
Abstract
Enzootic intranasal tumor (EIT) appears glandular in type and has recently been classified as an adenocarcinoma of low malignancy. The aim of this study was to characterize the secretion of surface glycoconjugates (GCs) in EIT and in normal respiratory and olfactory mucosae of the goat by means of conventional and lectin histochemistry, in order to shed light on the histogenesis of EIT. Morphologic and ultrastructural investigations showed two growth types of EIT: i.e., tubular and papillary patterns. Conventional histochemistry revealed the presence of neutral and carboxylated GCs in the olfactory glands and in the tubular part of EIT, as well neutral and sulphated GCs in the respiratory glands and in the papillary part of EIT, suggesting that the papillary pattern tumor arises from the respiratory glands, whereas the tubular portion of EIT arises from the olfactory glands. Lectin histochemistry gave further information on the expressed GCs.
Tumors are uncommon pathologies in sheep and goats; however, pulmonary adenomatosis, small intestinal carcinoma, lymphosarcoma, skin squamous cell carcinoma, and enzootic intranasal tumor (EIT) have been described, and EIT seems the most frequent neoplasia. In goats, tumors seem to be less frequent, although some cases of EIT have been reported in recent years. 4 5 10 14 17 23
The enzootic intranasal tumor is a chronic and afebrile disease characterized by a profuse initial seromucous and then purulent nasal exudate associated with clinical signs of respiratory distress. In the late period of the disease, facial swelling, sometimes with seromucous exudation, is found because of nasal bone erosion and skin ulceration. 15 EIT in sheep is distributed worldwide, with a high prevalence in affected ovine flocks. 2 8 12 Instead, EIT in goats has been reported only in Italy, Spain, France, Greece, India, and Canada; in the affected flocks, frequence did not exceed 2–3% of the animals, but tumor occurrence was quite constant. 5 15 Even though this tumor has a low and only local malignancy, it is generally lethal because of its particular position as it enlarges to completely invade the nasal and paranasal cavities.
Histologically, the tumor appears glandular in type. In the past it was classified as an adenopapilloma or adenoma, whereas it has recently been considered as an adenocarcinoma of low malignancy. 5 6 11 23
After numerous etiologic hypotheses, from environmental to genetic factors, a viral cause was demonstrated. Viral-like particles, compatible with a type D retrovirus, were isolated from nasal fluid, and neoplastic tissue and successful experimental transmission of EIT from goat to goat confirmed this hypothesis. 1 3 20
In spontaneous disease, the infection occurs through the respiratory route, and the incubation time is so long that the clinical disease is found in animals that are approximately 1 year old. 23
The complete histogenesis of EIT still remains unknown. Its probable histologic origin could be in the olfactory glands, in the supporting cells of the olfactory epithelium, or in the brush cells of the respiratory epithelium. 7 19 This paper reports on the histochemical characterization of the glandular secretions from EIT and from the normal respiratory and olfactory mucosa of goats.
Materials and Methods
Five Alpine goats with clinical signs of EIT were obtained from flocks in Central Italy. At necropsy, after dissection of the cranium through sagittal medial cutting, samples of neoplastic tissue were removed and fixed in Carnoy's fluid for 24 hours, postfixed in 2% calcium acetate and 4% paraformaldehyde (1:1) for 3 hours, and then dehydrated and embedded in paraffin wax.
Serial sections (5 µm thick) were treated for carbohydrate conventional histochemistry with periodic acid–Schiff (PAS; to detect vicinal hydroxyls), Alcian blue (AB) at pH 2.5 (to reveal acidic groups), Alcian blue–periodic acid–Schiff (AB/PAS; to demonstrate acidic groups and vicinal hydroxyls), low-iron diamine (LID; to detect sulfate and carboxyl radicals) and high-iron diamine (HID; to discriminate sulfate groups). 16 22
Lectin staining was performed as previously described.
