
Abstract
Climate change and technological advances are reshaping ecosystems and societies. Strategic choices that were best yesterday may be sub-optimal tomorrow; and environmental conditions that were once taken for granted may soon cease to exist. In dynamic settings, how people choose behavioral strategies has important consequences for environmental dynamics. Economic and evolutionary theories make similar predictions for strategic behavior in a static environment, even though one approach assumes perfect rationality and the other assumes no cognition whatsoever; but predictions differ in a dynamic environment. Here we explore a middle ground between economic rationality and evolutionary myopia. Starting from a population of myopic agents, we study the evolutionary viability of a new type that forms environmental forecasts when making strategic decisions. We show that forecasting types can have an advantage in changing environments, even when the act of forecasting is costly. Forecasting types can invade but rarely overtake the population, producing a stable coexistence with myopic types. Moreover, forecasting fosters collective intelligence by providing a public good which reduces the amplitude of environmental oscillations and often increases mean payoffs to forecasting and myopic types alike. We interpret our results for understanding the evolution of different modes of decision-making such as forecasting. And we discuss implications for the management of environmental systems of societal importance.
Keywords
Humans and non-humans face changing environments as they make decisions about their own behavior. Their actions influence one another, as well as the state of the environment, for example, by depleting resources or emitting pollution, which can feedback to change future strategic settings. We study how individuals make decisions in the context of environmental feedback—either as myopic agents, who consider only the present state of the environment, or as forecasters, who consider projections about future environmental states. We find that forecasting types can coexist with myopic types, dampen environmental oscillations, and increase the fitness of both types. These results highlight the potential for environmental forecasting to foster collective intelligence in groups, provide a theoretical explanation for multiple coexisting modes of decision-making, and have practical implications for sound stewardship of common-pool resources.Significance Statement
Introduction
Social-ecological systems are characterized by feedbacks that link ecological states and social decisions. These linkages can induce complex and cyclical dynamics in both human behavior and ecological states (Bieg et al., 2017; Levin et al., 2013; Tilman et al., 2018), driving the dynamics in systems of great societal importance. Models of social-ecological systems allow us to identify the key features that determine qualitative outcomes (Schlüter et al., 2012) and help us to guide management (Farahbakhsh et al., 2022). How humans make decisions over time can have important environmental implications. Individuals might deliberate and reason about their alternatives or follow heuristics and make rapid, automatic decisions (Evans, 2008; Kahneman, 2003, 2011). Different modes of decision-making have significant population-level consequences, and the interplay of automatic and deliberative thinking can drive cyclical dynamics in behavior and the environment (Rand et al., 2017; Tomlin et al., 2015).
The emerging field of eco-evolutionary game theory provides natural tools to model decision-making in social-ecological systems (Barfuss et al., 2020; Estrela et al., 2019; Lin and Weitz, 2019; Tilman et al., 2020; Weitz et al., 2016; Wang et al., 2020; Wang and Fu, 2020). Eco-evolutionary games arise when the strategies adopted by individuals influence the state of the environment, and the environment in turn influences game payoffs and strategic behavior, generating feedback. There is therefore a close relationship between models of social-ecological systems and the theory of eco-evolutionary games. In both cases, even simple systems can generate persistent oscillations with an ever-changing environment (Bieg et al., 2017; Tilman et al., 2020; Weitz et al., 2016).
Climate change is the result of decades of individual, corporate, and national decisions that have applied strong forcing to our climate system, such that an overshoot of the 2°C warming target is now likely (Pachauri et al., 2014). This overshoot is characteristic of the environmental oscillations seen in the theory of eco-evolutionary games (Menard et al., 2021; Tilman et al., 2020). The slow timescale at which the environment responds to our strategic behaviors sets the stage for this overshoot. Understanding this system as an eco-evolutionary game may provide important lessons for management and mitigation.
Cyclic dynamics also occur in fisheries, where harvester-driven collapse is ubiquitous (Essington et al., 2015; Pauly et al., 1998), which leads to reduced profitability of harvesting and sometimes eventual recovery of fish stocks (Hutchings, 2000; Worm et al., 2009). Understanding how modes of decision-making can aggravate or alleviate these human-driven environmental changes has profound practical implications. When should managers expect human behavior and decision-making to produce stable outcomes, as opposed to an unstable or oscillating environment (Bieg et al., 2017)?
