
Abstract
Many laboratory rodents grind their food into crumbs that are discarded at the bottom of the cage (sometimes called orts). This can have substantial impacts on measures of food intake and assimilation efficiency. We quantified food grinding in two laboratory mouse strains on eight different diets and distinguished between two hypotheses of why food grinding occurs: a stereotypic behaviour due to a lack of environmental enrichment, or part of an optimal food intake strategy. Orts were quantified when mice were exposed to environmental enrichment and when offered diets of differing energetic quality. Grinding was significantly different between diets, but not between strains, although there was a significant diet by strain interaction. Ort production was lowest on the hardest diets. Not accounting for orts could affect food intake estimates by up to 31.8% and assimilation efficiency by up to 16.7%. Environmental enrichment increased physical activity, but did not reduce grinding. Mice selected the higher energy density components of the food. We suggest a refinement of the current methodology for measuring food intake is essential, primarily because failure to take ort production into account created inaccurate estimates of food intake and assimilation efficiency in mice. Adding environmental enrichment is unlikely to reduce food grinding, but careful choice of diet will reduce the errors.
Laboratory mice are usually offered food in a hopper so they must gnaw at food pellets through metal bars. Food is offered this way to reduce contamination by faeces and urine and for easy measurement of food intake; calculated as the mass of food missing from the hopper each day. As mice feed, large chunks of food occasionally fall through the bars, which are usually put back into the hopper by researchers prior to weighing the food to estimate intake. 1 However, some mice also ‘grind’ food, resulting in a fine spillage discarded on the floor of the cage, sometimes called ‘orts’. 2 This term has been defined in the literature in different ways 3–10 and in many cases the exact definition is not stated. Both captive 11–14 and wild rodents 10,15,16 have been reported to grind food.
Koteja
Two separate hypotheses for the behaviour have been suggested. First, food grinding in captivity may be a stereotypic or compulsive behaviour 9 resulting from a lack of environmental enrichment. Normal behaviour patterns shown in wild mice, such as exploring and hiding may be lost in laboratory mice with no environmental enrichment. 23 This hypothesis predicts that grinding should be significantly reduced by providing mice with enrichment. Also, grinding should be independent of the quality and availability of the diet. Alternatively, optimal foraging theory predicts that ‘foragers should select a subset from the set of potential food items, to maximize net energy intake per unit time spent foraging’. 15 For example, wild animals often discard the non-digestible parts of the food (such as seed coatings) to select food of the highest energy density. 10,13,15 For mice, it may be beneficial to grind food to extract a more rewarding food component to maximize energy intake. 24 Therefore, as the heterogeneity of the diet increases and its quality decreases, it would be predicted that mice grind more to select the more energetically profitable parts of the food. Therefore, orts should have a lower energy content than the total diet as they should reflect poorer components. The impact of greater selectivity should generate a positive correlation between ort production and assimilation efficiency.
Koteja
Food grinding may impact on measurements of food intake if it is not adequately accounted for. We aim to evaluate the extent of the problem of ignoring ort production in food intake measures and to point to conditions under which orts may be particularly significant. In the context of ‘3Rs’, we aim to provide a refinement of the methodology for measuring food intake. Food grinding was quantified in two laboratory mouse strains: MF1, an outbred strain used extensively in studies of food intake during reproduction, 1,25,26 and C57BL/6J, an inbred strain used extensively, particularly in studies of obesity. 27 All mice were offered eight different commercially available pelleted rodent diets.
Animals, materials and methods
Experiment 1
Study animals were 16, six-month-old MF1 (HsdOla:MF1) mice. This strain was generated in 1970 from a cross of LACA and CS1 mice, maintained as a closed colony since 1970. Fifteen, three-month-old C57BL/6J (C57BL/6JOla:Hsd) mice were also used. Both strains were purchased from Harlan (Oxon, UK) and naïve to any other procedures. All procedures were licensed under the UK Animals (Scientific Procedures) Act of 1986 and received approval from the University of Aberdeen Ethical Review Committee. All mice were female and individually housed from birth, in cages of dimensions 48 cm × 15 cm × 13 cm (M3 base: North Kent Plastics, Kent, UK). Water was provided
Any large pieces of uneaten food found in the cage (that had fallen through the hopper bars) were returned to the hopper, and the mass of food missing from the hopper was recorded. Each day the paper bedding was shaken out and the contents of the cage separated by hand into faeces and orts. Orts and a sample of each diet were dried at 60°C (Gallenkamp, Loughborough, UK) for a minimum of 14 days. Food intake was calculated as the amount of food removed from the hopper, corrected for hydration and dried orts. Ort production was expressed as the mean amount fragmented (g/mouse/day). The percentage of the food fragmented (PF) was calculated as: (ort production/food intake) × 100%.