9
13
The following lectins were used: Peanut agglutinin from Arachis hypogaea [terminal β-
The respective peroxidase-conjugated lectins were omitted, or their hapten sugars (0.2–0.4 M) were added to control sections. As controls for enzyme digestion, sections were incubated with enzyme-free buffers under the same experimental conditions or with the lectins specific for sugar detached by enzyme pretreatment. Sections from normal olfactory and respiratory mucosae of four goats killed in an authorized abattoir were fixed, processed, and stained with the same methods.
Results
In normal goats, the respiratory mucosa is lined by a pseudostratified columnar epithelium with intermingled goblet cells. The propria-submucosa consists of a loose connective tissue in which tubuloacinar-mixed glands are located. The olfactory mucosa is grossly distinguishable from the respiratory one because of its yellowish pigmentation. Microscopically, it is usually thinner than the respiratory mucosa, and the epithelium consists of three different cell types: sensory, substentacular, and basal cells. Many tubuloacinar olfactory glands producing a serus secretion with occasional mucus-producing cells are located in the propria-submucosa.
The neoplasms were similar in all the cases examined and were classified as low-grade adenocarcinoma. They showed two types of growth patterns: tubular, generally found in the deeper areas; and papillary, present mostly on the surface of the tumor. In the tubular part, there were regularly and irregularly shaped acinar and tubular or cystic structures separated by thin vascularized stroma; the neoplastic cells were pyramidal or cuboidal in shape. The papillary pattern consisted of undulating or branching villous projections with delicate connective tissue stalks, covered by one or several layers of cuboidal to columnar epithelial cells with abundant eosinophilic cytoplasm and round nuclei located in a basal position.
The epithelial lining of both respiratory and olfactory mucosae appeared unstained after conventional histochemical procedures in normal subjects.