Forest management can also be viewed as a social-ecological system that undergoes cyclic dynamics of fire, harvesting and regrowth (Luce et al., 2012; Steelman, 2016). In the United States, a decades-long management emphasis on fire suppression led to increased tree density in western forests (Fellows and Goulden, 2008), which then contributed to increasing wildfire risk (Marlon et al., 2012). This growing risk has been compounded by climate change, which manifests as increased drought frequency and severity, and greater peak summer temperatures (McKenzie et al., 2004). The USDA Forest Service has developed a strategy for confronting the wildfire crisis which relies heavily on prescribed fire and other fuels treatments (US Forest Service, 2022). However, shifting from a strategy of fire suppression to a strategy of deploying fire to generate fire-resilient landscapes poses major challenges given the current high fuel loads, climatic conditions, and increased extent of the wildland-urban interface (Radeloff et al., 2018). While the local patterns of wildfire are inherently stochastic, there may be predictable relationships at the landscape and regional scales between management strategies, forest ecosystem states, and wildfire risks. Here again, eco-evolutionary games may provide a useful tool to model these interactions and inform management.
More generally, long-term outcomes in diverse social-ecological systems depend on how individuals make decisions. The psychology of decision-making integrates experiences from the past, observations about the present, and expectations for the future (Zimbardo and Boyd, 1999). Time discounting measures individuals’ inter-temporal preferences for rewards and costs, and it has been measured empirically in humans and other animals (Mischel et al., 1989; Odum, 2011). Different discount rates imply different degrees of future orientation in thinking and decision-making. These differences in the weight placed on expectations for the future is known to impact environmental behaviors (Carmi, 2013; Carmi and Arnon, 2014; Enzler et al., 2019). Theoretical work has also highlighted the impact of foresight on cooperation (Perry and Gavrilets, 2020). And, yet, much of the work on social-ecological modeling has assumed strategies and behaviors are based on the current payoffs alone, disregarding projections for the future. In this paper, we will explore how individuals who forecast future environmental states might emerge in a population of myopic decision-makers, and how forecasters then alter environmental dynamics.
Economic theory and evolutionary theory both address the problem of decision-making in strategic settings. But economic and evolutionary analyses make vastly different assumptions about what information is available to individuals and how individuals use that information. In classical economic analyses, equilibrium strategies are identified such that fully informed and perfectly rational agents have no incentive to deviate unilaterally, that is, a Nash equilibrium (Nash, 1950). On the other hand, evolutionary game theory does not require individuals to know much, if anything, about the game they are playing (Maynard Smith, 1982) but, instead, considers individuals who make strategic decisions through a myopic search process. Such agents that lack cognition altogether nonetheless typically approach an equilibrium called an evolutionary stable strategy (ESS) (Maynard Smith, 1982). Remarkably, in static environments, these two concepts are tightly coupled: all ESS’s are Nash equilibria, and all strict Nash equilibria are ESS’s.
This remarkable concordance between the long-term behavior of fully rational individuals versus the behavior of simple myopic agents breaks down somewhat when considering equilibrium selection, which depends critically on the nature of the update process in evolutionary models (Golman and Page, 2010), yet is a topic about which the Nash equilibrium is agnostic. The divide deepens when strategic interactions take place in a changing environment, or when the strategies themselves generate environmental feedback. In a changing environment, a strategy that is favorable today may be detrimental tomorrow. In this setting, there is a wide gulf between the behaviors predicted by traditional game theory and bioeconomic theory (Clark, 2010) versus those that arise dynamically in an evolving population of myopic agents.
Here, we explore a middle ground between myopic agents and fully rational agents, that is, between evolutionary and economic theories of decision-making. The goal is not to determine which models of decision-making or strategic behavior are correct. Rather, we take the myopic setting as a starting point and ask when individuals will emerge who use more information about their environment and the game they are playing in making strategic decisions. That is, we study how more sophisticated modes of decision-making can arise in a population and what effects this has on environmental dynamics. In particular, we study the emergence of decision-making by individuals who forecast the future state of the environment and account for future payoffs based on their forecasts. Such individuals are not strictly myopic but also not fully rational or perfectly informed.
Our primary goal is to understand both the evolutionary viability of forecasting types as well as their resulting impact on strategic and environmental dynamics. Whereas a population composed entirely of myopic individuals can experience persistent cycles, we will show that one composed entirely of forecasters can produce a stable equilibrium. Moreover, the average fitness in the population of forecasters can be higher than it would be in the myopic population, even when forecasters pay a fixed cost, representing the cognitive or economic burden of making environmental forecasts. This notion has parallels to studies that consider a cost to complexity in decision-making (Rubinstein, 1986). We will show that forecasting types can invade a myopic population, but their invasion is self-limiting. Once forecasters stabilize the environment, then forecasting no longer has value (the future will be just like the present), and myopic individuals outperform the forecasters by avoiding the cost of forecasting. As a result, forecasters tend to increase when rare, dampening environmental cycles, but if they can stabilize the environment they cannot overtake a population.
We will also show that forecasters and myopic agents can stably persist together and that this coexistence can generate both public and private benefits. First, forecasting generates public benefit because even a small sub-population of forecasters serves to reduce the magnitude of environmental variability, which often increases the average fitness of both forecasting and myopic types when they coexist. Second, forecasting also generates private benefits because forecasters anticipate environmental change and are better able to deploy the right strategies at the right times. The private benefits allow the evolutionary emergence of forecasters, who then provide public benefits to all. Collective intelligence can be measured by the relative performance of individuals and groups (Leonard and Levin, 2022). The environmental stability and increased average payoffs that forecasting types often provide can therefore be seen as a form of collective intelligence.