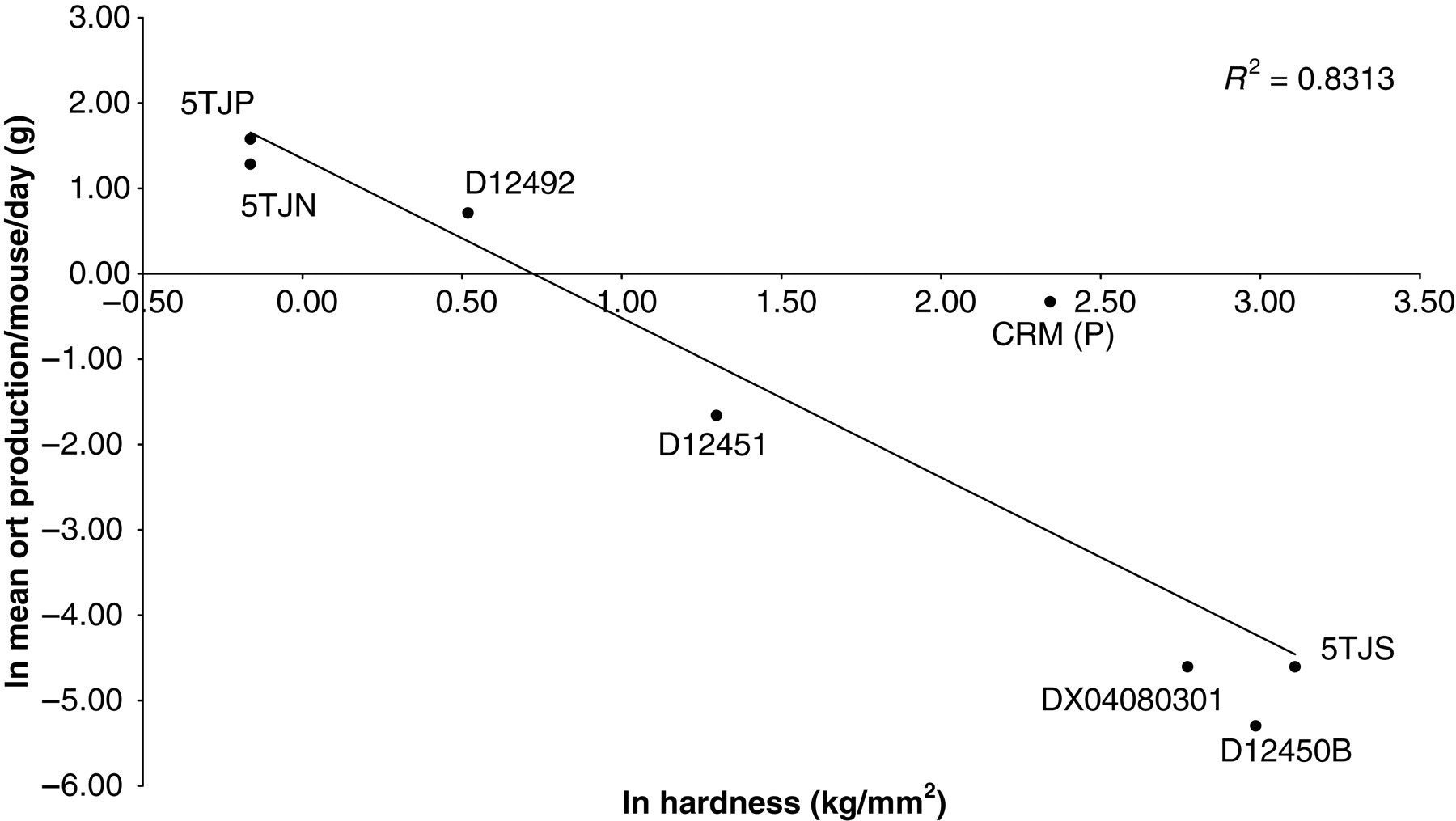
The hardness of each diet (kg/mm2) was measured using a digital microhardness tester fitted with a Vickers diamond indenter (Buehler, IL, USA). The hardness of each diet was tested from fresh material (as if at the point of feeding). Five replicates of each diet were taken and a mean hardness value calculated for each. Two diets (5TJP and 5TJN) were unsuitable for testing because they were too soft and easily crumbled by hand. The hardness of the softest measurable diet (D12492) was 1.7 kg/mm2. In the two cases where hardness could not be measured, it was set at the midpoint between 1.7 and 0 kg/mm2.
