
Abstract
Background:
Maternal prenatal stress is associated with physiologic and adverse mental health outcomes in the offspring, but the underlying biologic mechanisms are unknown. We examined the associations of maternal perceived stress, including preconception exposure, with DNA methylation (DNAm) alterations in the cord blood buffy coats of 358 singleton infants.
Methods:
Maternal perceived stress was measured prior to and throughout pregnancy in a cohort of women enrolled in Effects of Aspirin in Gestation and Reproduction Trial (EAGeR) trial. Perceived stress assessments based on a standardized Likert-scale were obtained in periconception (~2 months preconception and 2-8 weeks of gestation) and pregnancy (8-36 weeks of gestation). Cumulative perceived stress was estimated by calculating the predicted area under the curve of stress reported prior to and during pregnancy. DNAm was measured by the Infinium MethylationEPIC BeadChip. Multivariable robust linear regression was used to assess associations of perceived stress with individual CpG probes.
Results:
Based on a 0 to 3 scale, average reported preconception and early pregnancy stress were 0.76 (0.60) and 0.67 (0.50), respectively. Average mid- to late-pregnancy stress, based on a 0 to 10 scale, was 4.9 (1.6). Neither periconception nor pregnancy perceived stress were associated with individual CpG sites in neonatal cord blood (all false discovery rate [FDR] >5%).
Conclusion:
No effects of maternal perceived stress exposure on array-wide cord blood neonatal methylation differences were found.
Background
The developmental origins of health and disease (DOHaD) hypothesis links early life exposures (periconception, prenatal, postnatal) to long term health outcomes in children and adults. 1 Early life exposure to psychosocial stressors have been implicated in various postnatal or early childhood outcomes, including cognitive abilities or behavioral issues,2-5 as well as overall adult morbidity and mortality. 6 The mechanisms underlying the associations between early life stress exposure and health outcomes remain unclear. One potential mechanism is epigenetic modifications, such as DNA methylation (DNAm), which are made in response to early life exposures.7,8 Such modifications can influence gene expression that may lead to psychopathologic changes or potentially be utilized as biomarkers of future disease risk.9,10 For example in rodents, chronic restraint stress from 14 to 20 days gestation (a measure of prenatal stress) was associated with increased placental DNAm at CpG probes within the 11β-hydroxysteroid dehydrogenase type 2 (HSD11B2) gene promotor that regulates cortisol but decreased expression in HSD11B2. 11
Human studies assessing associations between measures of perceived stress and offspring DNAm alterations in cord blood initially focused on candidate genes rather than an epigenome-wide association study (EWAS) approach.12-16 These early studies found differential methylation in cord blood of NR3C1, 14 a gene involved in hypothalamic-pituitary-adrenal axis (HPA-axis) functioning, but methylation of other HPA-axis genes in cord blood (SLC64A, HSD11B2, or CRHR2) have not been implicated in perceived stress exposure specifically. 12 Further, differential methylation of imprinting control regions has also been identified in cord blood.12,16 More recently, EWAS studies have been conducted to examine prenatal depressive symptoms or stressful life events; however, these studies have not replicated the findings of candidate gene studies.17-20
Previous candidate gene and EWAS studies on prenatal stress have focused on the impacts of exposure during gestation. The timing of exposure is important in the developing epigenome, including exposure in the weeks immediately surrounding conception. 21 However, prior studies examining perceived stress have mostly recruited women in the third trimester and not prior to conception.13-15 Therefore, we aimed to identify novel newborn DNAm markers of periconception and pregnancy perceived stress exposure captured at multiple timepoints using an array-wide approach. Such markers detected at birth may inform a risk of adverse outcomes in early childhood and provide insights into potentially dysregulated mechanisms during early development.
Methods
Study design
The Effects of Aspirin in Gestation and Reproduction (EAGeR) trial (2007-2011; NCT00467363) was a multicenter, double-blind clinical trial that randomized women with a history of pregnancy loss to low-dose aspirin + folic acid (400 µg) versus folic acid prior to conception to investigate effects on reproductive outcomes (n = 1228).22,23 The current analysis is based on participants from the Salt Lake City, Utah study site, which recruited >80% of study participants. Cord blood was collected at Utah beginning in 2009 (2 years after enrollment began) and was successful for over 90% of deliveries thereafter (n = 428). Samples that had insufficient DNA or failed quality control checks were removed (n = 37). We also excluded 33 participants who were not of non-Hispanic white race/ethnicity. These included race (Black (n = 4), American Indian (n = 1), Asian (n = 6), and unidentified (n = 1)) and 21 participants that identified as Hispanic ethnicity since there are race/ethnic differences in self-reported maternal stress and genetic ancestry plays a role in establishment of DNA methylation. 24 Previous analysis of these data found that randomization to low-dose aspirin had no impact on DNA methylation in cord blood. 25 The study was approved by the institutional review board (IRB) at the University of Utah (Salt Lake City, Utah IRB #1002521), and all participants provided written informed consent prior to enrolling.