Model
A broad class of eco-evolutionary games produces cyclic dynamics (Tilman et al., 2020; Weitz et al., 2016). In most models, the cycles are driven by actions of myopic agents. Taking this setting as a starting point, we model the emergence of decision-makers who use environmental forecasting in a resident population of myopic agents. Myopic types update their strategy following standard replicator dynamics. They switch between strategies based on the current payoffs they experience. In turn, their choices also alter the state of the environment, generating a feedback. But myopic types do not know anything about the underlying environment; they attend only to the strategies that others employ and the resulting payoffs they receive.
In contrast to myopic types, forecasting types know and collect more information when making their strategic choices. Forecasting types make forecasts of the future states of the environment, and they account for how the changing environment will influence their future payoffs, discounting the expected future payoffs in combination with present value of each alternative strategy. We also model this with replicator dynamics.
To model the emergence of forecasting, we must also consider the process by which forecasting and myopic types compete. We assume that the act of forecasting carries a cognitive or economic cost. While forecasters may believe that their current strategy will yield large payoffs in the future, this does not give them an advantage over myopic types in the present. Thus, we assume that forecasting and myopic types compete with each other based solely on their instantaneous fitness. Figure 1 illustrates these different types of updates.
With these assumptions, the deck is stacked against forecasters. How could a population of forecasters emerge when each forecaster is at an inherent fitness disadvantage and must compete with myopic types based only on instantaneous payoffs? We find that, nonetheless, environmental forecasting allows individuals to anticipate when environmental change will cause a different strategy to be favored, leading to instantaneous advantages that allow forecasters to invade. Individuals interact in a changing environment, represented here by the level of the environmental resource shown in green. At any point in time, a focal individual may update their strategy or their type. When a forecasting type (represented by a triangle) updates their strategy, they account for expected future environmental changes. A focal individual may also switch types, from a forecaster to a myopic agent, or vice versa. The environment responds to the frequency of high- and low-impact strategies (shown in brown and cream, respectively) in the population. An individual’s type has no direct impact on environmental dynamics, but forecasting and myopic types often adopt different strategies and so environmental dynamics respond indirectly to the composition of forecasting and myopic types in the population. We study the emergence of decision-makers who use forecasting, and what effects they have on environmental dynamics. The game payoff matrix Π for two alternative strategies depends on the state of the environment, n(t), which changes over time in response to individuals’ actions. A myopic type individual chooses between two alternative strategies based only on the instantaneous payoff of each strategy s in the current environment, Π
s
(n (0)). Whereas a more complex type forms a forecast about the future environmental states, 

Eco-evolutionary games
We assume that agents are engaged in a two-strategy eco-evolutionary game, building on the framework developed by Tilman et al. (2020). The two strategies are called the “low-impact” L and “high-impact” H alternatives, to denote the magnitude of their effects on the environmental state. When all individuals follow the low-impact strategy, the environmental state tends toward its highest value. But when the high-impact strategy dominates in the population, the state of the environment declines toward its lowest possible value. The environmental state is described by a normalized quantity, n, that is bounded between zero and one. The payoffs for the eco-evolutionary game are assumed to be linear in the state of the environment, n, and in the frequencies of the low- and high-impact strategies. Therefore, the game can be represented by an environmentally dependent payoff matrix:

Allowing for both forecasting and myopic types of agents, there are in total four kinds of individuals in the population: forecasting or myopic types that follow either strategy L or strategy H. We use
The immediate payoffs to each strategy are
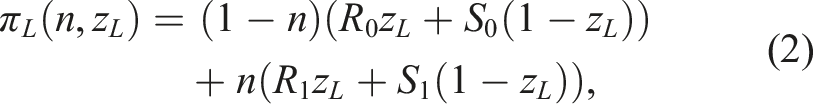
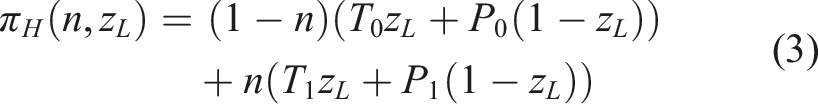
Myopic types consider only these immediate payoffs when updating their strategies. In contrast, forecasting types also account for their expectations of the future. These expectations integrate both environmental forecasting and payoff discounting.
Forecasting and discounting
When forecasting the future state of the environment, forecasting types form expectations about future payoffs. We assume that forecasters observe the current rate of environmental change and use linear extrapolation to project future environmental states,

A forecasting type must place some value on their future payoffs for their forecast to have any effect on their behavior. We assume that forecasting individuals put greater weight on their near-term payoffs than long-term payoffs. We model this through a normalized discount function of the form

Eco-evolutionary dynamics
We model strategic and environmental change based on pairwise interactions among the four possible kinds of individuals in the population. We assume that the act of forecasting is inherently costly.
