
Abstract
Checkpoint markers and immune checkpoint inhibitors have been increasingly identified and developed as potential immunotherapeutic targets in various human cancers. Despite valuable efforts to discover novel immune checkpoints and their ligands, the precise roles of their therapeutic functions, as well as the broad identification of their counterpart receptors, remain to be addressed. In this context, it has been suggested that various putative checkpoint receptors can be induced upon activation. In the tumor microenvironment, T cells, as crucial immune response against malignant diseases as well as other immune central effector cells, such as natural killer cells, are regulated via co-stimulatory or co-inhibitory signals from immune or tumor cells. Studies have shown that exposure of T cells to tumor antigens upregulates the expression of inhibitory checkpoint receptors, leading to T-cell dysfunction or exhaustion. Although targeting immune checkpoint regulators has shown relative clinical efficacy in some tumor types, most trials in the field of cancer immunotherapies have revealed unsatisfactory results due to de novo or adaptive resistance in cancer patients. To overcome these obstacles, combinational therapies with newly discovered inhibitory molecules or combined blockage of several checkpoints provide a rationale for further research. Moreover, precise identification of their receptors counterparts at crucial checkpoints is likely to promise effective therapies. In this review, we examine the prospects for the application of newly emerging checkpoints, such as T-cell immunoglobulin and mucin domain 3, lymphocyte activation gene-3, T-cell immunoreceptor with Ig and ITIM domains (TIGIT), V-domain Ig suppressor of T-cell activation (VISTA), new B7 family proteins, and B- and T-cell lymphocyte attenuator, in association with immunotherapy of malignancies. In addition, their clinical and biological significance is discussed, including their expression in various human cancers, along with their roles in T-cell-mediated immune responses.
Introduction
Tumor-associated antigens or immune checkpoint molecules play a pivotal role in the immune regulation of malignancies whose biological significance is crucial in cancer diagnosis, prognosis, and treatment. The checkpoints that are placed either on the surface of various immune cells including T lymphocytes or on tumor cells act like a switching protein through the induction of various signals to control the over-activation of T cells. In cancers, T-cell dysfunctionality may be due to persistent antigen exposure that is associated with overexpression of multiple inhibitory receptors that ultimately reduce T-cell proliferation capacity or functionality. Blockade of checkpoint proteins with immune checkpoint inhibitors prevents the ‘off’ signal from being sent while the blocked condition accumulates and increases T cells in the blood circulation that further amplifies their movement into the tumor to kill cancer cells. In addition, immune checkpoint therapies may consider a novel treatment strategy for infectious diseases and autoimmune disorders. Interestingly, under normal circumstances, inhibitory checkpoint proteins prevent autoimmune damage by suppressing immune responses. In malignancies, the cancerous cells hire mechanisms by which they can protect themselves from being attacked by immune cells through stimulating immune checkpoint targets. 1 To rescue the recognition of the immune response or to maintain homeostasis of the immune system while reducing undesirable immune responses, tumor cells continuously modify their expressions at their molecular level, while in this context, the types of cancers and their various stages significantly influence the compositions of the tumor microenvironment (TME). A great body of research has emphasized that in the TME, immune checkpoint molecules play an important role in the progression and outcome of tumors, and intensive research has eventually led to the development of various cancer immunotherapeutic reagents including monoclonal antibodies. Further to receptors and ligands of immune checkpoints on the cell membrane, various soluble immune checkpoints have been also discovered.2,3 The soluble immune checkpoints that circulate in the serum of patients are produced either under alternative splicing of mRNA or under cleavage of membrane-bound proteins. These circulating isoforms also play a crucial role in cancer immunotherapy by the interaction between soluble receptors or soluble ligands with full-length ligands or full-length receptors, respectively. Despite the aforementioned therapeutic competence of the immune checkpoints, these circulating biomarkers may additionally be useful for the determination of the prognosis of various cancers or/and screening of patients as a first line of diagnosis prior to intensive and invasive cancer diagnostic approaches.4–6 Among various types of surface and soluble receptors or ligands, cytotoxic tumor lymphocyte antigen 4 (CTLA-4) and programmed cell death-1 (PD-1)/programmed cell death-ligand 1 (PD-L1) are the most studied molecules for which blocking antibodies are developed and currently ipilimumab and pembrolizumab have been examined in various clinical trials, respectively.7–9 In this review, we discuss the role of recently targeted checkpoint molecules, including T-cell immunoglobulin and mucin domain 3 (TIM-3), lymphocyte activation gene-3 (LAG-3), T-cell immunoreceptor with Ig and ITIM domains (TIGIT), V-domain Ig suppressor of T-cell activation (VISTA), new B7 family proteins, and B- and T-cell lymphocyte attenuator (BTLA) in the diagnosis, prognosis, and treatments of various cancers.
