
Abstract
Marginal siderosis is recognized in humans as an uncommon clinicopathologic entity characterized by degeneration of neural tissue at the surface of the brain and spinal cord, in association with the accumulation of hemosiderin, and resulting from chronic subarachnoid hemorrhage. The sources of hemorrhage are various and include neoplasms, malformations, cysts, and vasculopathy. Marginal siderosis of the spinal cord due to a myxopapillary ependymoma was diagnosed in a 19-year-old Dutch Warm Blood horse with clinical signs of myelopathy. There is only one previous report of marginal siderosis in the veterinary literature, also in a horse with clinical myelopathy.
Marginal (superficial) siderosis is an uncommon but well-recognized clinical and neuropathologic entity in human medicine. 1 3 5–9, 11 12 13 The name derives from the diffuse accumulation of hemosiderin pigment at the surface of the brain and spinal cord, that extends a short distance into the substance of the neural tissue, where the hemosiderin is stored within glial cells. This process is accompanied by gliosis, degenerative changes in the affected neural tissue, and the development of clinical neurologic disease. The underlying pathologic process is chronic low-grade subarachnoid hemorrhage, and the condition has been reproduced experimentally in animal models. 6 9 10 Sources of hemorrhage are various and include neoplasms, 1 12 vascular malformations, meningeal cysts, meningeal cavities left following surgical removal of masses, 3 and cerebromeningeal vascular amyloidosis. 11
Marginal siderosis has been reported only once previously in the veterinary literature to our knowledge, in a horse with clinical myelopathy associated with an aneurysm of the basilar artery at the level of the medulla. 13 Degenerative changes affected the spinal cord and the brain, including the cerebellar cortex.
Here, we describe a second horse with a clinical myelopathy associated with an ependymoma arising from the fourth ventricle that was the presumptive source for chronic subarachnoid bleeding.
A 19-year-old castrated male Dutch Warm Blood horse had been “unsound in the pelvic limbs” since July 1988. In the spring of 1999, a clinical diagnosis of equine protozoal myelitis was made. Serology revealed elevated antibodies against Borrelia burgdorferi, and doxycyline treatment was instituted. The horse was donated to Cornell University in the late spring of 1999, and neurologic examination determined that abnormal pelvic limb gait appeared to be a combination of spinal cord disease in the T3–L3 region and fibrotic myopathy, especially in the right pelvic limb. A cerebrospinal fluid (CSF) sample taken 3 weeks prior to necropsy had a very yellow color, a protein content of 149 mg/dl, 3,800 red blood cells/ul, and 3 nucleated cells/μL, with no cytologic abnormalities evident. Because of high confidence in the sampling procedure, the numerous red blood cells and xanthochromia were considered indicative of subarachnoid hemorrhage.
The decision was made to euthanize the horse, and a full necropsy was performed. The surface of the entire brain and spinal cord appeared diffusely bronzed due to orange-yellow discoloration, which was most pronounced through the cervical and cranial thoracic sections of the spinal cord. There was a large soft red mass, approximately 5 × 3 × 2.5 cm, at the left side of the cerebellomedullary angle. Transverse sectioning revealed this mass to be entirely extramedullary and associated with the choroid plexus and lateral aperture of the fourth ventricle (Fig. 1).