For strategy change among forecasting types, discounting and forecasting are integrated into their assessment of the expected payoffs to the low- and high-impact strategies. This leads to the expected payoff of the low-impact strategy,
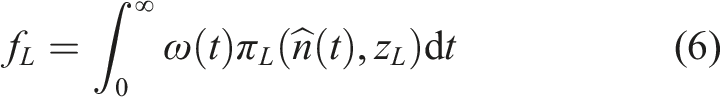

An equivalent expression holds for the high-impact strategy.
For myopic individuals, the immediate payoff difference drives strategy dynamics, that is

For forecasting individuals, the payoff difference between the strategies is

Now we develop dynamic equations that describe how the frequencies of all four kinds of individuals change in the population. First, we consider L-strategy myopic individuals. These individuals can emulate H-strategy myopic individuals, or they can choose to adopt either strategy of forecasting individuals. This leads to three terms in the rate of change of

The first term corresponds to replicator dynamics among myopic individuals. The second term corresponds to the flux of L-strategy forecasters into the myopic L-strategists caused by the cost of forecasting: all L-strategists receive the same immediate payoff from the game at any point in time, but those who are forecasters pay a fixed cost C for the forecasting ability. Herein lies the dilemma of forecasting.
The last term represents the flux of H-strategy forecasters to L-strategy myopic individuals, driven both by the (immediate) payoff advantage of playing strategy
Next, we consider L-strategy forecasting individuals. The dynamics of this sub-population of types follow
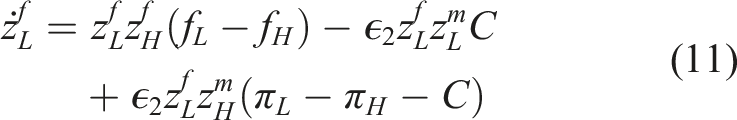
The first term reflects that forecasters switch between strategies L and H based on their perception of the net present value of each strategy, given forecasting and discounting. The second term reflects that forecasters are never favored over myopic types within strategy L or H, because they pay a cost to forecast, but the rate of switching types (myopic or forecasting) may be slower than the rate of switching strategies (L or H) if ϵ2 < 1. The last term shows that L-strategy forecasters transition to H-strategy myopic individuals according to the immediate payoff difference between the strategies and the cost of forecasting.
We can construct the other equations, for




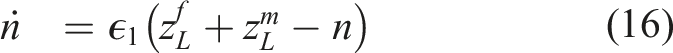
The equation for the environmental dynamic describes a decaying environmental variable (Tilman et al., 2020), which can correspond to, for example, pollution levels. In this case, the high- and low-impact strategies generate emissions of the resource. Tilman et al. (2020) show that systems with either decaying or renewing intrinsic environmental dynamics (corresponding, for example, a harvested population) generate qualitatively similar eco-evolutionary game dynamics. The timescale of environmental dynamics can differ from the timescales of strategy and type dynamics. Here, ϵ1 is the relative timescale of environmental dynamics, compared to strategy dynamics. In total, there are three timescales in the model. Without loss of generality, we consider relative timescales, with the timescale of strategy dynamics as the reference point. Thus, there are only two timescale parameters in the model. We summarize all model parameters and definitions in Table 1.
This formulation of the eco-evolutionary game rests on the assumption of mass-action kinetics, where there must be an encounter between any pair of types in order for a transition to occur between them. The likelihood of encounters is proportional to relative abundance, and so encounters among forecasters with different strategies will be rare when forecasting types are rare overall. We focus on the case where type switching is slow relative to strategy switching (ϵ2 < 1). We also assume that transitions between forecasting types and myopic types are based on the immediate payoff difference, not based on forecasters’ expectations of net present payoffs. In Supplementary Material Section 4, we present an individual-based micro-level model that converges to the system we study in the large population, weak selection limit.
Definitions of model notation.