Olfactory glands showed strong reactivity to PAS, moderate staining intensity after AB pH 2.5 and LID (Fig. 1a) treatments, and slight reactivity to HID (Fig. 1b). Glands and goblet cells of the normal respiratory mucosa intensely reacted to PAS and AB pH 2.5 staining; LID (Fig. 1c) and HID (Fig. 1d) treatments also elicited strong reactivity.
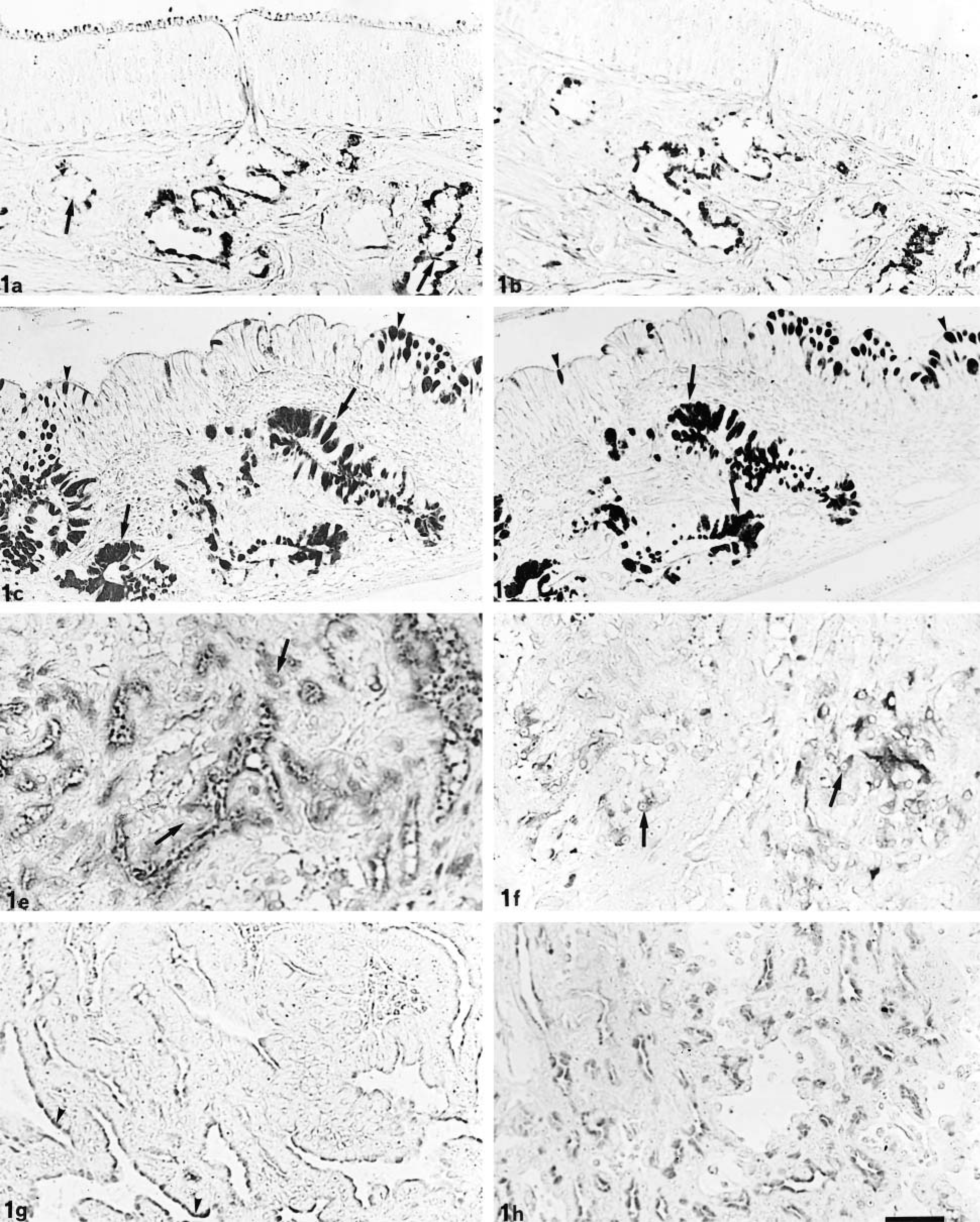
LID and HID staining. Olfactory glands (arrow) react moderately to LID (a). HID reactivity ranges from moderate to slight (b). In the respiratory mucosa, LID (c) and HID (d) react intensely at both goblet cell (arrowhead) and gland level (arrows). In the tubular part of EIT, LID (e) moderately stains some cells (arrow), whereas HID (f) reacts slightly (arrows). In the papillary part of EIT, only the apical cytoplasm of cells appears LID-positive (g) (arrowheads) and, in minor measure, HID-positive (h). Bar = 40 µm.
In the neoplastic tissue, the reactivity of the tubular and papillary parts were different, but no differences were found among the five cases examined. In the tubular part of EIT, all of the neoplastic cells showed diffuse cytoplasmic reactivity; PAS staining was intense, whereas AB pH 2.5 showed moderate staining. Response to LID was moderate (Fig. 1e), whereas reactivity to HID ranged from negative to moderate (Fig. 1f).
In the papillary part of EIT, only the apical cells of the papillary projections showed histochemical reactivity in the supranuclear region. Cells showed strong positivity to PAS, whereas AB pH 2.5, LID (Fig. 1g), and HID (Fig. 1h) gave a reactivity ranging from weak to moderate.
Lectin histochemistry showed weak PNA reactivity on the luminal border of both the olfactory and respiratory glands. Only the tubular portion of EIT showed moderate positivity. Sialidase treatment induced weak reactivity on the olfactory epithelium cell coat and in the apical zone of papillary EIT and enhanced the positivity of respiratory glands. DBA showed binding patterns in both respiratory (Fig. 2a) and olfactory mucosae, as well as in the two growth patterns of EIT. Sialidase digestion affected only the positivity of the respiratory epithelium goblet cells (Fig. 2a) and the papillary portion of EIT (Fig. 2b). In any case, KOH treatment did not modify the activity of sialidase.