Perceived stress assessment
At enrollment (preconception), study participants were provided a daily diary to complete during their first 2 menstrual cycles while attempting pregnancy (on average 2 months before pregnancy). Participants were instructed to record their average daily stress on Likert-scale of 0 = no stress, 1 = little stress, 2 = moderate stress, and 3 = a lot of stress. Similarly, participants were instructed to complete a daily diary in early pregnancy (up to 8 weeks gestation) which also captured average daily stress based on the same scale as the preconception dairy. Beginning at 12 weeks gestation, participants completed questionnaires every 4 weeks until 36 weeks gestation which asked “During the last month, how would you describe your stress level on a scale of 0 to 10? (0 = no stress, 10 = maximum stress).” For this analysis, we estimated cumulative periconception perceived stress (based on 0-3 scales captured in preconception and early pregnancy) and pregnancy perceived stress (based on 0-10 scales from 12 to 36 weeks gestation) based on a regression curve of the stress measurements.26,27 We approximated cumulative perceived stress in these 2 periods using linear mixed models with random effects for the intercept and the centered week of stress measurements in each period. For each participant, parameters from the model were then used to define the predicted area under the perceived stress curves. 27
DNAm measurement and processing
The measurement of DNA from cord blood samples has been previously described for this cohort. 25 Briefly, prepared cord blood buffy coat was shipped and processed for DNA extraction at the University of Minnesota. DNA underwent bisulfate conversion (EZ DNA Methylation™ kit, Zymo Research, Irvine, CA, USA). Samples were randomized and sample plate and positions were tracked as DNAm was profiled using the Infinium MethylationEPIC BeadChip microarray.28,29 DNAm microarray data were processed using the minifi package in R, including the quantile normalization and background signal and dye-bias adjustments of probes. 30 Probes on the sex chromosomes were removed. DNAm levels for each CpG probe were reported as β-values ranging from 0 (unmethylated) to 1 (methylated). β-values were determined by calculating the ratio of methylated probe fluorescence intensity to the sum of the methylated and unmethylated probe intensities.
Statistical analysis
For descriptive purposes, cumulative periconception perceived stress and pregnancy perceived stress were categorized by tertile. Chi-square or ANOVA tests were conducted to compare study participant characteristics by perceived stress tertile. To examine the association of maternal perceived stress with methylation β-values as the outcome we used multivariable robust linear regression. Separate models for periconception and pregnancy stress exposures were constructed to evaluate whether timing of stress influences methylation. In a secondary analysis, we examined preconception and early gestation (up to 8 weeks) perceived stress separately using the same approach to estimate cumulative stress exposure as described above. Cumulative stress was operationalized on a continuous scale in all linear models. A false discovery rate (FDR) correction using the method by Benjamini-Hochberg was applied to account for multiple comparisons. 31 All models were parsimoniously adjusted for infant sex, plate number, and cell count estimation. Plate number was used to adjust for batch effects from the measurement of DNAm microarray. Cell count estimation of 6 leukocyte subtypes and nucleated red blood cells was based on a cord blood-specific reference.32,33 Adjustment for infant’s epigenetically derived ancestry using GLINT 34 did not influence results and was thus not included in the final model (data not shown). Other potential covariates were not included in the final model due to lack of statistical association with perceived stress in this analytic sample (Table 1).
Study population characteristics by tertile of periconception or mid-to-late pregnancy perceived stress, n = 358.