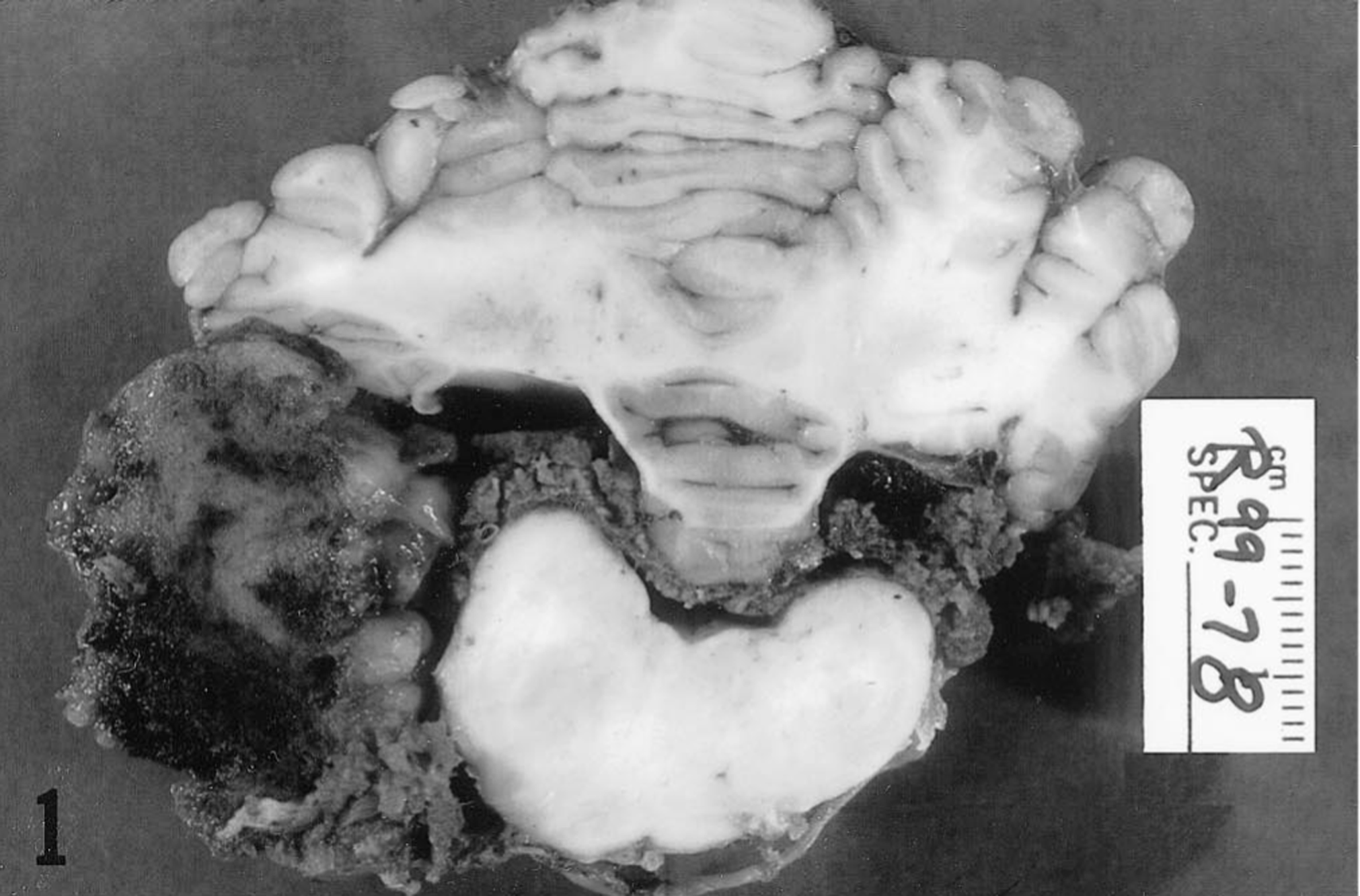
Caudal brain stem; horse. Cross section showing a large extramedullary mass of neoplastic tissue adjacent to the left lateral aperture of the fourth ventricle.