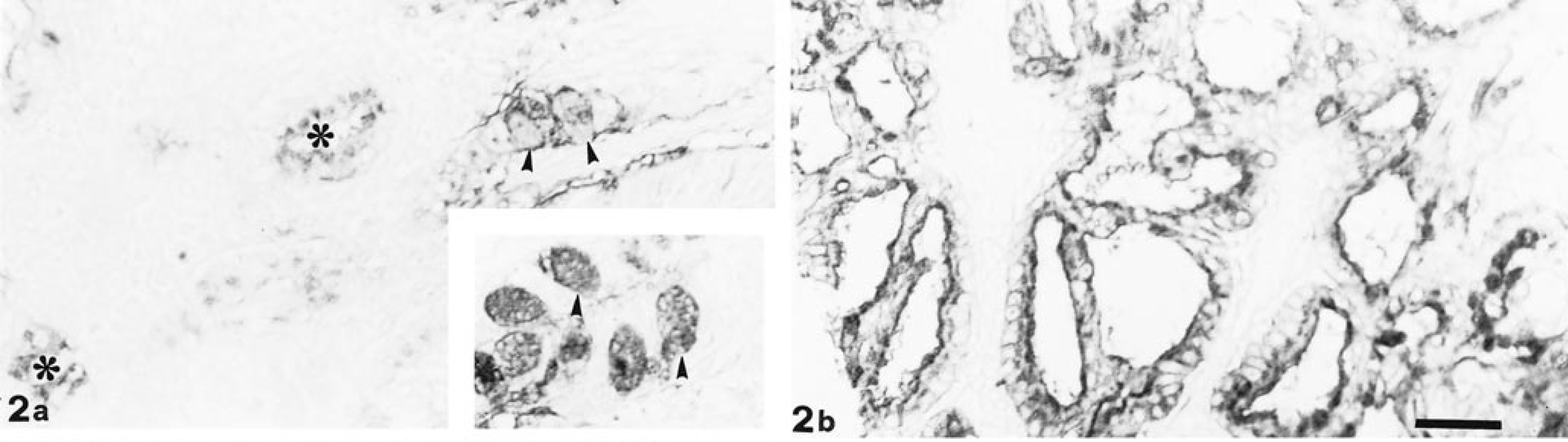
DBA staining. In the respiratory mucosa, goblet cells (arrowheads) and respiratory glands (∗) show reactivity ranging from weak to moderate (a); sialidase digestion enhances goblet cell (arrowheads) positivity (inset). Papillary portion of EIT appears positive with sialidase and DBA treatment (b). Bar = 33 µm.
Con A showed moderate positivity in both normal mucosae and in the two parts of EIT (Fig. 3). WGA showed strong reactivity, especially in the respiratory and olfactory glands (Fig. 4a) and in the tubular portion of EIT (Fig. 4b). GSA II and WGA reactivities were overlapping only in the goblet cells of the respiratory epithelium. Binding sites for GSA IB4 were present in the respiratory and olfactory glands and in the two parts of EIT.