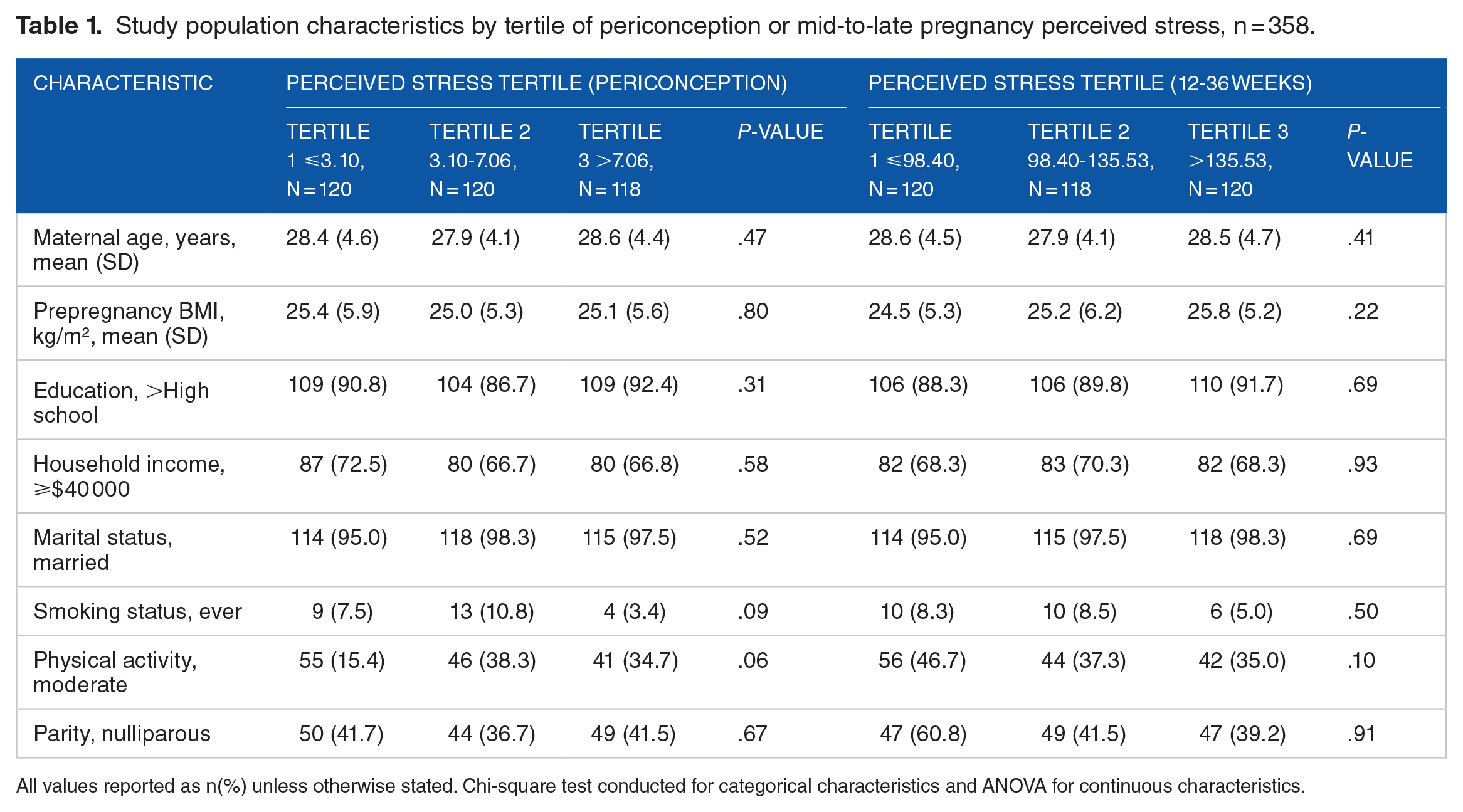
All values reported as n(%) unless otherwise stated. Chi-square test conducted for categorical characteristics and ANOVA for continuous characteristics.
In a secondary analysis, we examined whether perceived stress was associated with differentially methylated regions (DMRs) using the dmrff package and summary statistics from our array-wide analyses as inputs. 35 DMRs were defined as genomic regions that covered a set of CpG probes (nominal EWAS P-values) with at most 1000 bp between consecutive probes and were consistently positively or negatively correlated with the exposure. Analyses were conducted in SAS 9.4 and R 4.0.2.
Gene annotations were identified using the Illumina database and Ingenuity Knowledge Database and verified in the University of California Santa Cruz genome browser (GRCh37/hg19). Biologic networks and functional pathways were generated through the use of Ingenuity Pathway Analysis (IPA, QIAGEN Inc.). 36
Results
Women averaged 28.0 years of age (SD = 4.5), the majority were never smokers (92.1%) and did not report consuming alcohol in their preconception diary (73.1%) (Table 1). Women recorded on average little to no stress on the preconception diaries on a scale of 0 to 3 (mean = 0.76; SD = 0.60; median = 0.50; IQR = 0.50). Similarly, women reported little to no stress in daily diaries in early gestation (mean = 0.67; SD = 0.50; median = 0.64; IQR = 0.86). Throughout the second and third trimester, women reported moderate stress on a scale of 0 to 10 (mean = 4.9; SD = 1.6; median = 5.0, IQR = 3.0).