Results
Myopic types alone
We focus our analysis on eco-evolutionary games in which a population of myopic individuals generates cyclical dynamics. These parameter regimes have been identified by prior work on eco-evolutionary games with myopic agents (Tilman et al., 2020). Figure 3 panel (a) shows the temporal dynamics in such a population of myopic individuals, which approaches a limit cycle where the state of the environment and the frequencies of high- and low-environmental impact strategies oscillate. These persistent cycles can directly reduce the long-term average fitness of the population compared to the fitness that could be achieved under a stable environment, while also increasing temporal payoff variability which would reduce the utility of risk-averse agents. There also exist cases where the cyclic dynamics driven by myopic types can increase average payoffs (see Supplementary Material, Section 3.3); however, we focus our analysis on regimes environmental stability increases payoffs, because this regime reflects the environmental dilemmas we seek to address. (a) Eco-evolutionary game dynamics in a population of myopic types, who can each assume either the high-impact strategy (h) or the low-impact strategy (L). The feedback between strategic choices and the environment creates persistent cycles, in which the L strategy is advantageous when the environmental state is high and the H strategy is advantageous when environmental state is low. (b) If all individuals are forecasting types who integrate environmental forecasts into their strategy updating, then the state of the environment and the strategic frequencies reach a stable equilibrium. (c) Forecasting types can co-exist with myopic types. After a lengthy transient (see Figure 4), both types co-exist in a stable limit cycle with reduced magnitudes of environmental and strategic variation compared to a population of myopic types alone. 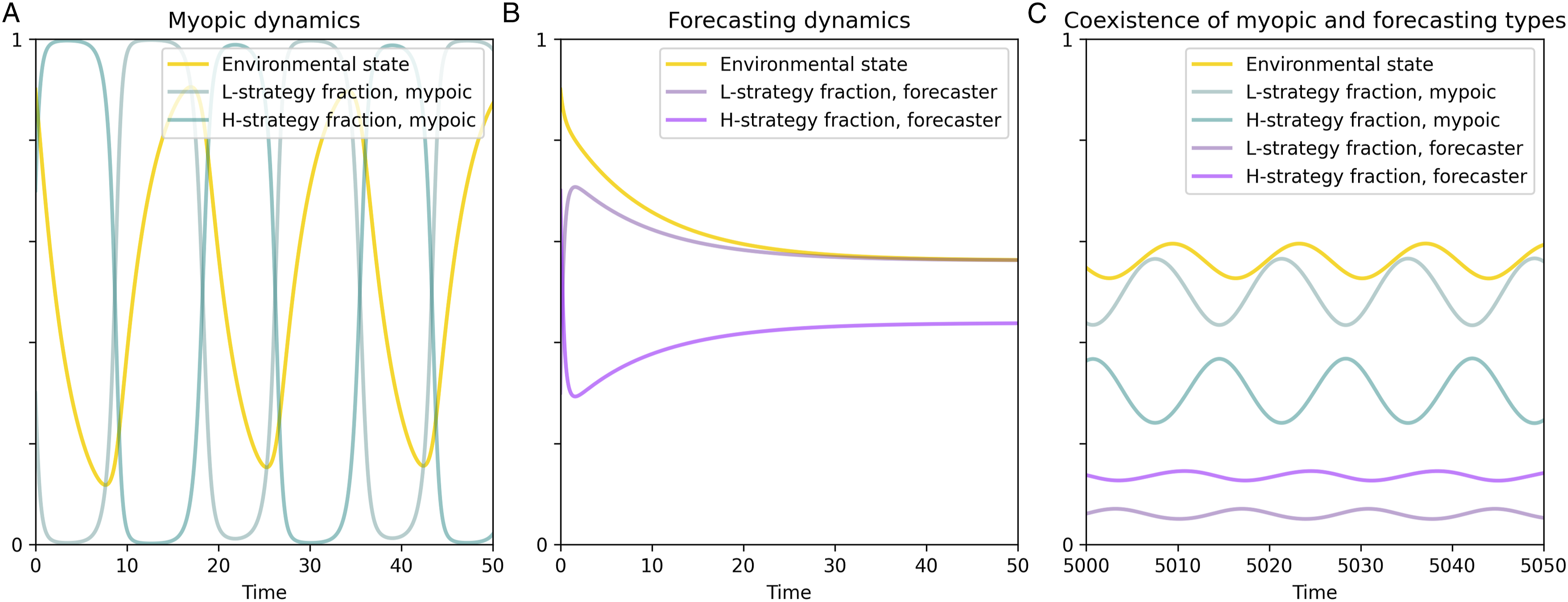
Forecasting types alone
In contrast to a myopic population, a population consisting entirely of forecasters generates a qualitatively different outcome. Provided forecasters care sufficiently about the future (i.e., have a sufficiently small discount rate), then forecasting types can produce a stable outcome for the same eco-evolutionary game that would exhibit cyclical dynamics in a myopic population. Figure 3 panel (b) illustrates a case where a population of forecasters produces a stable mixed equilibrium, with a fixed proportion of high- versus low-impact strategies and a fixed state of the environment. At this equilibrium, the average fitness attained by the population of forecasting types exceeds the average fitness that would arise in a population of myopic types. In other words, a population of pure forecasters can stabilize the eco-evolutionary system and increase average public welfare, compared to myopic decision-makers.