Classical checkpoints regulators PD-1 and CTLA-4 and their recent clinical application
Adaptive and innate immune systems both interact independently or complementary via the expression of a wide family of inhibitory receptors. Of interest, malignant cells can escape or exhaust the immune system through alteration of the dynamic balance between stimulatory and inhibitory signaling molecules. In this context, T cells, B cells, and natural killer (NK) cells exhibit coinhibitory molecules belonging to the B7-CD28 group of proteins such as CTLA-4 and PD-1 receptors that can adjust the production of antigen-specific T cells while negatively regulating immune tolerance. 10 The PD-L1 that is expressed on antigen-presenting cells (APCs) or cancer cells serves as a ligand for the PD-1 receptor, binds to PD-1 on immune cells, and suppresses immune responses through either suppression of T-cell activation or escaping of cancer cells from immune surveillance. 11 In clinical studies, benefits of monotherapy either with PD-1 or CTLA-4 blockers were associated with low response rates while few patients were found to have a response to therapy. 12 A number of studies revealed that the majority of metastatic melanoma patients could not respond properly to ipilimumab, nivolumab, and pembrolizumab as evaluated by objective response rate.13,14 On the other hand, it has been confirmed that a combination of CTLA-4 and PD-1 blockade can improve the response rates along with the survival rates in multiple cancer types including melanoma, renal cell carcinoma (RCC), colorectal cancer, small-cell lung cancer, mesothelioma, and sarcoma.15–20 Although combining CTLA-4 and PD-1 blockers has been shown to improve treatment outcomes in multiple cancer types, it has been associated with the induction of adverse events in affected patients. In this regard, besides further research to evaluate the combination of other CTLA-4 blockers (tremelimumab) and other PD-1/PD-L1 blockers (cemiplimab, durvalumab, avelumab, and atezolizumab) and their associated dose titration or administration sequence, the search for efficacious combination therapies with newly discovered inhibitory molecules continues.
T-cell immunoglobulin and mucin domain 3
TIM-3, also known as hepatitis A virus cellular receptor 2, which is encoded by HAVCR, comprises TIM-1, TIM-3, and TIM-4 in humans and Tim-1 through Tim-8 in mice. 21 Among various tumor-infiltrating cells in the TME, Tim-3 as the cell terminal dysfunction biomarker is mostly expressed at higher levels by cluster of differentiation 8 (CD8+) tumor-infiltrating lymphocytes (TILs) and cluster of differentiation 4 (CD4+) regulatory T cells (Tregs) and until now, four specific ligands including galectin-9, phosphatidylserine (PtdSer), high mobility group protein B1 (HMGB1), and carcinoembryonic antigen-related cell adhesion molecule-1 (CEACAM-1) have been introduced for this immune checkpoint. 22 Further support for the role of TIM-3 is provided by the analysis of genetic polymorphisms. A recent study indicates that TIM-3 and lectin, galactoside-binding soluble 9 (LGALS9) polymorphisms are associated with clear cell renal cell carcinoma (ccRCC) risk and overall survival (OS). 