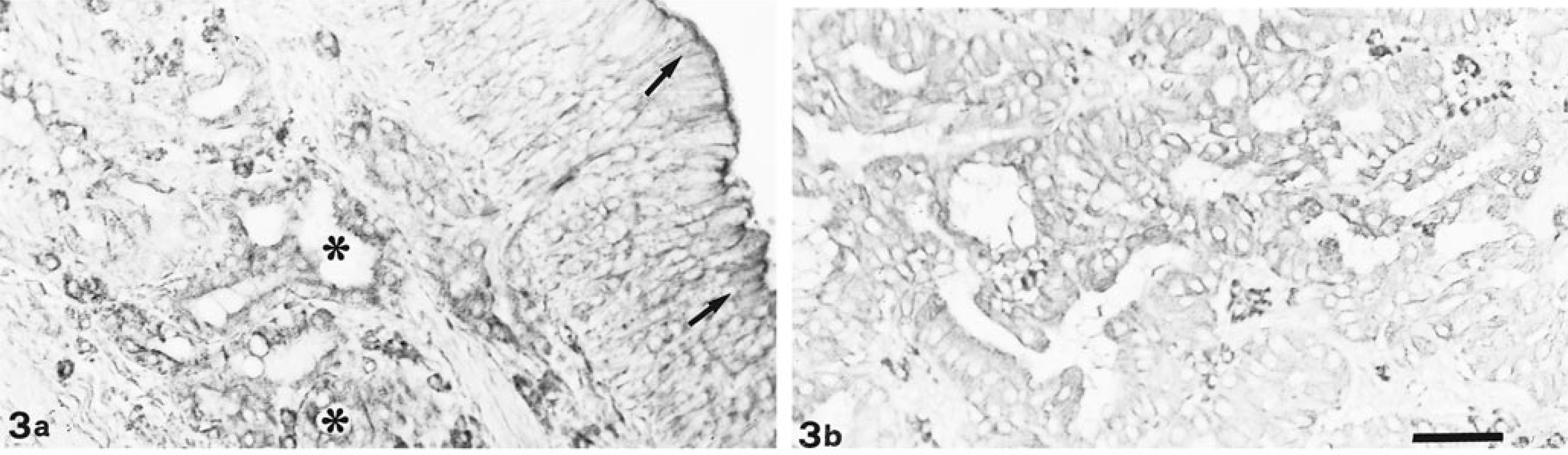
Con A staining. Both olfactory epithelium (arrows) and glands (∗) react strongly to Con A (a). Also tubular portion of EIT shows strong positivity (b). Bar = 33 µm.
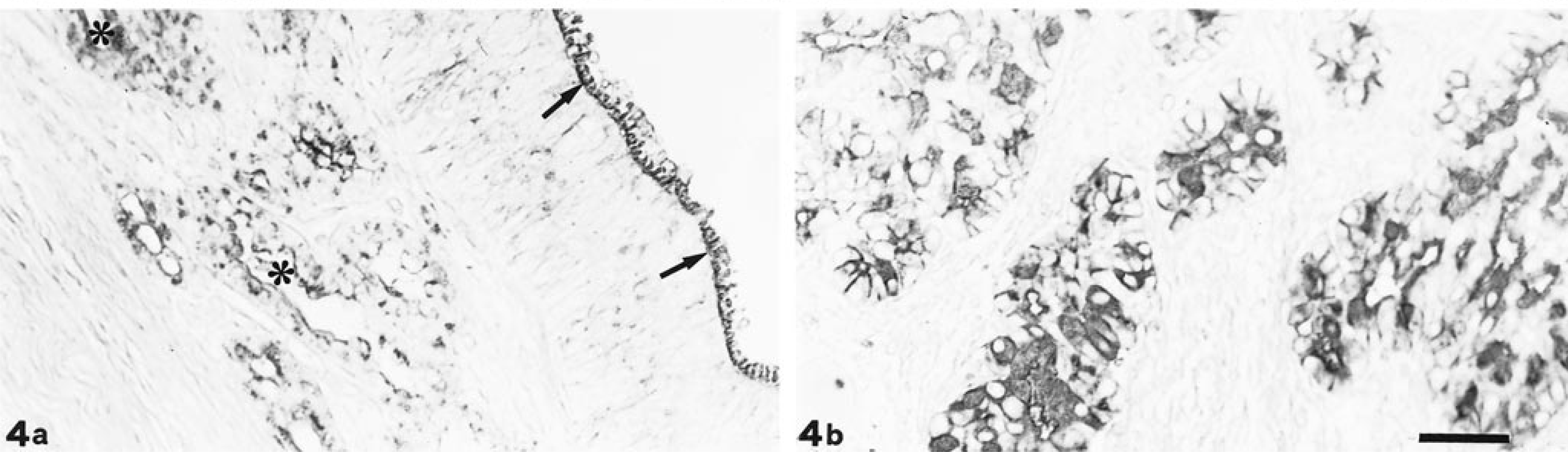
WGA staining. Olfactory glands (∗) react strongly to WGA; also, the cell coat of olfactory epithelium (arrow) appears positive (a). EIT tubular portion shows very strong staining (b). Bar = 33 µm.