Experiment 2
Fifteen, 4.5-month-old female MF1 laboratory mice were used to explore the effects of diet quality, environmental enrichment and caloric restriction on food grinding. Housing and daily measurements were as detailed in experiment 1. Mice were sedated and implanted intraperitoneally with a wireless E-mitter (Model PDT-4000 E-Mitter, Mini-Mitter, Bend, OR, USA). Mice were sedated (isoflurane and oxygen mix) and maintained under anaesthesia throughout surgery via a nose tube. Surgical anaesthesia was assessed by checking the eyelid reflexes and limb movements. Mice were situated on a heat blanket throughout and breathing was continually monitored by observation. The E-mitter (size: 23 × 8 mm) was inserted intraperitoneally, and the wound sutured (Ethicon Vicryl; Dunlop's Ltd, Dumfries, UK). The surgical procedure took 20–30 min/mouse. Mice were administered with an analgesic by subcutaneous injection (Vetergesic [buprenorphine]; Dunlop's Ltd) and left to recover on a heat pad for up to one hour. Mice were put back in their cage and monitored for wound healing, but otherwise left alone for four weeks postsurgery. The implant recorded a series of activity counts each minute recorded by the Windows PC-based data acquisition system VitalView™ (Mini-Mitter).
Three independent experiments were carried out to determine if food grinding was affected by diet quality, environmental enrichment and caloric restriction. To test if ort production was affected by diet quality, mice were offered diets of three different energy densities consisting of standard pelleted rodent chow (Purina Mills chow #5001), with 0%, 20% or 40% cellulose added by weight (custom-made by Research Diets Inc). Diets were offered
Gross energy determinations and apparent energy assimilation efficiency
Faeces were collected from eight individuals during experiment 2 after at least five days on each diet
Statistics
Experiment 1