23 Generally, and as mentioned earlier, the dysfunctionality of T cells in malignancies is due to loss of cytotoxicity or deficiency in the production of pro-inflammatory cytokines along with increased expression of various checkpoint proteins. Hence, it has been speculated that antagonizing transcription factor T-cell factor 1 (TCF-1) with Tim-3 can reduce the stemness of CD8+ T cells, the cells that are crucial mediators of tumor clearance. TCF-1 probably maintains stemness and restrains effector differentiation in CD8+ T cells; however, the contribution of Tim-3 to terminal dysfunction of CD8+ T cells is not clearly understood. Elevated expression levels of Tim-3 have also been reported on CD4+ Tregs in human and murine tumors, which were further associated with Foxp3 expression. Interestingly, 60% of fox3+ TILs were found to be TIM-3+, a condition that highlights the crucial role of Tim-3+ Tregs in advanced tumor stages. 24 Furthermore, TIM-3 expression in non-T cells, including macrophages, dendritic cells (DC), and NK cells, also has been reported in several studies; however, the regulation of Tim-3 expression in these cells remains to be uncovered.25–27 Besides the influence of various transcriptional factors, diverse signaling pathways additionally influence the regulation of Tim-3 expression on T cells. For instance, both nuclear factor interleukin 3 regulated (Nfil3) and interleukin (IL)-27, IL-15, IL-12, and IL-35 as novel members of the IL-12 cytokine family upregulate TIM-3 expression in humans and animal models of various malignancies.28–31 TIM-3+, PD-1+, and CD8+ TILs have shown inhibition of the production of the cytokines such as tumor necrosis factor-α (TNF-α), interferon-gamma (IFN-γ), and IL-2. 32 Interestingly, treatment with TIM-3 blockade antibody significantly increased IFN-γ and IL-22/IL-17 levels while it decreased IL-10, that is correlated with enhanced Th1- and Th17-mediated immune response and decreased Treg immune response, respectively. 33 As a prognostic marker in cancer, Tim-3 expression has shown an association with negative outcomes and poor OS in several solid tumors while it can be a valuable therapeutic target. 34 In a study conducted by Pu et al., 35 high expression of TIM-3 was significantly associated with an increased risk of mortality among patients with primary osteosarcoma. In fresh tumor tissue samples of these patients, overexpression of TIM-3 further was reported. Importantly, a positive correlation of TIM-3 expression with lymph node metastasis, tumor grade, and PD-1 expression should not be neglected. 34 In hematological cancers, a recent study with broad immunogen analysis that investigated immune cell co-stimulation/inhibition and cancer antigen expression patterns in association with cancer subtypes or genomics deserves particular attention. 36 In this context, the influence of cancer cell molecular phenotype on immune cell infiltration and function has been described elsewhere. 36 Recently, it has been revealed that in patients with acute myeloid leukemia (AML), there is a correlation between the levels of TIM-3 expression and NK cells cytotoxicity. In this context, the elevated TIM-3+ cell expression within the circulating NK cells has shown positive prognostic value in patients with M1+2 grade but not M4+5. This study indicates that NK cell circulation along with TIM-3 expression levels serves as positive prognostic biomarkers in effective immunity against AML. 37 Consistent with this study, in patients with newly diagnosed AML, the high TIM-3 expression from peripheral blood (PB) samples was also associated with decreased OS rates suggesting that TIM-3 might be considered as a weak prognosis biomarker in patients with AML. 38 In patients with chronic lymphocytic leukemia (CLL), an increased Tim-3 expression pattern on NK cells was observed in comparison to healthy controls. 