Tissues for histologic examination were fixed in 10% neutral buffered formalin and processed routinely for paraffin embedment. Sections stained with hematoxylin and eosin (HE) indicated that the mass in the caudal fossa was a neoplasm composed of fusiform cells with round to ovoid nuclei approximately 10–25 μm in diameter and containing coarsely granular chromatin. These cells had abundant but indistinct vacuolated eosinophilic cytoplasm and were frequently arranged in pallisade-like arrays, in which strands of cytoplasm extended towards thin-walled blood vessels (Fig. 2). Vessels were frequently cuffed by dense amorphous eosinophilic extracellular material (Fig. 3), which did not react as amyloid in sections stained with Congo red or crystal violet. Apart from these vessels, there was minimal stroma, and focal hemorrhage and hemosiderosis were prominent. Immunohistochemically, the tumor cells stained strongly for both vimentin (Fig. 3) and glial fibrillary acidic protein, which emphasized their slender fibrillar nature. The diagnosis was myxopapillary ependymoma, consistent with this variant of ependymoma as described in humans. 14
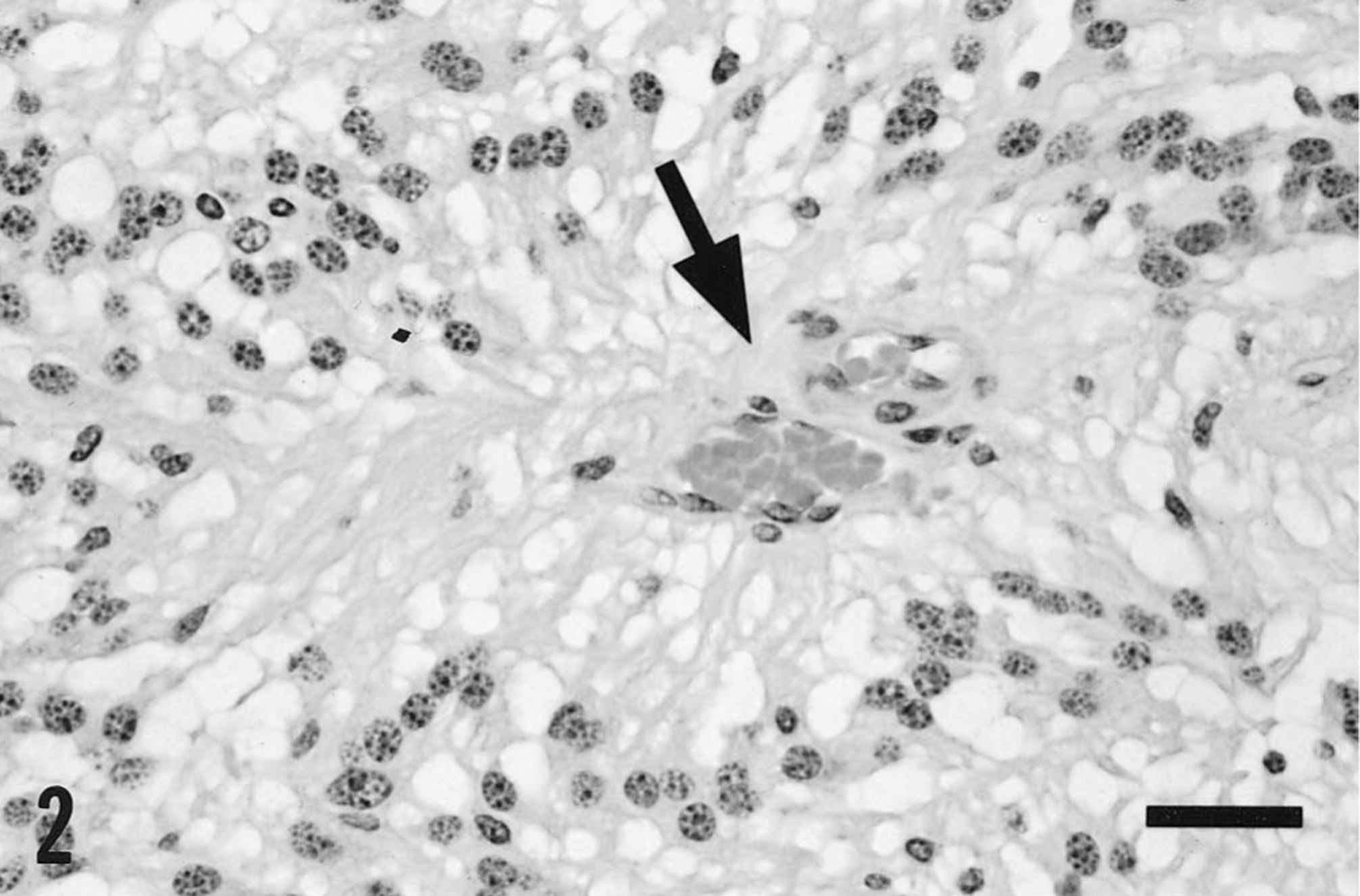
Mass from the caudal fossa; horse. Neoplastic tissue, showing pallisade arrangement of fusiform cells around a small blood vessel. The vacuolated and indistinct cytoplasm of the neoplastic cells “streams” from the perikaryon towards the blood vessel (arrow). HE. Bar = 30 μm.