Neither higher periconception perceived stress nor pregnancy perceived stress were associated with individual CpG probes in neonatal cord blood (FDR P-values <.05). Moreover, no FDR significant differences in methylation were observed when perceived stress in preconception or early gestation (up to 8 weeks) periods were analyzed separately. Tables 2 and 3 provide the top 5 P-value ranked CpG probes from periconception and pregnancy analyses. Coefficients of these top P-value ranked CpG probes suggest less than 1% change with no clear pattern of associations. Of note, the top 5 P-value ranked probes from the periconception analysis were the same as those from the early gestation analysis (ie, not including preconception stress) and the effect sizes were of similar magnitude and direction. Supplemental Table S1 presents the regional analysis results in which we identified 4 DMRs associated with periconception stress and 2 DMRs with pregnancy stress.
Top-rank CpG probes of the periconception perceived stress and cord blood DNA methylation analysis, n = 358.
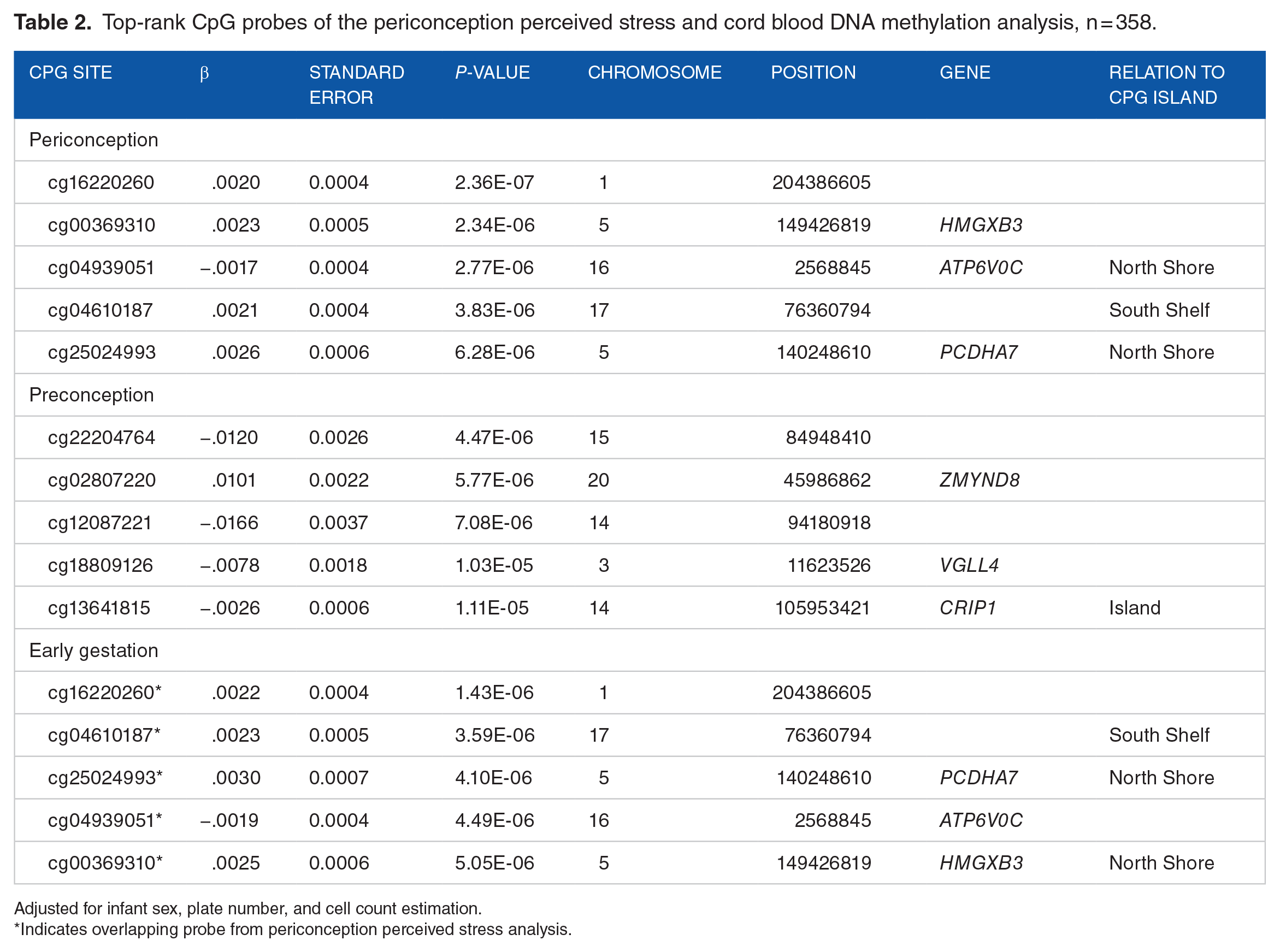
Adjusted for infant sex, plate number, and cell count estimation.