39 Consistently, low hemoglobin level, high absolute lymphocyte count, and high serum C-reactive protein level have been reported as a poor prognostic profile. Recently, an in vitro investigation on CD8+ T cells isolated from PB of patients with early-stage CLL who were treated with anti-PD-1 and anti-TIM-3 blocking antibodies showed no improvement in CD8+ T cell proliferation. 40 This observation indicates that pretreatment of CD8+ T cells with blocking antibodies in CLL patients does not influence the restoration of CD8+ T-cell functionality, at least in the early clinical stages. These findings provide a rationale for further in vitro and in vivo studies to investigate the efficacy of checkpoint inhibitors for CLL patients. From an epigenetic regulation point of view, the regulation of TIM-3 via DNA methylation of the encoding genes and also its crucial ligand galectin 9 (LGALS9) in association with molecular and immune correlates in malignant melanoma and patients’ survival deserves attention. It has been revealed that DNA methylation has a pivotal role in various biological activities of T cells including T-cell differentiation or exhaustion and tumorigenesis. Holderried et al. 41 have found that TIM-3 and LGALS9 mRNA expression and methylation levels correlated significantly with tumor immune cell infiltration. Although the expression of TIM-3 is associated with T-cell exhaustion, they reported a significantly better OS in association with high TIM-3 and LGALS9 mRNA expression. For both TIM-3 and LGALS9, significant positive correlations between mRNA expression and gene body methylation were reported while inverse correlations between mRNA expression with promoter methylation were identified. Since TIM-3/LGALS9 methylation levels were evaluated in isolated immune cells, melanocyte, and melanoma cell lines, it would be of interest to examine methylation analysis of isolated, exhausted T cells, to understand whether hypomethylation of 5′-C-phosphate-G-3′ (CpG) sites located in the TIM-3 promoter area might serve as a surrogate biomarker for T-cell exhaustion. Finally, it has been reported that TIM-3 expression in melanoma cells is associated with non-responsiveness to PD-1 immune checkpoint blockade (ICB). Importantly, the upregulation of TIM-3 has been observed after PD-1-targeted ICB. Therefore, the analysis of TIM-3 methylation could be a promising predictive approach to select the subgroup of melanoma patients who would benefit from TIM-3-targeted ICB. In addition, the increased co-expression of TIM-3 along with PD-1, CTLA-4, LAG-3, and TIGIT on TILs in early breast cancers (BCs) has indicated these immune checkpoints might synergistically inhibit the response to the tumor. 42 Hence, from a therapeutic point of view, the combined inhibition of these immune checkpoints may synergistically enhance the T-cell response to various tumor antigens. In this context, the in vitro study of combined PD-L1 and TIM-3 blockade indicates enhanced expansion of fit human CD8+ antigen-specific T cells for adoptive immunotherapy. 43 Recently, various clinical trials include TIM-3 as a novel target in cancer immunotherapy. Ongoing clinical trials with Tim-3-specific monoclonal antibodies or antagonist agents are presented in Table 1.
Current ongoing clinical trials for Tim-3-specific monoclonal antibodies or antagonist agents.

aHSCT, autologous hematopoietic stem cell transplantation; ESCC, esophageal squamous cell carcinoma; HNSCC, squamous cell carcinoma of the head and neck; HSCT, autologous hematopoietic stem cell transplantation; LAG-3, lymphocyte activation gene-3; RCC, renal cell carcinoma; TIM-3, T-cell immunoglobulin and mucin domain 3.