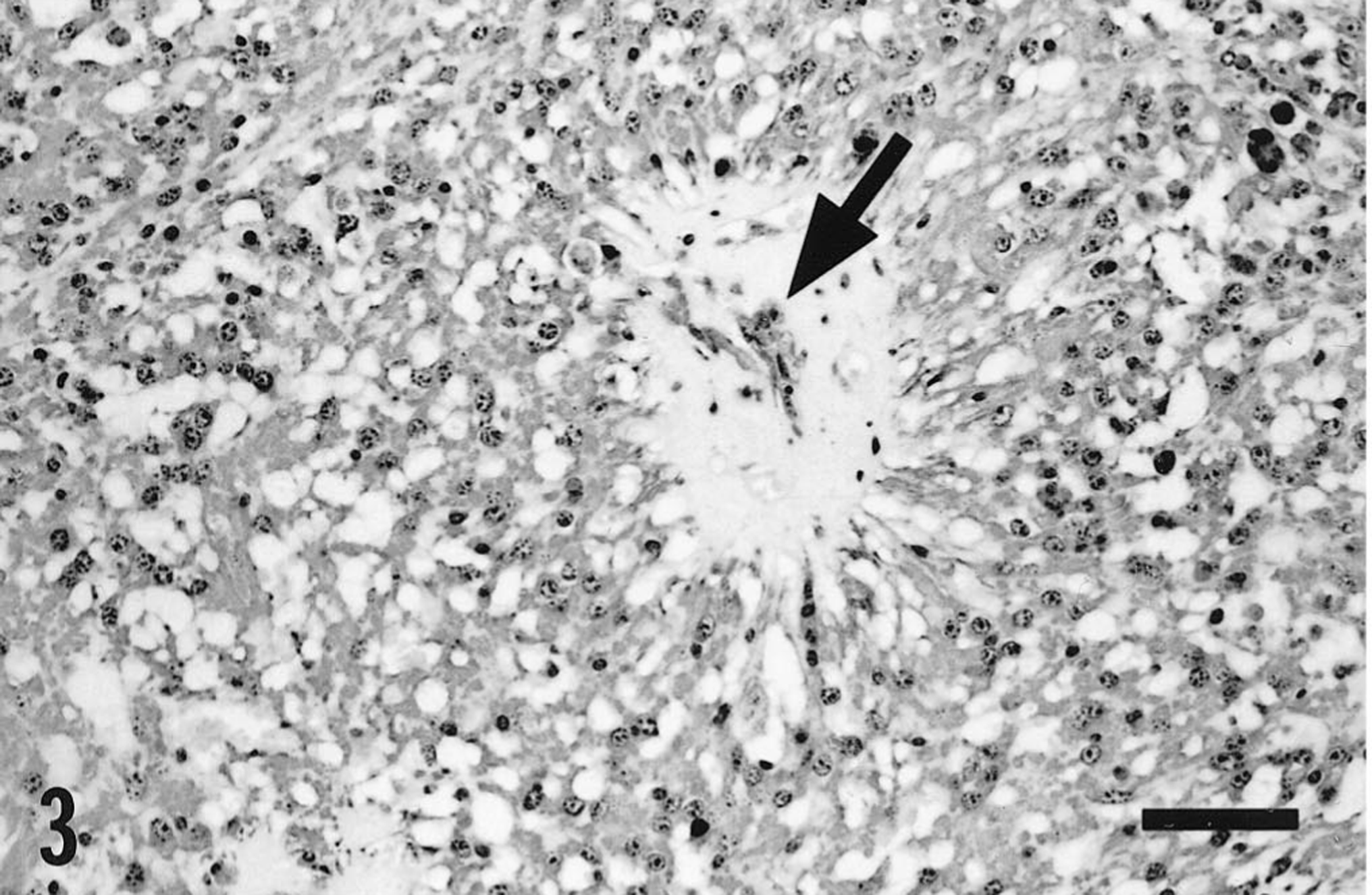
Mass from the caudal fossa; horse. Neoplastic tissue stained immunohistochemically for vimentin. There is strongly positive cytoplasmic staining of neoplastic cells. Also evident is a perivascular amorphous deposit (arrow), which was eosinophilic in sections stained with HE but did not stain as amyloid. Avidin biotin peroxidase complex method, Mayer's hematoxylin counterstain. Bar = 125 μm.
