
Abstract
Monitoring of rodents for Pasteurellaceae infection may be carried out by the polymerase chain reaction (PCR). We tested which of 17 rodent Pasteurellaceae strains were detected by three PCR primer sets. By phylogenetic analysis, 12 strains were assigned to the Rodent cluster and five strains to other clusters, namely the Somnus cluster, Pasteurella sensu stricto, Actinobacillus sensu stricto, the Mannheimia and Rossii cluster. A primer set developed to detect biotype Heyl [Pasteurella] pneumotropica produced amplicons from three strains and appeared specific for this taxon. A primer set developed to detect biotype Jawetz [P.] pneumotropica produced amplicons from the [P.] pneumotropica type strain and two other strains within the Rodent cluster. A primer set as described by Bootz and his co-workers (Bootz F, Kirschnek S, Nicklas W, Wyss SK, Homberger FR. Detection of Pasteurellaceae in rodents by polymerase chain reaction analysis. Lab Anim Sci 1998;
Pasteurellaceae belong to the most frequently occurring opportunistic pathogenic bacteria in laboratory rodents. 1 FELASA recommends the monitoring of rodents for all Pasteurellaceae. 2 Pasteurellaceae infection in rodents can be monitored by culture, serology and the polymerase chain reaction (PCR). 3–8
Bergey's Manual of Systematic Bacteriology 9 subdivides the Pasteurellaceae into 21 phylogenetic clusters. Only five clusters are represented by named genera, namely Actinobacillus sensu stricto, Haemophilus sensu stricto, Pasteurella sensu stricto, Mannheimia and Lonepinella. The remaining 16 clusters contain a wide variety of hitherto unnamed species and strains. Among these are strains misclassified as Actinobacillus, Haemophilus or Pasteurella. Misclassified strains are by convention indicated by brackets around the genus (or the species) name, 10 examples being [A.] muris, [H.] influenzae-murium and [P.] pneumotropica.
Strains reported as [P.] pneumotropica are genetically diverse 11–16 and Pasteurellaceae from hamsters 17 and guineapigs belong to other taxa than those from mice and rats. 9 Different PCR primer sets used might, therefore, differ in their ability to detect Pasteurellaceae infection in rodents.
We tested which of 17 rodent Pasteurellaceae strains were detected by three PCR primer sets namely those developed for the detection of all Pasteurellaceae, 6 biotype Jawetz [P.] pneumotropica 3 and biotype Heyl [P.] pneumotropica. 12
Materials and methods
Polymerase chain reaction
Bacterial strains examined comprised 17 Pasteurellaceae (Table 1) from hamster (n = 4), gerbil (n = 1), guineapig (n = 3), mastomys (n = 2), mouse (n = 5) and rat (n = 2) and 14 non-Pasteurellaceae namely Bacteroides distasonis, Bordetella bronchiseptica, Clostridium perfringens, Corynebacterium kutscheri, Escherichia coli, Helicobacter hepaticus, Klebsiella pneumoniae, Pseudomonas aeruginosa, Salmonella sp., Staphylococcus aureus, Streptobacillus moniliformis, Streptococcus faecalis, Streptococcus zooepidemicus and Yersinia enterocolitica.
*Species designations as supplied with the bacterial strain. A.: Actinobacillus; H.: Haemophilus; P.: Pasteurella. A bracket around the genus name denotes phylogenetic misclassification
†Strains were obtained from ATCC: American Typed Culture Collection; CCUG: Culture Collection University of Göteburg Sweden; DKFZ: Deutsches Krebsforschungs Zentrum, Heidelberg Germany; HIM: Hygiene Institut Philipps Universität Marburg, Germany; MHH: Medizinische Hochschule Hannover Germany; SSI: Statens Serum Institut, Copenhagen, Denmark. Round brackets (..) indicate subculture by Section of Laboratory Animal Microbiology (LAM), National Institute of Public Health, Bilthoven, The Netherlands
The Haemophilus strains were grown on chocolate agar (Oxoid, Haarlem, The Netherlands) and incubated for 18 h at 37°C under 7.5% CO2. All other bacterial strains were grown on 5% sheep blood agar (SBA) (Oxoid) incubated for 18 h at 37°C under 7.5% CO2, except Helicobacter hepaticus, which was incubated under microaerophilic conditions at 37°C for 48 h. Growth was harvested in 100 μL MQ (Sigma Chemical Co, St Louis, MO, USA) and heated for 10–15 min at 100°C in a heating block. Lysates were frozen at −86°C until testing.