Lymphocyte activation gene-3
LAG-3 is a transmembrane molecule closely related to CD4, which has been reported to be expressed in multiple immune cells, including T cells, NK cells, natural killer T (NKT) cells, Treg, activated B cells, natural plasma cells, and plasmacytoid dendritic cells (pDCs).44–48 Due to the structural similarity between LAG-3 and CD4, it has been proposed that major histocompatibility complex class II (MHC-II) molecules are ligands for LAG-3 with approximately 100 times greater binding affinity than CD4. 49 Alternative ligands include galectin-3, a 31-kDa lectin that has been shown to modulate T-cell responses, and liver sinusoidal endothelial cell lectin (LSECtin), which is expressed in liver or human melanoma tissues with its growth-enhancing activity through inhibition of antitumor T-cell-dependent responses.50–52 In melanoma cells, IFN-γ production was inhibited by the interaction between LAG-3 and LSECtin through mediating antigen-specific effector T cells. 52 Since LAG-3 influences CD8+ T-cell function as well as CD4+ T cells, the two alternative ligands mentioned above may serve to explain the intrinsic role of LAG-3 on CD8+ T cells in the TME. Furthermore, in a mouse model of Parkinson’s disease, LAG-3 was reported to bind with α-synuclein that produces fibrils in the central nervous system and this finding further indicates a role of LAG-3 immunologically relevant ligands apart from its potential in TME. 53 In tolerogenic conditions like chronic inflammation or tumor environment, LAG-3 co-expression with other immune checkpoints including PD-1, TIGIT, TIM-3, 2B4, and CD160 on immune and cancer cells can lead to functional exhaustion along with a reduction in proliferation and depletion of cytokine secretion. 54 Notably, the inhibitory effects of LAG-3 on CD8+ T cells are different from those of CTLA-4 or PD-1. In this context, the biochemical study of the PD-1 signaling pathway has shown an increased dephosphorylation of the CD28 receptor upon PD-1 ligation with PD-L1 in comparison with the T-cell receptor (TCR). 55 Equivalently, CTLA-4 inhibits T cell activities via competitive binding with CD80 and CD86 co-stimulatory ligands for CD28. As an alternative mechanism, CTLA-4 may delete CD80/CD86 from APCs. By contrast, LAG-3 may decrease T-cell activation through a TCR reducing pathway. The different mechanism of action between LAG-3 and PD-1 or CTLA-4 highlights the importance of synergistic enhancement of its combined antitumor blockade with PD-1. 56 In Table 2, clinical trials involving combined LAG-3 with PD-1, CTLA-4 checkpoints blockade, or with bispecific agents targeting LAG-3 are presented. In addition, several bispecific agents targeting LAG-3 deserve attention. FS118 is a dual antagonist bispecific agent targeting both LAG-3 and PD-L1 that enhances T-cell activation with enhanced antitumor activity. 57 A phase I first-in-human study of FS118, in patients with advanced cancer and PD-L1 resistance, was well tolerated with no dose-limiting toxicity (DLTs). This result highlights the need for further investigation to determine its clinical benefit in patients who have become refractory to anti-PD-(L)1 therapy. 58 In preclinical studies, FS118 could clear MC38 tumor cells while it induces the shedding of LAG-3. The reduced surface expression of LAG-3 and elevated soluble sLAG-3 (sLAG-3) in the serum indicate its potential importance for combined immunotherapy. MGD013 is another bispecific dual affinity retargeting antibodies (DART) molecule that binds PD-1 and LAG-3, and phase I clinical trials in various malignancies have been undertaken. Elevated LAG-3 mRNA expression has also been reported in the red pulp of the spleen, cerebellum, and thymic medulla. 59 Indeed, altered LAG-3 expression together with its cleavage from the surface of immune or tumor cells governs optimal T-cell activity. Notably, through this cleavage, sLAG-3 is released into circulation, although its biological function has not been clearly understood.60,61 A poor expression of sLAG-3 in PB was positively associated with IL-12 and IFN-γ expression in patients with gastric cancer (GC) and increased sLAG-3 expression has shown better prognosis in these patients. 