Coexistence of myopic and forecasting types
When forecasting types and myopic compete and interact, the coupled system does not approach equilibrium as quickly. Figure 3 panel (c) shows the long-run dynamics of forecasting types and myopic types who can each assume either a high-impact strategy or a low-impact strategy. Since forecasting is costly, it is not possible for forecasters to displace the myopic agents altogether—if they did, the environment would be completely stabilized, in which case the myopic agents make the same strategic choice as forecasters while avoiding the cost of forecasting. As a result, forecasting types and myopic types both persist, and so do environmental oscillations; but the amplitude of oscillations is diminished compared to what occurs in a population of myopic agents alone.
These results show that both cognitive types can coexist and that the presence of forecasting types, even when they comprise only a small fraction of the population, can have a significant impact on the dynamics of the system and the fitness of both cognitive types. Despite the persistence of some environmental oscillations when forecasting and myopic types coexist, the resulting population mean fitness over one environmental cycle can be greater than when only myopic types are present, and it can even transiently exceed the fitness when only forecasting types are present.
Invasion of forecasting types
We have shown that forecasters can coexist with myopic types and have a large effect on strategic and environmental dynamics. But the question remains: can the forecasting type invade a population initially composed of myopic types and reach a substantial frequency? To study the invasion of forecasting, we initialize simulations with only myopic individuals and let this system relax to its steady-state, which features a stable limit cycle with large oscillations. We then perturb the system by introducing forecasters at a low frequency and simulate that invasion process.
For any particular set of parameters, we find that the likelihood of a successful invasion is greater when the initial frequency of forecasting types is greater. The success of invasion also depends on the strategy composition among the invading forecasters as well as the exact timing of the introduction of forecasters within the environmental cycle.
Nonetheless, given sufficiently slow switching between forecaster and myopic types (ϵ2 ≪ 1), our simulations show that a successful invasion results in system-level behavior that always approaches the same limit cycle, regardless of invasion timing and strategy mix. The long-run dynamics after forecasters successfully invade and coexist with myopic types are relatively simple: both types persist in a stable limit cycle with a relatively small amplitude of oscillation (Figure 3(c)). But the invasion process itself is quite intricate.
The dynamics of a successful invasion occur at a much slower timescale than the environmental oscillations (Figure 4(a)), so that the invasion process takes many environmental cycles before the forecasters reach an appreciable relative abundance. Figure 4(b) shows the dynamics of fitness for forecasting and myopic types (averaged over one environmental cycle). The dashed lines represent the long-run average fitness attained by a population composed entirely of myopic individuals or by a population of pure forecasters. For these parameters, when only forecasting types are present, the environment equilibrates to a fixed-point and this leads to a greater long-run fitness. When only myopic types are present, long-run average fitness is lower, and the persistent environmental oscillations are substantial. Forecasters can invade under highly variable environments because forecasting allows them to foresee and predict the best time to switch strategies (L or H). This mitigates environmental oscillations and leads to a decrease in the magnitude of environmental variability. The reduction in environmental variability that forecasters provide acts as a rising tide that lifts all boats: when the environment is more stable, both forecasters and myopic individuals achieve payoffs as high as those that are attained when only forecasters are present. Since forecasting is individually costly and its environmental benefits are shared by all, forecasting is a public good. Despite serving as a public good, we see that forecasting can nonetheless emerge via evolution and increase the fitness of both myopic and forecasting individuals, even when forecasters comprise only a small portion of the population over the long term ( (a) Environmental forecasting is assumed to be costly, but forecasting types can nonetheless invade a resident population of myopic types and eventually reach coexistence, while reducing the amplitude of oscillations. As this example shows, a successful invasion of forecasters takes orders of magnitude longer than a single environmental cycle. (b) Dashed lines indicate the long-run fitness that would be attained by a population of purely myopic or purely forecasting types. Solid lines indicate the average fitness of forecasting and myopic sub-populations during the invasion process with both types present. The amplitude of environmental variability is reduced once forecasters reach appreciable frequency, which increases the fitness of forecasting and myopic types alike, eventually producing fitness as high as when only forecasters are present. Forecasting is thus a public good: it is individually costly and beneficial for all. While forecasters can invade and coexist with myopic types, a successful invasion is not assured and depends upon the timing of introduction. (a) In a purely myopic population, eco-evolutionary game dynamics converge to the outer black orbit and proceed in a counter-clockwise direction. We study the potential invasion of forecasting by introducing a small sub-population of forecasters at some point along this orbit. We assume that the forecasting sub-population has the same mix of strategies as the resident myopic population at the time of introduction. Points on the outer orbit indicate simulated introduction points that are evenly spaced in time. The color of the point illustrates whether or not the invasion of forecasters is successful from this point. The inner black orbit shows the long-run dynamics of the system for successful invasions, which are independent of the timing of the invasion. The purple and teal orbits illustrate the corresponding long-run dynamics among forecasting and myopic sub-populations. (b) Invasions progress very slowly, so that many environmental cycles pass before they approach the long-run dynamics illustrated in panel (a). The frequency of forecasters oscillates rapidly throughout this slow invasion process, and it may take a long time before it is clear whether the invasion will succeed or fail. 

