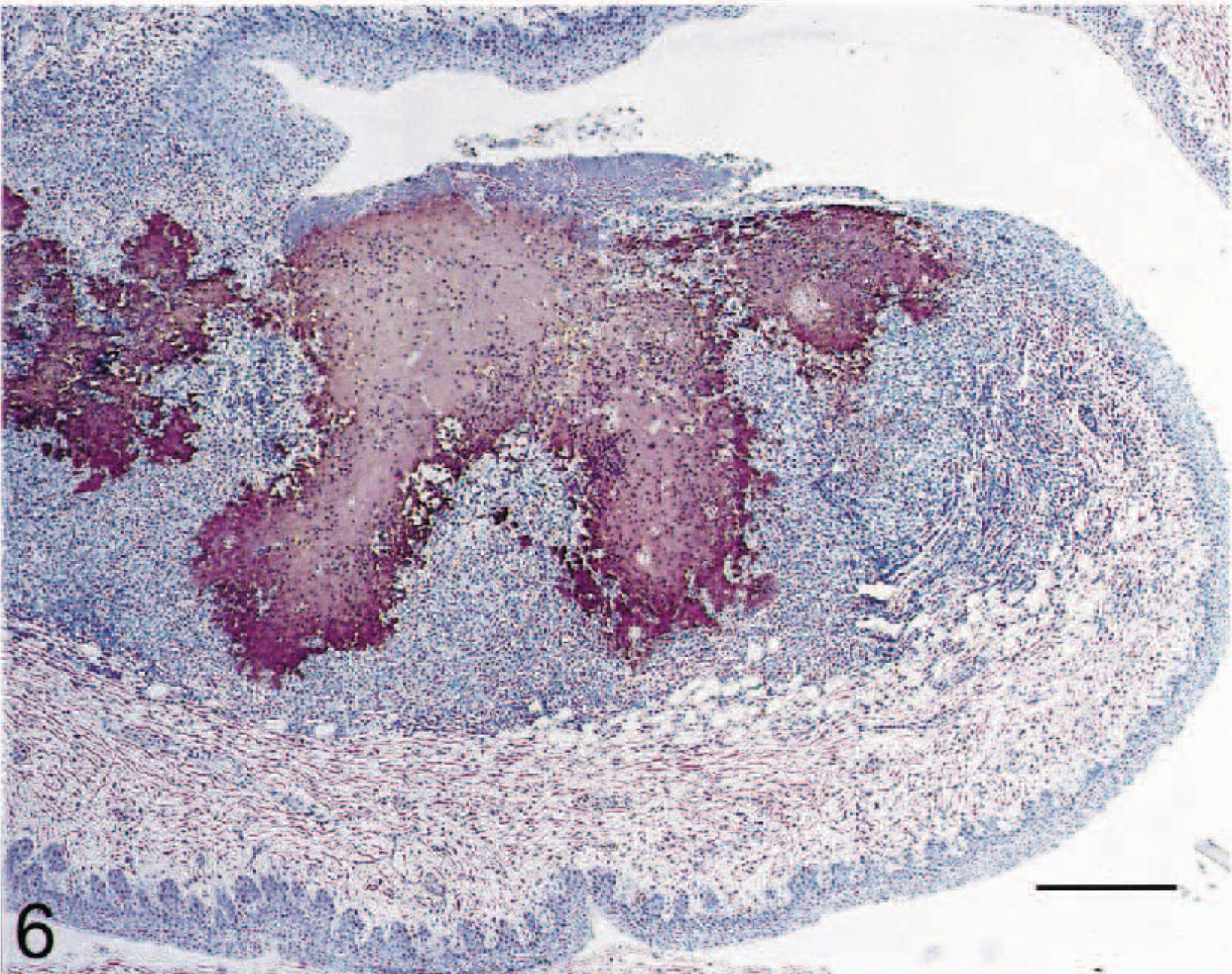
Pharyngeal tonsil; cat No. 2. Same tonsil as in Fig. 5. There is intense immunolabeling of necrotic lymphoid areas. Streptavidin–biotin–peroxidase complex method, Mayer's hematoxylin counterstain. Bar = 210 µm.