LTA and UEA I showed overlapped binding sites, with the exception of goblet cells of the respiratory epithelium (Fig. 5). Fucosidase digestion did not affect any lectin reactivity. Reactivities to the histochemical treatments are summarized in Table 1.
Reactivity of samples to histochemical treatments.∗
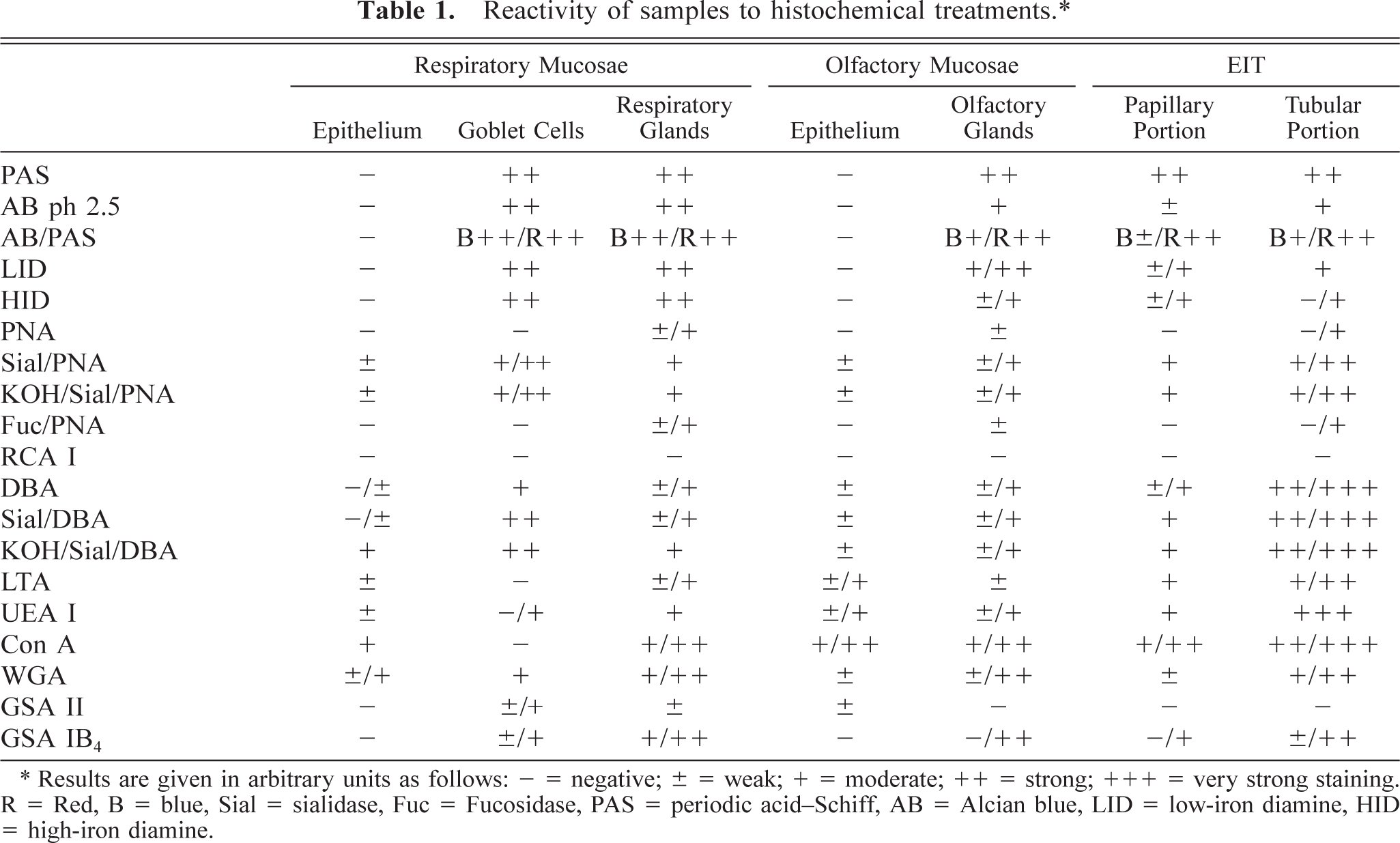
Results are given in arbitrary units as follows: - = negative; ± = weak; + = moderate; ++ = strong; +++ = very strong staining. R = Red, B = blue, Sial = sialidase, Fuc = Fucosidase, PAS = periodic acid-Schiff, AB = Alcian blue, LID = low-iron diamine, HID = high-iron diamine.