Primers
DNA lysates of all bacteria were tested in duplicate using the primer sets described for biotype Jawetz [P.] pneumotropica by Wang et al., 3 Heyl [P.] pneumotropica by Kodjo et al. 12 and for Pasteurellaceae by Bootz et al., 6 which should yield PCR amplicons of 395, 937 and 515 bp, respectively, from Pasteurellaceae strains. All primers were synthesized by Gibco BRL Life Technologies (Breda, The Netherlands).
PCR protocol
DNA amplification was done in 25 μL reaction volumes. The PCR mix for the different primer sets was identical. Each sample consisted of 17.075 μL MilliQ, 2.5 μL 10X SuperTaq buffer (HT Biotechnology Ltd, Cambridge, UK), 0.125 μL TaqStart antibody (7 μmol/L, Clontech, Mountain View, USA), 0.3 μL SuperTaq (5 U/μL, HT Biotechnology Ltd), 1.5 μL MgCl2 (25 mmol/L), 0.5 μL dN(u)TP (10 mmol/L), 0.5 μL forward primer (10 pmol/L) and 0.5 μL reverse primer (10 pmol/L). Two microlitres of template DNA was added to the reaction mix. All PCRs were carried out in a GeneAmp PCR system 9700 (Applied Biosystems, Foster City, USA) under the conditions given in Table 2.
Amplicons were detected by performing electrophoresis of 10 μL of the reaction mixture in an ethidium bromide-stained 2% agarose gel (MP agarose; Roche, Basel, Switzerland). Marker IX (Boehringer, Mannheim, FRG) was used as the DNA size standard (72–1353 bp). DNA was visualized on an UV transilluminator and photographed.
Sensitivity studies
DNA lysate of rat [P.] pneumotropica study no. 9 was tested in a dilution series of 1 ng to 1 fg using Bootz's primers.
Measures to prevent DNA contamination
Preparation of PCR mixtures, addition of template DNA, and detection and analysis of PCR products were done in separate rooms to prevent intercurrent contamination of reaction mixtures by Pasteurellaceae DNA.
Statistical analysis
Differences in the number of Pasteurellaceae detected by the three primer sets were analysed using Fisher's exact test.
The level of significance was set at P < 0.05. Calculations were performed using the Kyplot statistical package, version 2.0 (KyensLab Inc, Tokyo, Japan) run on a PC.
Phylogenetic analysis
Part (about 407 nucleotides) of the gene coding for the 16S rRNA was sequenced for all Pasteurellaceae strains from the gerbil, hamster, mastomys, mouse and rat (Table 1).
The sequences were compared with sequences of reference bacteria from rodents, including the three guineapig Pasteurellaceae in our study (study nos. 2–4) deposited in Genbank (Table 1). A 16S rDNA dendrogram was constructed using the neighbour-joining method (multi-way option AlignPlus 5.0 Sci-Ed software (Durham, USA). Cluster names used are based on the Pasteurellaceae phylogeny as described in Bergey's Manual of Systematic Bacteriology. 9
Results
PCR sensitivity
The sensitivity of the PCR using Bootz's primer sets determined with serially diluted DNA lysates of rat [P.] pneumotropica study no. 9 varied from 1 to 10 pg.
PCR specificity
The primer set developed by Bootz generated a 515 bp amplicon from DNA lysates of all Pasteurellaceae strains (Table 1).