Large numbers of bacteria were found in the lumina of blood and lymphatic vessels both within and near affected lymph nodes. Two cats (Nos. 2 and 4) had acute fasciitis with few bacteria present adjacent to a large peripheral nerve. Spleens of four cats (Nos. 2, 4, 5, and 7) contained few to many bacteria and suppuration to a degree that approximately paralleled the number of bacteria present.
Adrenal glands in four cats (group I: Nos. 4 and 5; group II, Nos. 6 and 7) had moderate to marked multifocal hemorrhage within the cortex. Small focal accumulations of necrotic cells were present in some areas of hemorrhage, and in one cat few bacteria were present in these lesions. An additional cat in group I (cat No. 5) had hemorrhage and fibrin adjacent to the adrenal gland capsule and extending slightly into the surrounding fibroadipose tissue.
One cat from group I (cat No. 4) had focally extensive necrosis of lymphoid cells in the gut-associated lymphoid tissue of the ileum. The Peyer's patch was replaced by eosinophilic cellular debris that contained numerous 3–5-µm bacilli, whose morphology was consistent with enteric bacterial flora. This cat and one other in the same group (No. 5) had mild to moderate hemorrhage multifocally within the small intestinal submucosa.
Sections of thymus were normal except in one cat (No. 1). In this cat, the thymus contained multiple randomly placed aggregates of foamy macrophages, often accompanied by acicular (cholesterol) clefts. Similar lesions were not found in any other organs of this cat or in any of the other cats, and the significance of this finding was not determined. In the perithymic adipose tissue there was moderate acute necrotizing steatitis, and there was vascular necrosis and thrombosis of nearby small-caliber blood vessels.
No significant lesions were found in any other organs examined histologically.
Immunohistochemistry
Immunohistochemical procedures demonstrated that the bacteria visualized with HE staining were Y. pestis and highlighted the distribution of these bacteria in target organs. Positive staining was found in all lymphoid tissue and lung sections where bacteria were visible by HE staining. In some of the lymphoid tissue where inflammation and necrosis were observed, the staining pattern was strongest at the periphery and became weaker toward the centers of the lesions, in correlation with the viability of the bacterial antigens.
Positive staining also was present in some areas where fewer bacteria were aggregated and were difficult to discern with HE staining alone (e.g., within intramuscular fascia). One section of subcutaneous tissue from a group II cat (No. 7) stained positively for Y. pestis and was likely a sample from the inoculation site.
Discussion
Although the idiosyncratic susceptibility that felines have for Y. pestis infection was first noted in 1905, 27 the present study is the first to provide a thorough description of the histopathology of plague in cats. In a previous prospective study, 26 five cats were exposed to Y. pestis: three cats were fed plague-infected rodents and two cats were injected subcutaneously. All five cats became ill, two died within 6 days, one died on day 20 postexposure, and the remaining two (one orally infected and one subcutaneously infected) survived. Necrotic lung lesions with numerous bacilli and cervical abscesses were mentioned but not described. A retrospective study was conducted on 119 cats with naturally occurring plague infections. 9 Sixty-seven of these cats displayed lymphadenopathy characteristic of bubonic plague. Of these cats, 50 had enlarged submandibular nodes suggestive of a route of infection through the oropharynx. Twelve of the 119 cats had pneumonic plague, nine with neither pneumonic or bubonic plague were confirmed to have septicemic plague, and 35 of the plague cases were unclassified. The mortality rate for this group of cats was 33% and included animals that were treated. Most textbooks list the primary mode of plague transmission between and within mammalian species as via flea bite. However, ingestion of Y. pestis-infected rodents is likely the primary source of exposure for carnivores. 4–6, 16 20 The lesions seen in cats with naturally occurring plague and those experimentally exposed via an infected rodent meal are analogous (lymphadenopthy of the cranial and cervical lymph nodes). Many naturally occurring cases of plague in cats, if not the majority, probably are initiated through ingestion of Y. pestis-infected rodents rather than via flea bite.