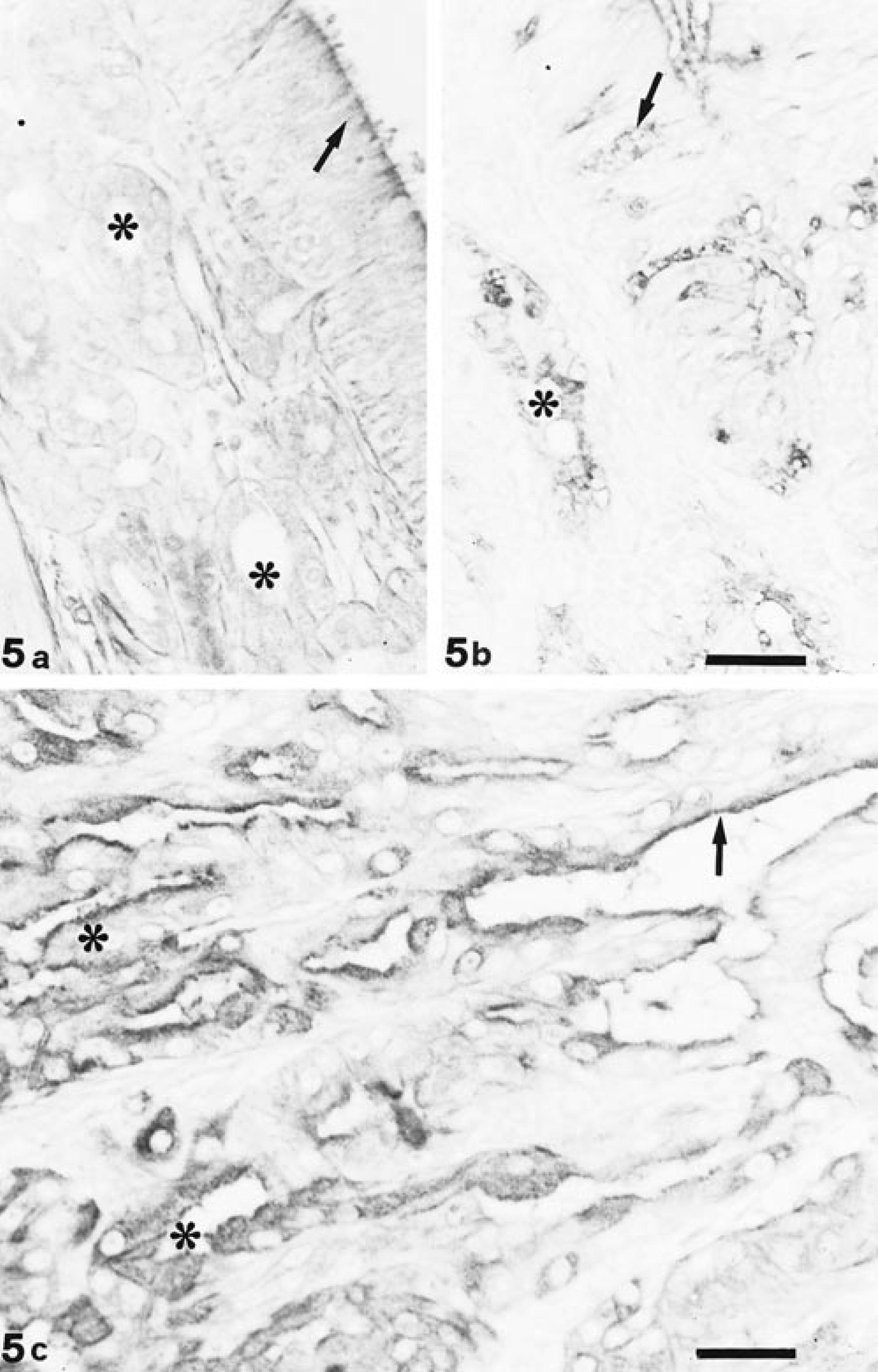
UEA I staining. Both epithelium (arrow) and olfactory glands (∗) react moderately to UEA I (a); respiratory mucosa shows positive sites at goblet cell (arrow) and gland (∗) level (b). Bar = 33 µm. Cells of tubular portion (∗) of EIT appear strongly reactive to UEA I, whereas in papillary portions, the reactivity is restricted to the luminal border (arrow; c). Bar = 25 µm.
Discussion
The aim of this work was to determine the histochemical characteristics of secretion products in the EIT and in normal respiratory and olfactory mucosae of goats in order to clarify the histogenesis of EIT. We expected to find some histochemical differences between normal and tumoral tissues as a consequence of both qualitative and quantitative modifications of cellular secretions caused by the neoplastic transformation.
The results of our conventional histochemical procedures led us to hypothesize a strong presence of neutral and prevalently sulphated acid and glycoconjugates (GCs) in the glands of the respiratory mucosa. We also hypothesized that the olfactory mucosa seems to produce neutral GCs and a very small amount of carboxylated GCs.
Cells of the papillary portion of EIT prevalently express neutral GCs and a smaller amount of sulphated GCs; the tubular portion of EIT synthesizes a great amount of neutral GCs and a small quantity of prevalently carboxylated acid GCs.
The two recognizable growth patterns in EIT are not only different morphological expressions but also reflect different histochemical behavior that could testify to a twofold histologic origin of the tumor.