A one-way analysis of variance (ANOVA) was used to compare ort production, PF, food intake and body mass between each diet for each strain. Food intake, strain, individual and diet were all analysed in a general linear model (GLM) to determine which significantly influenced ort production and PF, including body mass as a covariate. Interactions between all these factors were considered.
Experiment 2
The effects of diet quality (0%, 20% or 40% cellulose) on ort production and PF were analysed using one-way repeated measures ANOVA. GLM was used to first examine if ort production was influenced by food intake or diet, including body mass as a covariate and second to determine if ort production differed within individuals on the different quality diets. To examine the effects of environmental enrichment, mean food intake, ort production, PF, body mass and activity were calculated for each mouse on three days: the day prior to, on the day of and the day following enrichment. One-way repeated measures ANOVA was used to find significant differences between days for each parameter. Data for the 0% and 40% cellulose diets were analysed separately. A paired Student's
Results
Experiment 1
Mean ort production, food intake, PF and body mass for mice from each strain on each diet are shown in Table 1. MF1 mice were on average 41% heavier than C57BL/6J mice ( Mean ort production for each mouse, on each diet in experiment 1. The MF1 strain is shown in (a) ( Ort production on each diet for mice from both strains combined in experiment 1 ( Average values of ort production, food intake, PF and body mass as measured in experiment 1 for PF: percentage of the food fragmented Values represent the mean ± standard deviation and significant differences are highlighted in bold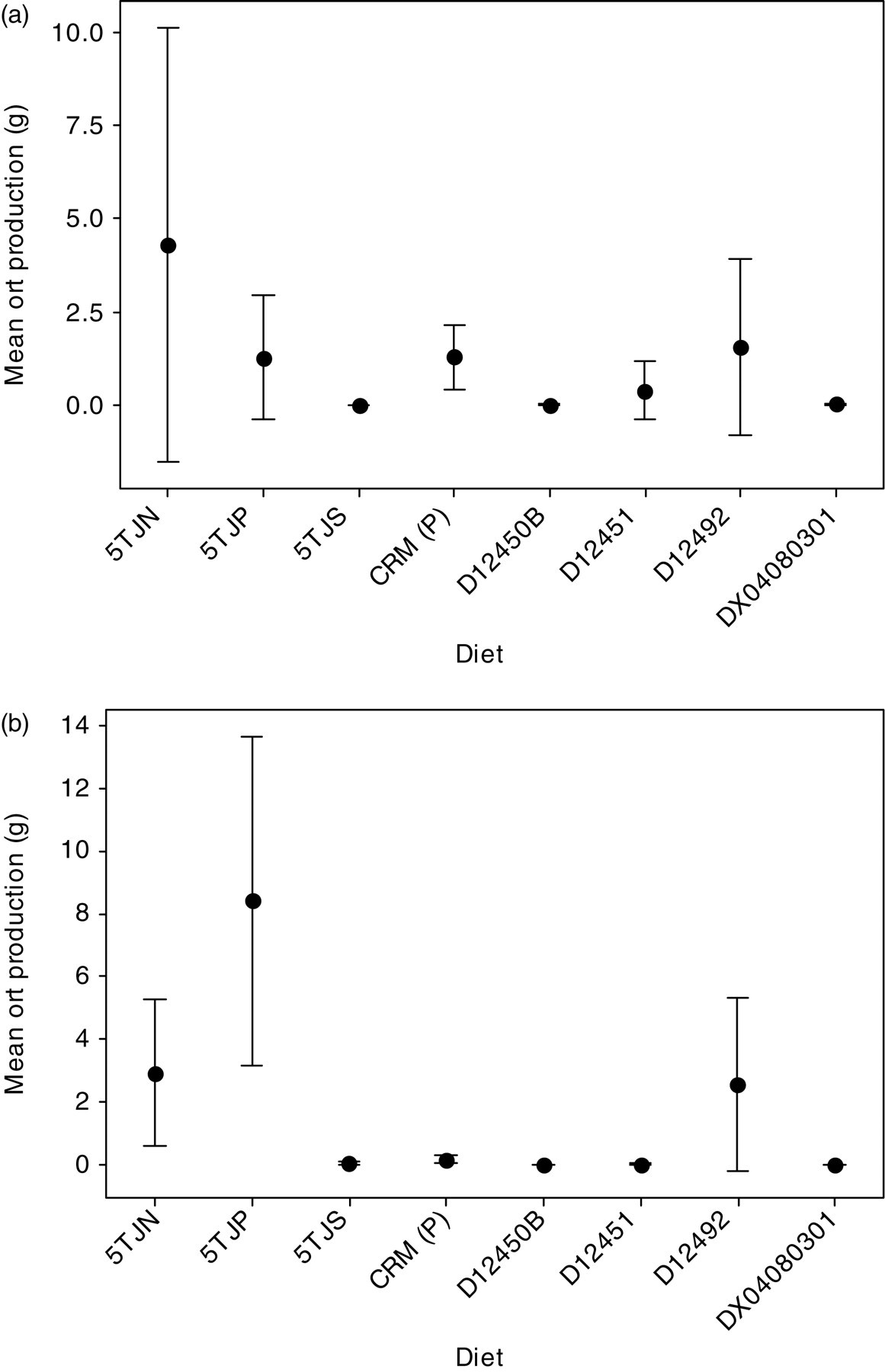
Experiment 2
During the diet quality experiments, ort production was not significantly related to body mass ( Mean ort production for each mouse (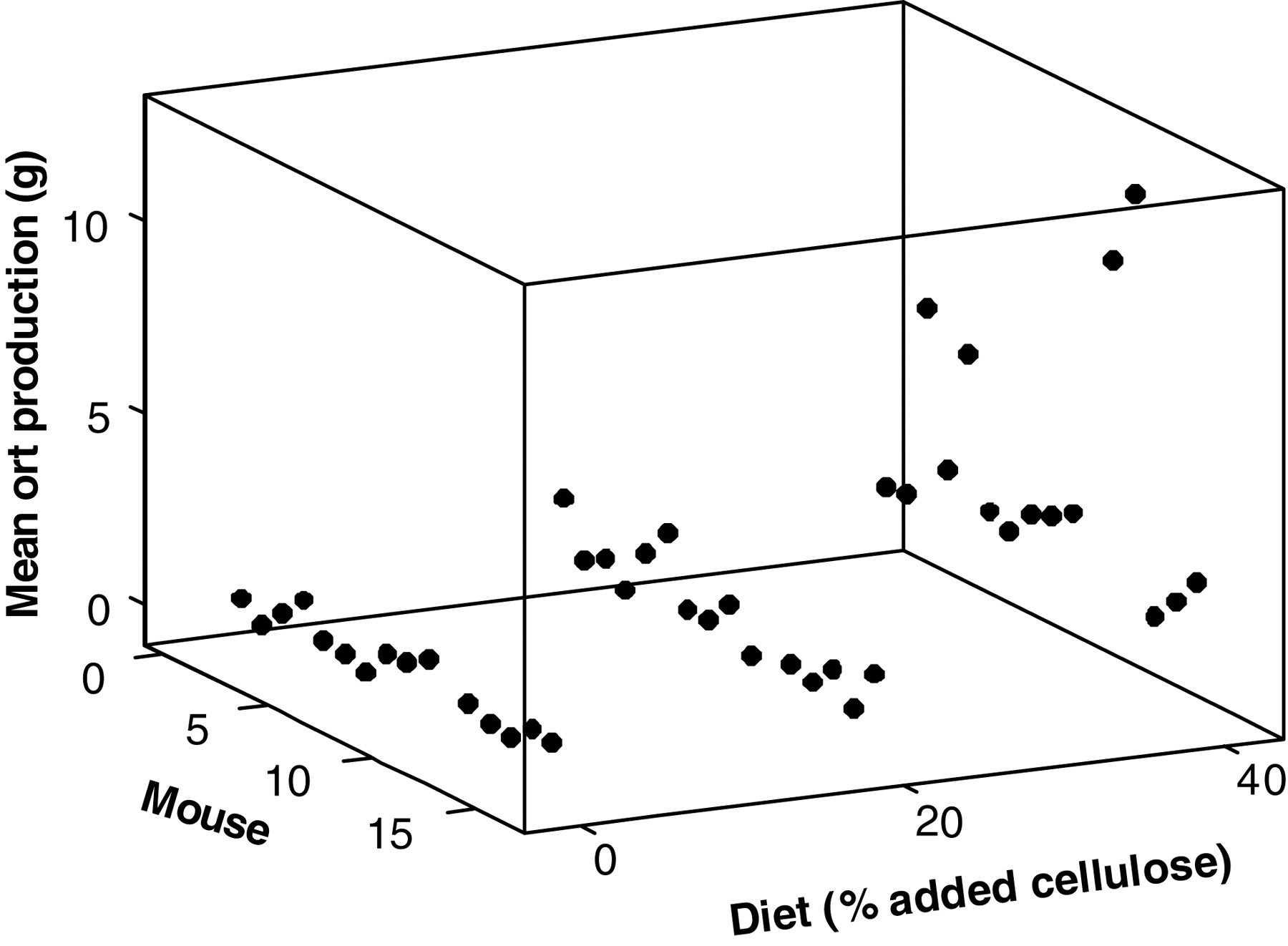