62 Importantly, the in vivo experiments with GC-bearing mice revealed that elevated levels of sLAG-3 may inhibit tumor growth, along with promotion in the secretion of CD8+ T cells, IL-12, and IFN-γ. Hence, prolonged OS along with increased survival rate was correlated with increased levels of sLAG-3 in GC-bearing mice. In a study of human BC expressing estrogen or progesterone receptors, a high level of sLAG-3 was also associated with better OS. 63 In patients with early BC, following treatment with neoadjuvant chemotherapy significantly increased levels of soluble co-inhibitory checkpoints including LAG-3, PD-L1, and TIM-3 have been reported that may indicate the recovery of immune homeostasis in treated patients. 64 Inconsistently, in CLL and ccRCC patients, sLAG-3 was found lightly associated with poor survival, while in the CLL group, the sLAG-3 could enhance leukemic cell activation along with anti-apoptotic properties.65,66 A recent study has shown a correlation between elevated levels of sLAG-3 and advanced tumor stage in ccRCC patients. 66 Elevated sLAG-3 expression has also been reported in Treg cells in the PB of patients with advanced melanoma and colorectal cancer. 67 This contradiction could be explained by the differential regulatory role of sLAG-3 in governing interaction between LAG-3 and MHC-II or by the individual immune response at different cancer sites.68,69 Last but not least, a negative correlation between sLAG-3 expression and CD8A (cluster of differentiation 8a; T-cell marker) in ccRRCC patients may indicate that elevated sLAG-3 may influence T-cell suppression at the tumor site, which may lead to cancer development in these patients. In addition, in virally mediated tumors, overexpression of the LAG-3 for Epstein–Barr virus (EBV) in GC and human papillomavirus (HPV) in cervical cancer and head and neck squamous cell (HNSC) cancer deserves attention. Significant overexpression of LAG-3 has been reported in EBV+ tumors in gastric cancer (stomach adenocarcinoma), and HPV+ tumors in cervical [cervical squamous cell carcinoma and endocervical adenocarcinoma (CESC)] and HNSC cancer. 70 Interestingly, while the expression of PD-1 and CTLA-4 immune checkpoints were also significantly increased in EBV+ GC and HPV+ cervical cancer, neither the ligands of PD-1 nor the ligands of CTLA-4 were upregulated in HPV+ HNSC cancer. 70 Hence, regarding the overexpression of LAG-3 in HPV+ HNSC cancers, LAG-3 blockade alone or in combination with other immune checkpoint pathways blockade including PD-1 might be of particular benefit. However, the results from a recent phase I/II clinical trial of the LAG-3 inhibitor ieramilimab (LAG525) either with anti-PD-1 spartalizumab (PDR001) or alone in patients with advanced malignancies deserve particular attention. 71 Although ieramilimab was well tolerated as monotherapy and in combination with spartalizumab, a moderate response was seen with combined treatment in patients. Hence, with respect to LAG-3 biology, there are still serious debates including whether ligands other than the MHC-II can be actively involved in the course of immunotherapy. Clinical trials that have been conducted so far on LAG-3 are mainly in combination with the PD-(L)1 axis, hence, whether LAG-3 combination with other inhibitory molecules may show promising results or not should be discovered with further experiments. Certainly, data collection from completed and ongoing clinical trials would be extremely helpful to test the hypothesis. Table 2 summarizes the characteristics of anti-LAG-3-specific monoclonal antibodies or antagonists currently used in clinical trials.
Current ongoing clinical trials for LAG-3-specific monoclonal antibodies or antagonist agents.

CTLA-4, cytotoxic tumor lymphocyte antigen 4; Fc, fragment crystallizable; GC, gastric cancer; HNSCC, squamous cell carcinoma of the head and neck; LAG-3, lymphocyte activation gene-3; mAb, monoclonal antibody; NSCLC, non-small cell lung cancer; PD-1, programmed cell death-1; PD-L1, programmed cell death-ligand 1; PK, pharmacokinetics; SCCHN, squamous cell carcinoma of the head and neck.