Histologically, the bronze surface discoloration of the brain and spinal cord was due to leptomeningeal and superficial parenchymal hemosiderosis, evident in routine HE-stained sections as brown intracellular pigment granules but appearing blue in sections stained by Perl's Prussian blue technique. In the leptomeninges, the pigment was present within macrophages, whereas in the nervous tissue it appeared largely within microglial cells and astrocytes (Fig. 4).

Margin of the cervical spinal cord; horse. Longitudional section shows hemosiderin storage (dark stain) in macrophages in the leptomeninges, and in both macrophages and glial cells within the neural tissue. The intensity of storage decreases away from the surface. Perl's Prussian blue. Bar = 125 μm.
In the brain, this pigmentation was not accompanied by any other histologic abnormality. By contrast, throughout all segments of the spinal cord, there were distinct degenerative and reactive changes involving tracts at the surface of the white matter in the dorsal, lateral (Fig. 5), and ventral funiculi but not adjacent to the ventral median fissure. Perl's staining revealed iron-containing glial cells in association with degenerative changes but not in deeper tissue in which degenerative change gave way to histologic normality (Fig. 4).
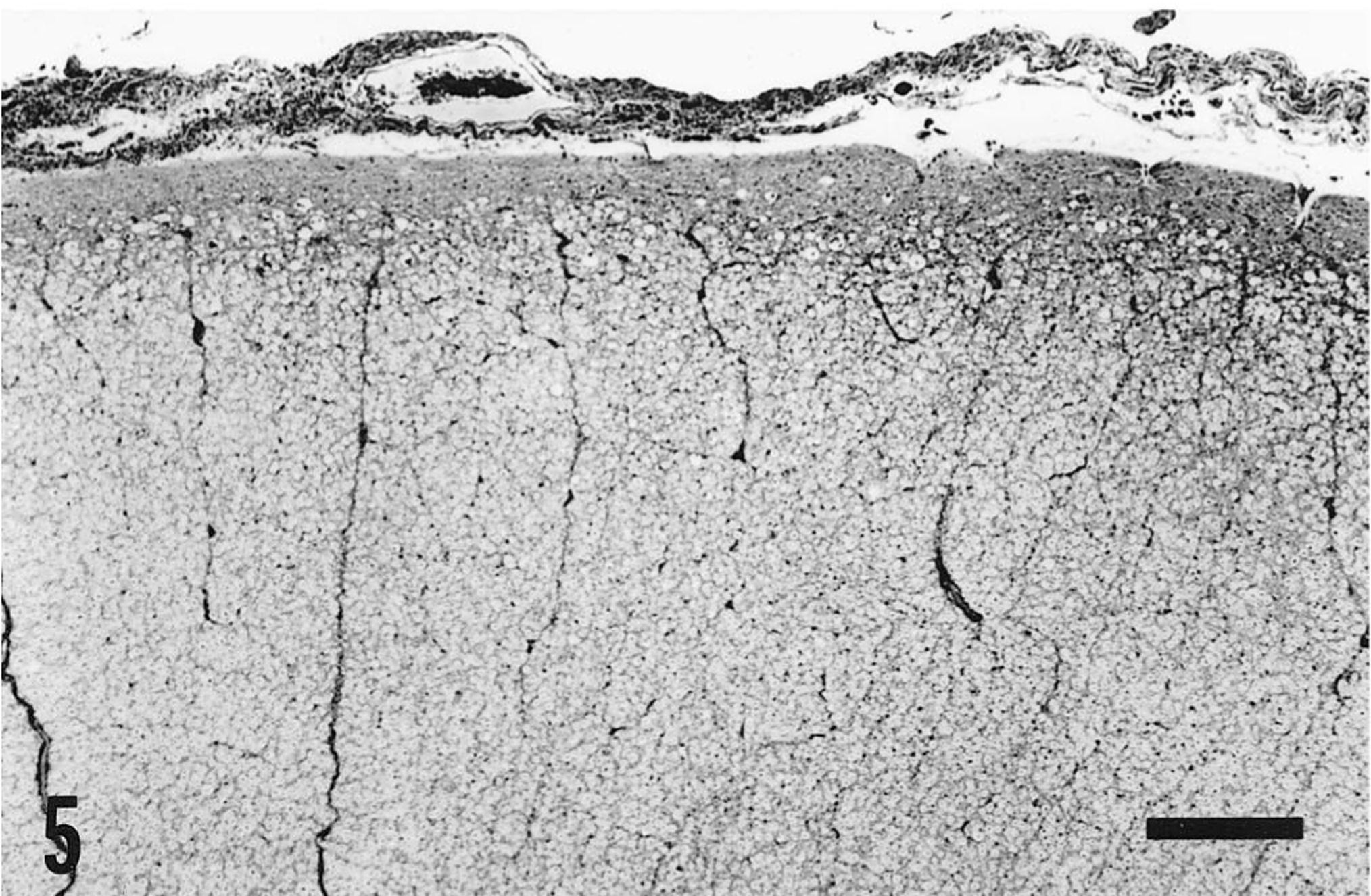
Cervical spinal cord; horse. Cross section showing a portion of a lateral funiculus. A superficial zone of gliosis and axonal degeneration is evident as an irregularity in the inner aspect of the glia limitans, which is slightly thickened, and as vacuolation of the neural tissue. This surface change was present to the same extent around the entire margin of the spinal cord with the exception of the surfaces adjacent to the ventral median fissure. Bielschowski. Bar = 400 μm.
The degenerative changes were manifest as swollen and fragmenting axonal segments, ballooning and loss of myelin, and fibrillary astrogliosis and were interpreted as ongoing axonal degeneration of some standing, with secondary myelin loss (Fig. 6). The combination of pigmentation, superficial tract degeneration, and clinical signs of spinal cord disease were considered justification for a diagnosis of marginal siderosis of the spinal cord. There were no pathologic findings to support the clinical diagnoses of fibrotic myopathy or protozoal myelitis. No Sarcocystis neurona could be isolated from cultures of spinal cord tissue.