We have focused on the region of the parameter space where cyclic dynamics under myopic types are well understood analytically. In Supplementary Material Section 3.4, we explore a different region of the parameter space where cycles can also arise under limited circumstances (Tilman et al., 2020), and we find qualitatively distinct invasion outcomes, including cases where forecasting types fail to invade a resident population of myopic types.
Successful and unsuccessful invasions
Whether or not forecasters will successfully invade and reach a stable frequency depends on initial conditions, in addition to game payoffs. In the simulations described above, we assume that when forecasters arise, they have the same mix of strategies as the resident myopic population. Nonetheless, the timing of the invasion within the environmental cycle and the initial frequency of invading forecasters determine whether the invasion will ultimately be successful and lead to coexistence.
Figure 5(a) shows invasion points that are evenly spaced in time, within a phase plane plot. The outer black orbit represents the long-run dynamics of a population of purely myopic individuals, with dynamics proceeding in a counter-clockwise direction. Whether or not a small frequency of forecasters can invade depends upon at what time point, within this periodic cycle, the forecasters are introduced: successful timings are shown as red dots and unsuccessful timings as brown dots. Even a successful invasion requires many environmental cycles to establish a stable frequency of forecasters (Figure 5(b)).
The inner black orbit in Figure 5(a) shows the long-run population-level dynamic after a successful invasion of forecasters. The magnitude of environmental and strategic variability is decreased substantially after invasion, which is reflected by a much smaller orbit in phase space.
Whether or not forecasters can successfully invade also depends on how much they value the present versus the future. Our simulations suggest that when forecasters place greater value on expected future payoffs (that is, a small discount rate r), this tends to increase their probability of successful invasion and establishment in a myopic resident population (Supplementary Material, Figure 3). Whereas if forecasters primarily value the present (large r), then they do not differ much in their strategic behavior compared to the myopic types, except they must pay a cost and so they cannot invade (Supplementary Material, Figure 3). And so, placing great value on the future always benefits forecasters, even if they end up putting more decision-making weight on predictions for the far-future that may prove false. These results make intuitive sense considering that forecasters types are continually updating their forecasts based on the current trajectory of environmental change.
Discussion
Strategic interactions often play out in changing environments. But if the environment is always changing, will individuals who account for environmental dynamics be favored when making strategic decisions? We find that in many circumstances environmental forecasters, who pay a cost for their forecasting ability, can indeed invade otherwise myopic populations engaged in eco-evolutionary games. Although a population of pure forecasters can produce a completely stable environment, forecasters cannot, in these cases, entirely over-take a population of myopic agents. Nonetheless, even when forecasters remain at low abundance, they have a striking effect on the dynamics of the eco-evolutionary game, greatly mitigating the amplitude of environmental variation and often increasing fitness for all.
Whereas classical bioeconomic approaches to decision-making assume that agents have complete information about the dynamics of the environment (Clark, 2010), evolutionary approaches often assume the opposite: individuals update their strategy based solely on present conditions (Safarzyńska and van Den Bergh, 2010). Here, we analyze a middle ground and ask whether agents who act slightly more like those studied in bioeconomic approaches can emerge in a strictly myopic population. That this invasion can be successful means that evolution by natural selection, or myopic imitation, can promote the emergence of more sophisticated cognition. A surprising and important caveat is that forecasters tend not to replace myopic individuals; instead, both types coexist.
The invasion of forecasters in an eco-evolutionary game can act as a rising tide that lifts all boats, increasing everyone’s fitness. In our simulations, forecasting behaves like a public good—providing benefits to forecasters and myopic types alike—and, thus, the invasion of forecasters can be thought of as a meta social dilemma, where both strategies (L or H) and decision-making types (myopic or forecasting) exhibit features of a social dilemma. When a myopic population experiences cyclic dynamics, the long-run average payoffs can be even lower than what would occur under the tragedy of the commons. This phenomenon has been referred to as an oscillating tragedy of the commons (Weitz et al., 2016), but given the damaging consequences for individuals’ average fitness and the risk that arises due to payoff variability, the “catastrophe of the commons” may be more apt. The invasion of forecasters resolves this catastrophe by ushering in greater environmental stability and increased fitness for forecasting and myopic types alike.
Rationality is a bedrock assumption that underlies much of microeconomic theory. However, the extent to which economic rationality applies to human decision-makers has long been questioned. Bounded rationality is an alternative model that acknowledges the limits of human cognition and the role of heuristics in decision-making (Gigerenzer and Selten, 2002; Simon, 1955), and it can arise as a result of evolutionary dynamics in stochastic settings (Brennan and Lo, 2012; Lo and Zhang, 2021). Similarly, prospect theory accounts for decision-making heuristics that violate classical notions of rationality and alter behavior (Tversky and Kahneman, 1974). These advances were central to the formation of behavioral economics, which explores how economic, social, and cultural factors affect decision-making. More recent work has considered the potential for multiple modes of decision-making to coexist in a population or within a single individual (Kahneman, 2011; Rand et al., 2017). Models starting from an evolutionary perspective have integrated foresight, where agents consider how changes in their behavior may alter the choices of others in the future (Perry et al., 2018; Perry and Gavrilets, 2020). However, little work has explicitly analyzed the consequences of feedback among modes of decision-making, dynamics of behavior, and the environmental setting, with a few notable exceptions (Adamson and Hilker, 2020; Austrup, 2020).