Indicates overlapping probe from periconception perceived stress analysis.
Top-rank CpG probes of the 12 to 36 weeks pregnancy perceived stress and cord blood DNA methylation analysis, n = 358.
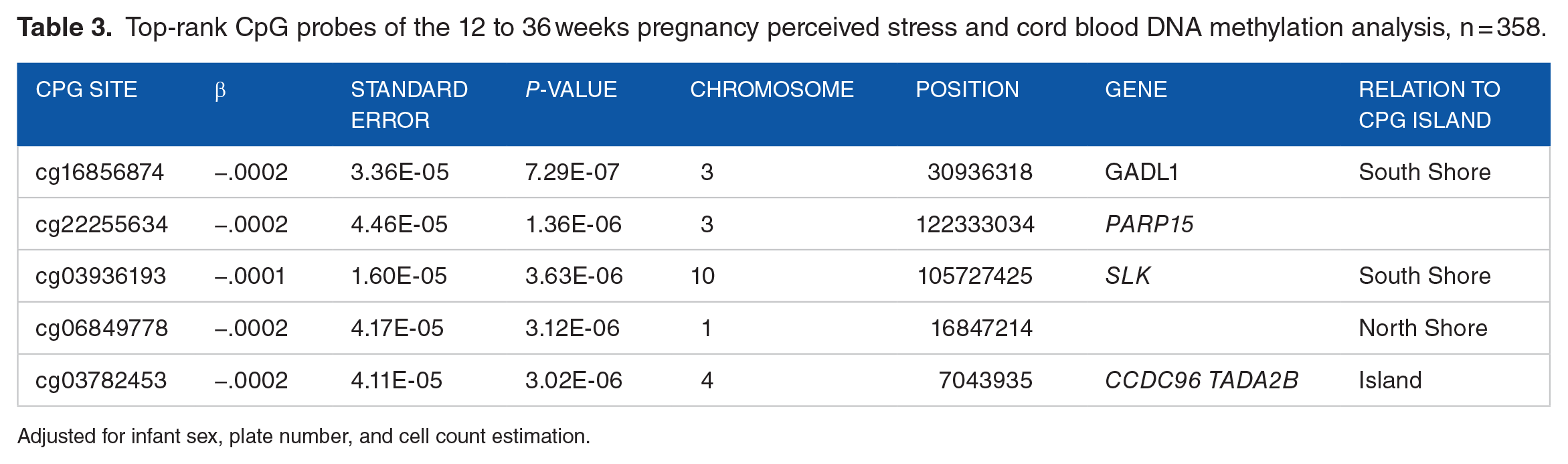
Adjusted for infant sex, plate number, and cell count estimation.
Overlaying the top-100 ranking CpG probes (P-value >.0001) found in the periconception and pregnancy perceived stress array-wide analysis with the IPA Knowledge Base, we identified networks of interconnected genes and potential biologic implications. Supplemental Tables S2 and S3 present the IPA networks identified and all associated molecules. We have included these networks for hypothesis generation considerations only; though it should be noted that pathways identified in IPA are only potential biologic processes that could be impacted by the methylation of these CpG probes. We cannot rule out that the identified pathways may be due to the differing number of probes per gene that are present on the EPIC 850K.