Cervical spinal cord; horse. Longitudinal section showing marginal axonal and myelin degeneration and gliosis. The presence of fragmented axonal segments and ballooned myelin sheaths indicates an ongoing process. Bielschowski Bar = 160 μm.
Hemosiderin storage per se is not considered a harmful process, but it does reflect heavy iron trafficking. In humans, superficial neural siderosis is associated with severe cerebellar ataxia, neurosensory hearing loss, and spastic paraparesis, reflecting degeneration of the cerebellar cortex, eighth cranial nerve, and spinal cord. 3 7 The pathogenesis remains obscure, but most red cells released into the CSF are destroyed locally and their heme is metabolized to iron-free derivatives by heme oxygenase, during which there is potential for tissue injury via iron-catalyzed lipid peroxidation. 8 Microglia offer some protection from this outcome because they contain the biochemical apparatus for the conversion of iron to hemosiderin via ferritin biosynthesis. This process is promoted by the inactivation of a ferritin repressor protein (FRP), which normally binds to ferritin mRNA. 8 In the human cerebellum, heme-triggered inactivation of FRP appears to involve an undefined interaction between astrocytes (Bergmann glia), probably the major source of FRP, and microglia, the major site of ferritin biosyntheis. 8 This interaction is somehow related to the particular vulnerability of the cerebellum to iron-related injury.
In this horse significant degenerative change was confined to the spinal cord despite the fact that siderosis was extensive in the brain and spinal cord, and the site of hemorrhage was adjacent to the cerebellum. These findings may represent interspecies regional differences in glial metabolism and free radical-mediated cell injury.
Reports of equine ependymomas are rare, although two ependymomas have recently been described, apparently arising from the ventral aspect of the third ventricle. 2 4 Both showed papillary arrangements of the neoplastic cells but lacked the cytoplasmic vacuolation seen in this horse. Neither was associated with marginal siderosis.
Footnotes
Acknowledgements
We thank Ms. Alexis Wenski-Roberts for the preparation of photographs.