Plague is described as an endemic disease of rodents, in which certain species and their fleas are believed to be the natural reservoir for the bacterium. There appears to be a schism between the few descriptions of the lesions of rodents with natural infections and those from other mammals that die from Y. pestis infection. The lesions seen in rodents vary considerably depending on whether the species is a maintenance or enzootic host or whether it is involved in epizootic die-offs or amplification. 22–25, 29 Susceptible mice and other rodents do not typically exhibit bubonic or pneumonic plague lesions when found dead from natural infections. Furthermore, susceptible rodents die more acutely, usually within 3 days as opposed to the 5–10-day period seen in cats and humans (T. Quan and S. Price, personal communication). Rodent species may be more sensitive to the fatal effects of the Y. pestis murine toxin, whereas nonrodent species are not significantly affected by the toxin. 2
Almost all carnivores that have been tested, including black bears, raccoons, badgers, and dogs, either experimentally or in surveys for natural exposure in endemic areas mount a robust immune response that prevents the development of clinical signs or pathologic changes. 4–6, 13 17 20 28 34 Cats and probably black-footed ferrets are an exception in plague susceptibility among carnivores. 15 26 33 Bubonic, pneumonic, and septicemic plague are technically morphologic diagnoses of Y. pestis infection in humans. 21 23 Within veterinary medicine, these classic lesions of plague have only been described in monkeys, cats, and black-footed ferrets (see Table 2). 7 9 33 Cats incur pathologic changes analogous to human lesions, suggesting that the pathogenesis of Y. pestis in these two species is similar.
Plague lesions∗ in carnivores following experimental challenge with Yersinia pestis compared with human lesions.

+ = occurs; - = does not occur.
The presence of numerous intralesional bacteria is a consistent finding in fatal plague. Each of the affected cats exhibited enlargement and abscessation of various lymph nodes, with disseminated petechiation and ecchymosis as the principal pathologic manifestation. Grossly, the infection appeared to be contained within the nodes, which were swollen and hemorrhagic. However, microscopic examination revealed that the massive numbers of Y. pestis within the lymph nodes extended beyond the node borders; bacteria were frequently seen in the extranodal connective tissue. In all the cats studied, Y. pestis escaped the lymph nodes and entered the blood stream, as indicated by positive blood culture data obtained from each cat. Plague organisms were cultured from the livers and spleens from some of the cats in this study at the time of necropsy. 15 However, the positive cultures may have resulted from hematologic contamination; little evidence of colonization of these tissues was seen histologically. Although low numbers of bacteria were present in the spleen of four cats (Nos. 2, 4, 5, and 7), no positive staining was found by immunohistochemistry. Too few bacteria may have been present in replicate tissue sections to obtain a visible immunohistochemical reaction. Alternatively, the splenic bacteria could represent postmortem proliferation of intestinal flora.
There was widespread endothelial necrosis and loss within both venules and arterioles, especially in lymph nodes, with many small venules and fewer arterioles partially or completely occluded by fibrinocellular thrombi. These lesions were noted in all seven cats and are consistent with a fatal gram-negative sepsis and disseminated intravascular coagulopathy.
Because of the severity of the lesions described here, future investigations should focus on the pathogenetic mechanisms that underlie the unique susceptibility of cats to subacute fatal infections by Y. pestis. In contrast to most other carnivores, cats fail to prevent Y. pestis from multiplication within target organs at an early phase of the infection. Cats produce antibodies to the fraction 1 antigen of Y. pestis and to some of its major virulence factors; however, this humoral response is not correlated with survival. 15 Because early response to infection is crucial, a component of the early response mechanism may protect most other carnivores from fatal plague infection.
Footnotes
Acknowledgements
We gratefully acknowledge the technical assistance and skills of S. Kodsi of The Armed Forces Institute of Pathology and Specialist J. Hart in the Veterinary Immunohistochemistry Laboratory at the Walter Reed Army Institute of Research. We also thank D. Miles of The Walter Reed Army Institute of Research for the photography of pathology specimens and Drs. A. Barnes and T. J. Quan for conceptualizing, facilitating, and conducting the experimental studies that provided the archival tissues for this study.