The focus of our paper has been on human-environmental systems, yet our modeling approach may possibly be applied to non-human organisms that live in variable environments. Flowering plants are most successful when they time their blossoming to align with favorable environmental conditions, such as temperature, pollinator abundance, and density of competitors. Accounting for environmental cues, including environmental change, may help plants achieve optimal flowering timing (Vermeulen, 2015). Migratory species face a similar dilemma about when to leave one habitat and head for another (Johansson and Jonzén, 2012); and some form of environmental forecasting may be advantageous in this context. Female Dusky Warblers modify their nest site choice in response to changes in the density of predators (Forstmeier and Weiss, 2004). This could generate feedback between the birds’ nesting strategies and the predation environment that the birds face. Likewise, Tengmalm’s owls in Western Finland seem to adjust their clutch sizes in accordance to the 3-year population cycle of voles, their prey, in the area (Korpimäki and Hakkarainen, 1991). In each of these settings, our model suggests that individuals with the ability to forecast environmental change, even if this capability comes at a cost, may be favored by selection.
While we have studied the evolution of a mode of decision-making that forms beliefs about future environmental states, a closely related phenomenon called theory of mind falls outside the scope of our analysis. Theory of mind describes the ability to conceptualize the way in which others make decisions (Apperly, 2012). Future work could study forecasting of others’ strategy dynamics, in addition to or in lieu of environmental forecasting. We expect that strategic forecasting may have qualitatively distinct population-level impacts than environmental forecasting, and it may be favored in different settings.
Throughout our analysis we have assumed the dynamics of the environment and decision-making are deterministic and the cost of forecasting fixed. However, a substantial body of research on what drives bounded rationality has drawn on the finance literature and assumes stochastic dynamics of the environment and payoffs, where the efficacy of forecasting depends on the prevalence of forecasting types (Lo, 2017). Stochastic payoffs that are correlated can give rise to bounded rationality (Brennan and Lo, 2012). This can occur even in the absence of cognitive costs, when there is a risk of over-fitting predictions to noisy outcomes (Lo and Zhang, 2021). Similar models have also shown coexistence of multiple types (Lo and Zhang, 2022) and the breakdown of collective intelligence. This breakdown of collective intelligence is exemplified in interactions among rational traders and noise traders, who respond to hype rather than fundamentals. Such interactions can favor noise traders despite their seemingly misguided behavior (De Long et al., 1990a) and in some settings can generate destabilizing feedback effects that resemble price bubbles (De Long et al., 1990b). Future work that integrates stochastic environmental dynamics and dynamic costs of forecasting could help connect this study with theories of bounded rationality in finance and economics.
We have analyzed a well-mixed model where all individuals of all types and strategies interact. But in some contexts, a hierarchical model might be more realistic. In Supplementary Material Section 1, we describe a hierarchical model where forecasting types and myopic types form two distinct sub-populations that compete with each other at the population level. The hierarchical model produces dynamics of greater complexity than the well-mixed model presented in the main text.
Rather than predicting a steady advance in the sophistication of cognition, our analysis suggests that a more likely outcome is the coexistence of multiple modes of decision-making. This finding aligns with results from psychology that indicate most people have automatic modes of decision-making as well as more deliberative, and mentally taxing, modes (Kahneman, 2011; Rand et al., 2017). Our finding of coexistence between forecasting and myopic types provides theoretical support for behavioral-economic hypotheses about how decisions are actually made. Our results help explain why automatic decision-making persists, and they describe a context in which higher-level cognition can be favored evolutionarily while also bringing public benefits to all cognitive types.
Supplemental Material
Supplemental Material - The evolution of forecasting for decision-making in dynamic environments
Supplemental Material for The evolution of forecasting for decision-making in dynamic environments by Andrew R Tilman, Vítor V Vasconcelos, Erol Akçay, and Joshua B Plotkin in Collective Intelligence
Footnotes
Acknowledgments
The findings and conclusions in this publication are those of the authors and should not be construed to represent any official USDA or U.S. Government determination or policy.
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by the JBP acknowledges funding from the John Templeton Foundation (grant number 62281). VVV and ART acknowledge funding from the United States Department of Agriculture (USDA), Forest Service Northern Research Station (agreement number 22-IJ-11242309-044). EA acknowledges support from BSF Grant 2019156.
Supplemental Material
Supplemental material for this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.