The primer set developed by Wang generated a 395 bp amplicon from three Pasteurellaceae strains comprising mouse [H.] influenzae-murium (study no. 5), a Jawetz-like [P.] pneumotropica strain from a gerbil (no. 11) and biotype Jawetz [P.] pneumotropica strain NCTC 8141T (no. 13).
The primer set developed by Kodjo generated a 937 bp amplicon from three Pasteurellaceae strains comprising two strains from mastomys (nos. 7 and 8) and mouse biotype Heyl [P.] pneumotropica strain ATCC 12555 (no. 12).
Bootz's primer set detected significantly more Pasteurellaceae strains than both the other primer sets (Fischer's exact test; P < 0.0001). There was no difference in the number of Pasteurellaceae strains detected by Wang's and Kodjo's primer sets.
None of the primer sets produced amplicons from the non-Pasteurellaceae strains tested.
Phylogenetic analysis
Figure 1 shows the genetic relationship of the 17 Pasteurellaceae strains with reference strains based on 16S rDNA sequences.
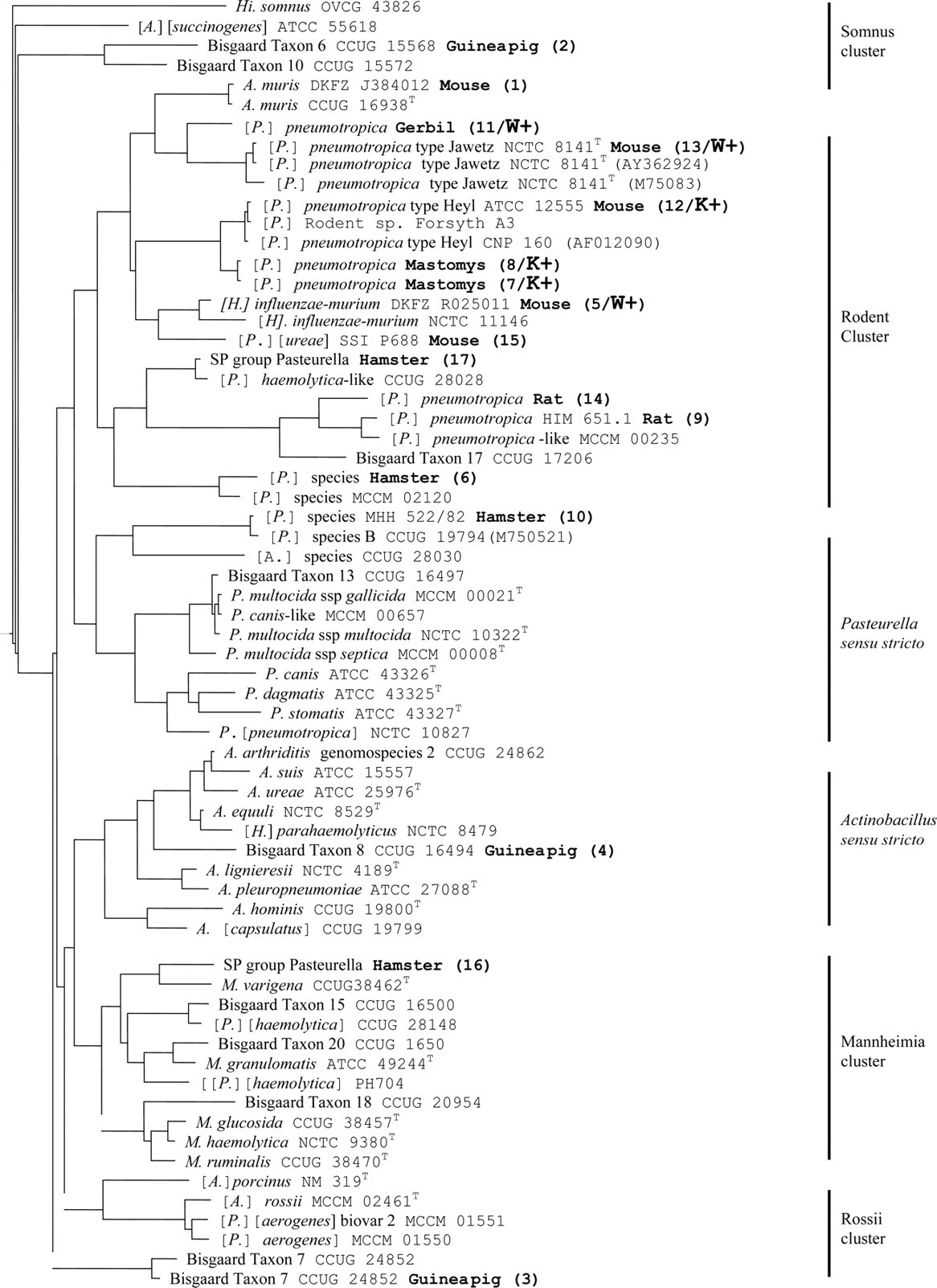
Unrooted neighbour-joining tree based on partial 16S rDNA gene sequences showing the phylogenetic relationship of rodent Pasteurellaceae and which strains were detected by primer sets developed for Jawetz biotype 3 and Heyl biotype [P.] pneumotropica. 12 The Pasteurellaceae phylogeny as described in Figure BXII.γ.215 (Bergey's Manual of Systematic Bacteriology, Vol. 2, Part B, pp. 854–5) 9 was used for cluster names and genus and species designations of bacterial strains. A.: Actinobacillus; H.: Haemophilus; Hi.: Histophilus; M.: Mannheimia; P.: Pasteurella and T: type strain. Current taxonomic name that needs revision between brackets, for example [P.]. Genbank accession numbers for 16S rDNA sequences of bacterial reference strains only included when not given by Olsen et al. 9 or where ≥2 sequences were available for comparison. In bold: host origin (study number/PCR outcome) using Wang's (W + ) or Kodjo's (K + ) primer set