Data from experiment 2, from
PF: percentage of the food fragmented
Difference from the mean value of ort production, food intake, body mass, PF and activity were calculated on the day prior to, the day of and the day following addition of enrichment to the cages. One-way repeated analysis of variance was used to find significant differences between days for each parameter on each diet (0% and 40% cellulose). Values represent the mean ± standard deviation and significant differences are highlighted in bold
Average ort production on the 20% cellulose diet presented
GE contents and AEAE
Mean GE content of the faeces, orts, diets (0%, 20% and 40% cellulose) and cellulose from
GE: gross energy
Values represent the mean ± standard deviation and significant differences are highlighted in bold
Mean AEAE was 77.5 ± 9.4%, 68.9 ± 6.1% and 45.4 ± 8.8% for the 0%, 20% and 40% cellulose diets, respectively (
Discussion
The magnitude of error induced by failing to take orts into account when calculating food intake and AEAE can be large. The present study suggests that the effect varies with diet and with mouse strain, but errors in food intake can be as large as 31.8%, and assimilation efficiency estimates can be in error by up to 16.7%. Peterson and Wunder 13 found only five out of 30 studies accounted for orts in digestibility calculations. We also found that only five of 50 studies made reference to accounting for food grinding. 18–22 It is unclear whether this failure to mention grinding is because researchers do not recognize its significance and do not account for it. However, our data suggest that on some diets, mice produce significant levels of orts. We therefore suggest that a refinement in experimental designs involving the measurement of food intake and assimilation efficiency in mice is required (by collecting orts) to gain accurate estimates of each.