In all the goats examined, the tubular portion of EIT produced neutral and prevalently carboxylated acid GCs like the glands of the normal olfactory mucosa. Staining intensity was almost the same in both tissues; the only difference concerned the distribution of stained products. Indeed, in the cells of the tubular portion, staining was widespread in the cytoplasm, whereas in the olfactory mucosa, only the apical cytoplasm of the glandular cells reacted. The papillary portion of EIT showed the presence of neutral and prevalently sulphated acid GCs, as occurred in the glands of normal respiratory mucosa, although to a lesser extent.
On the basis of the above considerations, we may speculate on the hypothesis of a nonunivocal histologic origin of EIT, as proposed by several authors. 4 23 The neoplastic process could start from the olfactory glands for the tubular portion of EIT and from the respiratory mucosa glands for the papillary one, suggesting the possibility of a double glandular origin for the two growth patterns of goat EIT. In addition, the constant histochemical negativity of the epithelial lining in the normal nasal mucosa seems to exclude its involvement in EIT histogenesis.
Lectin histochemistry confirms the strong presence of neutral glucidic residues in all the samples examined. In particular, there is a strict correspondence between the presence of α-N-acetylgalactosamine, α-
In the olfactory glands and the tubular portion of EIT, there is a correspondence between the presence of α-N-acetylglucosamine and α-galactose residues and the sialic acid–β-
Lectin histochemistry partially confirms the hypothesis that EIT originates from both respiratory and olfactory glands. However, because further evidence to strengthen this hypothesis on the histogenesis of EIT has emerged, the problem requires additional approaches.
Footnotes
Acknowledgements
We wish to thank Ms. Natalina Cammertoni for technical assistance. This study was supported by a University Grant for Research.