Twelve strains were assigned to the Rodent cluster namely all strains from mastomys (n = 2), mouse (n = 5), gerbil (n = 1), rat (n = 2) and two of the four hamster Pasteurellaceae strains. The remaining two hamster strains were assigned to the genus Pasteurella sensu stricto and the Mannheimia cluster. The three guineapig strains belong to the Somnus cluster, the genus Actinobacillus sensu stricto and the Rossii cluster. 9
All Pasteurellaceae detected by Wang's and by Kodjo's primer sets were assigned to the Rodent cluster.
The three strains detected by Wang's primer set were in two subclusters of the Rodent cluster. The gerbil strain (no. 11) clustered with the Jawetz biotype [P.] pneumotropica type strain and [H.] influenzae-murium clustered with a mouse strain (no. 15) that was not detected by Wang's primer set.
The three strains detected by Kodjo's primer set clustered together around the Heyl biotype [P.] pneumotropica reference strain CNP160.
Discussion
FELASA recommends the monitoring of rodents for all Pasteurellaceae taxa. 2 The reasons for this recommendation reflect the unsettled taxonomic structure of the bacterial family, 9 the occurrence of taxa other than [P.] pneumotropica in rodents, 1 difficulties in proper classification of bacterial strains by commonly used methodology and the wide distribution of research complicating pathogenicity factors over Pasteurellaceae taxa. 18,19
Strains misclassified as [A.] muris, [H.] influenzae-murium and [P.] pneumotropica are assigned to the Rodent cluster. 9 Based on partial 16S rDNA sequencing, most of the Pasteurellaceae in our study comprising all strains from the mastomys, mouse, gerbil and rat could indeed be classed within the Rodent cluster.