T-cell immunoreceptor with Ig and ITIM domains (TIGIT)
TIGIT, which belongs to the poliovirus receptor (PVR)precursor family, has one extracellular immunoglobulin (Ig) variable domain and a short intracellular domain, while a type I extracellular transmembrane domain shares sequence homology with DNAX accessory molecule-1 (DNAM-1), CD96, CD155, CD111, CD112, CD113, and poliovirus receptor-related 4 (PVRL4). 72 The short intracellular domain includes one immunoreceptor tyrosine-based inhibitory motif (ITIM) and one immunoglobulin tyrosine tail-like motif where both the extracellular and the intracellular domains share 58% sequence homology between humans and mice while in contrast, the ITIM has an identical sequence in mice and humans.72,73 The low expression of TIGIT has been reported on naïve cells including CD4+ T cells, CD8+ T cells, and Tregs along with NK cells and its expression is upregulated to the highest levels in those activated cells that are influenced by various oncogenic expression factors.72,74 Three ligands have been identified for TIGIT including CD112 (also known as nectin-2), CD113, and CD155 (also known as PVR) while the last one is the main ligand in both humans and mice. 75 Notably, most human malignancies have shown overexpression of CD155 and CD112 on various immune cells and in hematopoietic or non-hematopoietic tissue, respectively.76–78 Upregulation of CD155 and CD112 has been found to be caused by IFN-γ in tumor cells. 79 Consistently, it has been also proven that cytokines are produced by DCs through interaction between TIGIT and CD155. 72 TIGIT inhibits effector T cells and NK cells via either cell-extrinsic or cell-intrinsic manner and through interaction with CD155 or DNAM-1 ligand. 80 In addition, the direct inhibitory signals delivered by TIGIT via interaction with PVR and PVRL2 that inhibits human NK cell cytotoxicity are significant. 74 In this context, the mechanisms of action of TIGIT have been recently reviewed elsewhere. 81 Despite the in-depth discovery of the mechanism of action of TIGID, it is still unclear whether all these pathways are active in every TIGIT expression cell or whether each individual immune cell acquires a specific mechanism of action. As we indicated above, on TILs, TIGIT expression increases and its upregulation has been reported in various cancers, including BC, melanoma, GC, chronic myeloid leukemia, colorectal cancer, nasopharyngeal carcinoma, and non-small-cell lung cancer (NSCLC).82–89 For the mentioned malignancies, CD8+ T cells have shown significantly increased levels of TIGIT expression; however, elevated TIGIT levels have been also reported on tumor-infiltrating Treg and NK cells. There are several disappointing clinical outcomes and the expression of TIGIT includes the development of metastases, poor survival, and disease severity along with relapse post-transplantation. Consistent with these findings, there is a study that indicates TIGIT deficiency produced potent antitumor immunity with protection contra B16 experimental lung metastasis in mice. 90 On the other hand, in myelodysplastic syndrome (MDS), which is characterized by abnormal quality and quantity of blood cells including PB cytopenia and dysfunctional bone marrow hematopoiesis, high expression of TIGIT was reported in PB NK and T cells, which was involved in disease progression along with immune escape of MDS. 91 Furthermore, the increased expression of TIGIT in NK, CD8+, and CD4+ restricts the function of NK and T cells through decreasing cytokines expression including IFN-γ, TNF-α, and CD107a. 91 Decreased expression levels of aforementioned cytokines were also observed in higher-risk MDS patients compared to lower-risk patients. Interestingly, TIGIT+ T and NK cells show significantly increased proliferation compared to TIGIT− T and NK cells. The observed results revealed that TIGIT can be considered as a negative immune checkpoint in MDS, with inhibitory effects on cytokines secretion and proliferation ultimately leading to inhibition of antitumor immune response in MDS patients. Consistent with this conclusion, Han et al. 92 recently reported new pathways of antitumor response by TIGIT blockade through analysis of fragment crystallizable gamma receptor (FcγR) interaction and myeloid cell activation. 92 The authors found that the anti-TIGIT therapeutic effect rather is achieved by possible reverse activating signals via FcγRs on myeloid cells with induction of cytokines and chemokines expression, than depletion of TME Treg or any other immune cell expressing TIGIT. The production and control of various cytokines through activating FcγRs have been also previously reviewed in human pathogen defense and autoimmunity. 