Grinding was directly proportional to the hardness of the diet, as also reported by Ford. 14 Rodent food manufacturers could strive to minimize the error in estimating food intake by making their pellets harder. Our data indicate that a target hardness of at least 2.5 kg/mm2 is desirable to minimize ort production. Currently, commercially available pellets containing high percentages of fat or fat from a number of different sources tended to fall below this critical hardness and were ground more. Failure to account for grinding may seriously compromise studies examining the impact of high-fat diet feeding on food intake. The Research Diets 45% kcal from fat diet (D12451) was the only ‘high-fat’ diet in our sample that combined a high level of fat with a hardness above this critical limit.
The difference between strains detected here supports the hypothesis that food grinding is heritable.
9
Recently, neurophysiological signalling pathways involved in grinding behaviour have been examined, further suggestive of a genetic component to the behaviour.
28,29
The fact that grinding may have a significant genetic component is particularly important when considering studies that concern genetically manipulated mice, as observed by Hastings
It has been suggested that grinding may be a stereotypic or compulsive behaviour because of a lack of environmental enrichment. 9 In our experiments, physical activity was stimulated when enrichment was added, indicating mice were responding to the presence of enrichment. However, this did not reduce the level of ort production, as was also reported in laboratory rats. 6 In fact, the presence of some plastic enrichment items (igloos) resulted in additional grinding of the plastic. The present data suggest that grinding is not due to a lack of environmental enrichment and that adding enrichment to cages as a strategy to reducing grinding behaviour in mice is unlikely to be successful. In our experiments, we only added enrichment for a single day and it might be argued that there would have been a more significant effect on grinding if it was in place for longer. However, this seems unlikely as the novelty aspect of enrichment would be lost over time and its capacity to distract the animals from grinding food probably also reduced.
Alternatively, grinding might be the consequence of an optimal food intake strategy. In support of this hypothesis, the energy content of the orts was lower than that of the food, suggesting mice were selecting the higher energy density parts of the pellets. Second, ort production fell to zero during caloric restriction when there was no advantage to be gained from being selective. Third, the extent of grinding increased as the cellulose content of the food increased indicating mice were being more selective of the best parts of the diet when there was more incentive to do so. However, the energy density of the diet was correlated with hardness and since in experiment 1 we showed that hardness was a key feature influencing the level of grinding, this effect may have been a coincidence. Moreover, if grinding the food served to eliminate the poorest parts of the food, the impact would be an increase in the energy content of the ingested portion of the diet. The impact of selecting out the ort fraction on the energy content of the ingested food was however very minor. Koteja
In conclusion, the present data suggest that food grinding by mice is not due to lack of environmental enrichment, but may reflect an optimization strategy. Grinding was dependent on diet hardness and was particularly high in high-fat diets. Moreover, the significant strain effect on levels of grinding supported the idea that grinding may have a genetic basis. Together, these effects mean that quantifying ort production is paramount in studies of different genotypes of mice and high-fat feeding. Failure to take ort production into account may cause serious errors in estimates of food intake and assimilation efficiency.
Footnotes
Acknowledgements
This work was partly funded by a studentship from the University of Aberdeen. Thanks to Yuko Gamo for help with ort collection. Thanks to Professor Steve Bull, Head of the School of Chemical Engineering and Advanced Materials and Chris Aylott, Design Unit at Newcastle University for assistance with diet hardness measurements.