The finding that some hamster Pasteurellaceae are genetically different from [P.] pneumotropica 17 may explain that two of our four strains from the hamster were outside the Rodent cluster (Figure 1). Other hamster Pasteurellaceae have been found to be genetically related to strains from the mouse and rat. 5,13
The separate position of guineapig Pasteurellaceae, which was already apparent from biochemical 20 and serological studies, 21 has been confirmed by phylogenetic analyses. 9 That in our 16S rDNA-based dendrogram the Bisgaard taxon 7 strain (study no. 4) does not seem to belong to the Rossii cluster (Figure 1) is likely due to the fact that we sequenced only part of the 16S rDNA gene of our strains. Based on full length sequencing of the gene, the strain is assigned to the Rossii cluster. 9
The fact that our 17 Pasteurellacaea strains that were in various clusters of the family were all detected by Bootz's primer set (Table 1 and Figure 1) confirms the probable suitability of this primer set to detect all Pasteurellaceae taxa claimed by the authors. In addition to the expected 515 bp amplicon, Bootz et al. 6 described a 1200 bp amplicon generated from [H.] influenzae-murium strains but the amplicon was absent in their earlier report on the same strains. 22 We did not observe a 1200 bp amplicon from [H.] influenzae-murium. Negative PCR results obtained with non-Pasteurellaceae confirmed what had to be expected from in silico evaluation of Bootz's primer set.
We also confirm the specificity of Kodjo's primer set for [P.] pneumotropica biotype Heyl (Table 1 and Figure 1, Rodent cluster). Biotype Heyl strains are genetically homogeneous and have less than 30% DNA–DNA hybridization homology with [P.] pneumotropica biotype Jawetz strains. 11 We did not find other studies in which Kodjo's primer set was used for PCR amplification of Pasteurellaceae.
Jawetz biotype [P.] pneumotropica strains are genetically heterogeneous and form a complex of various taxa at least at the species level. 11
The biotype Jawetz strain chosen for primer development might therefore be relevant to the ability of the set to detect other Pasteurellaceae.
Wang et al. 3 used the 16S rDNA sequence (genbank accession no. M75083) of strain ATCC 35149 that is identical to the [P.] pneumotropica type strain NCTC 8141T. Wang's primers indeed produced an amplicon of the expected size from this strain (Table 1 and Figure 1, Rodent cluster).
The heterogeneity of the [P.] pneumotropica biotype Jawetz complex might explain different findings using Wang's primer set in testing rodent Pasteurellaceae.
We obtained an amplicon of the expected size from [H.] influenzae-murium and a gerbil [P.] pneumotropica strain, whereas Weigler et al. 5 obtained the amplicon from Heyl biotype [P.] pneumotropica ATCC 13669 (not in our study).
Weigler also found the amplicon from [A.] muris strain Ackerman 80/443D (CCUG 16938T), which is highly related to the Wang PCR-negative [A.] muris strain from our study. Using Wang's primer set, Hayashimoto et al. 13 also found PCR-positive Pasteurellaceae strains that were highly related to PCR-negative strains. Although cross-reactivity of Wang's primer set with rodent Pasteurellaceae seems variable, it can be explained from in silico 16S rDNA sequence comparisons (data not shown).
Both Kodjo's and Wang's primer sets must be considered unsuitable to monitor rodents for Pasteurellaceae infection as they will detect only a fraction of the taxa from the bacterial family that have been cultured from rodents.
Predictably, Bootz's primer sets detected significantly more Pasteurellaceae than both the other sets (Table 1).
PCR testing might be supported by culture. The best sampling site for both PCR and culture remains the upper respiratory tract. Culture should be done when only few samples out of 10 (sample size recommended by FELASA) appear positive by PCR to exclude accidental laboratory error. If, however, 10/10 samples are found positive by PCR, it is less likely to be due to non-specific PCR reactions.
We conclude that the primer set developed by Bootz et al. 6 should be used to monitor rodents for Pasteurellaceae infection by PCR. Health monitoring reports should specify the primer set(s) used for PCR testing of rodents for Pasteurellaceae infection.