Discussion
We did not find associations between periconception or prenatal perceived stress exposure and differential DNAm markers in cord blood after FDR adjustment. Further, identified DMRs had no clear biological ties to perceived stress exposure. These null findings are consistent with previous EWAS investigations examining objective prenatal stressors like stressful life events and mood disorders with DNAm alterations in cord blood.18,37-39 Previous candidate-gene studies of prenatal stress have focused largely on DNAm alternations in genes related to HPA-axis functioning, such as NR3C1 or FKBP5. The secretion of systemic glucocorticoids (eg, increasing serum cortisol) is the product of the HPA-axis’s response to stressors. During pregnancy, the fetus is beginning to be exposed to maternal cortisol when the placenta forms (~12 weeks) and levels naturally rise during pregnancy. Although cortisol exposure is regulated by the placental enzyme, HSD11B2, 40 prenatal challenges like maternal stress can impact functioning. Epigenetic processes are highly tissue specific. Thus, it is possible that associations of maternal perceived stress and DNAm may be more apparent in offspring tissues other than cord blood, such as the placenta. This is supported by generally low correlations between cord blood and placenta DNA methylation.12,41,42
A growing number of studies have begun to assess the impacts of maternal stress on placental DNAm.12,43-45 For example, higher perceived maternal stress was associated with DNAm of HSD11B2, as well as NR3C1 and FKBP5 in human placenta tissues. 44 Further, retrospectively assessed lifetime maternal stress exposure was associated with placental DNAm aberrations at 112 CpG probes. 43 In comparison to our findings, of these 112 CpG probes 11 probes were identified in our EWAS as having nominal P-values <.05 (cg16717626, cg16900746, cg24335984, cg18941410, cg25545479, cg15365888, cg13817341, cg05218210, cg02721947, cg23095615, and cg01903374). Future work should assess the effects of perceived stress on placental methylation and determine if alterations correlate with transcription.
Periconception is considered a critical period of early development. Though our study did not find a significant effect of maternal perceived stress on DNAm in the offspring at birth, we cannot rule out the potential impacts of periconception or prenatal exposure. First, impacts of more distal stress exposures occurring earlier in the woman’s life course (eg, adolescence) may have influences. 46 Though our study is unique in the assessment of periconception perceived stress, future research from a life course perspective is needed to account for stressors in earlier life. Second, in this same cohort despite the low level of average perceived stress, we have previously shown that higher preconception stress was associated with a longer time to pregnancy 47 and it is possible that perceived stress exposure in the first 8 weeks of gestation may impact implantation and follows the line of work regarding placental differences.
Our study is unique in our ability to prospectively assess perceived stress in the context of both periconception and prenatal exposure. We had multiple measures of perceived stress before and throughout pregnancy. However, the assessment of perceived stress was not based on a validated instrument, such as the Perceived Stress Scale 48 or the Life Stressor Checklist 49 as used in prior studies.12,14,16,43 Further, the generalizability of our findings may be limited as the study population consists of non-Hispanic white women with 1 to 2 pregnancy losses who then went on to have a live birth and who were of higher socioeconomic status. We also did not collect information on paternal levels of stress that may impose epigenetic effects on offspring50,51; nor did we have information on epigenome-wide associations in relevant tissues, like the placenta, both of which should be considered in future studies. In summary, we found no evidence of the effects of periconception or pregnancy perceived stress exposure on newborn methylation in cord blood leukocytes.
Supplemental Material
sj-docx-1-gae-10.1177_25168657221082045 – Supplemental material for Periconception and Prenatal Exposure to Maternal Perceived Stress and Cord Blood DNA Methylation
Supplemental material, sj-docx-1-gae-10.1177_25168657221082045 for Periconception and Prenatal Exposure to Maternal Perceived Stress and Cord Blood DNA Methylation by Kristen J Polinski, Diane L Putnick, Sonia L Robinson, Karen C Schliep, Robert M Silver, Weihua Guan, Enrique F Schisterman, Sunni L Mumford and Edwina H Yeung in Epigenetics Insights
Footnotes
Funding:
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: Supported by the Intramural Research Program of the Eunice Kennedy Shriver National Institute of Child Health and Human Development (contract numbers HHSN267200603423, HHSN267200603424, HHSN267200603426, and HHSN275201300023I-HHSN2750008).
Declaration of conflicting interests:
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Authors’ Note
Registered as NCT00467363 at clinicaltrials.gov.
Author Contributions
KJP, EFS, and EHY contributed to the conception or design; WG provided essential materials; KJP and SLR contributed to the analysis; DLP, KCS, RMS, WG, and SLM contributed to the interpretation of data; KJP drafted the manuscript; DLP, SLR, KCS, RMS, WG, EFS, SLM, and EHY provided essential feedback in writing the manuscript; and all authors approved the final manuscript and are accountable for the work.
Availability of Data
Data described in the manuscript will be made available upon request pending application and approval.
Supplemental Material
Supplemental material for this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.