93 In a recent study on patients with oral squamous cell carcinoma (OSCC), a dysfunctional phenotype including low secretion of IL-2, TNF-α, and IFN-γ was reported for TIGIT highly expressed CD4+ and CD8+ T cells from PB mononuclear cells and TILs. 94 Notably, inhibitory functions such as high expression of Foxp3 and elevated levels of IL-10 were also measured for TIGIT+ CD4+ T cells in these patients. Last but not least, in this study, the in vitro proliferation and cytokine production of CD4+ and CD8+ T cells improved with TIGIT blockade. Connected to these findings, the Fc-dependent antitumor capabilities of anti-TIGIT antibodies, which are independent of Treg depletion, deserve attention in malignant diseases. Hence, a novel design of a new class of engineered Fc-antibodies may enhance the therapeutic abilities of anti-TIGIT antibodies improving their antitumor immune response either alone or in combination with other ICB. In this context, the simultaneous blockade of three different checkpoint receptors including PD-1, LAG-3, and TIGIT along with NBTXR3-enhanced localized radiation combinatorial therapy in an anti-PD1-resistant lung cancer model in mice, has shown improved therapeutic efficacy. 95 Consistently, in preclinical studies of the ICB-resistant colorectal tumor model MC38 expressing human carcinoembryonic antigen, the combined treatment strategy of three independent pathways TIGIT:CD155, PD-1/PD-L1, and transforming growth factor beta (TGF-β) has shown significant antitumor efficacy. 96 In addition, the combination of atezolizumab (anti-PD-L1) and tiragolumab (anti-TIGIT) treatment in an ex vivo assay for microsatellite stable colorectal tumor cells could restore CD4 and CD8 TILs functionality in this type of cancers which are also resistant to anti-PD-1/PD-L1 therapy. 97 Finally, the recent study of dose escalation for etigilimab (anti-TIGIT antibody) alone or in combination with nivolumab (anti-PD-1 antibody) in patients with locally advanced or metastatic solid tumors showed an acceptable safety profile that warrants further research in clinical trials. 98 Besides, recently developed bispecific PD-L1/TIGIT antibodies have shown promising results in preclinical studies including increased OS in transgenic mice. 99 The bispecific PD-L1/TIGIT antibodies have also shown improved human IL-2 secretion by primary human T cells. Consistently, by applying CD122-directed IL-2 complexes in mice treated with radiation and anti-PD-1, the circulating stem-like CD8+ T cells were increased. 100 Notably, a recent study has also indicated that low-affinity IL-2 (laIL-2) to PD-1+ T cells could induce better tumor control with lower toxicity. 101 It should be mentioned that in this study, the laIL-2 could neither activate peripheral CD8+ nor Treg cells due to low binding features to IL-2Rα and IL-2Rβ, while it conversely was able to activate CD8+ T cells in the tumor particularly when coupled with anti-PD-1. Until now, there are two bispecific antibodies targeting PD-L1/TIGIT including HLX301 and PM1022, from Henlius Inc. and Biotheus Inc., respectively, which have been under study. Ongoing clinical trials for TIGIT-specific monoclonal antibodies or antagonists are presented in Table 3.
Current ongoing clinical trials for TIGIT-specific monoclonal antibodies or antagonist agents.

CAR-T, chimeric antigen receptor T; EGFR, epidermal growth factor receptor; LAG-3, lymphocyte activation gene-3; NSCLC, non-small-cell lung cancer; PD-1, programmed cell death-1; PD-L1, programmed cell death-ligand 1; PFS, progression-free survival; TGF-β, transforming growth factor beta; TIGIT, T-cell immunoreceptor with Ig and ITIM domains.
V-domain Ig suppressor of T-cell activation
VISTA is a type I Ig membrane protein with 55–65 kDa molecular weight which is also known as differentiation of embryonic stem cells 1, platelet receptor Gi24 precursor, B7-H5, SISP1, death domain 1α (DD1α) and programmed death protein-1 homolog (PD-1H) that belongs to the B7-family while shares 22% homology with PD-L1 but has large structural differences with CD276, CD80, and CD86 among the B7 family. 102 VISTA expression on hematopoietic and myeloid cells has been addressed together with its high expression on mature APCs, which are analyzed for their high CD11b. The extrinsic transformation inhibitory signals induced to T cells by VISTA, when expressed on APCs, further address its action as a ligand on myeloid cells, particularly, APCs. 103 In addition, V-Set and immunoglobulin domain-containing 3 (VSIG-3) is also known as immunoglobulin superfamily member 11 (IGSF11) or brain-specific testis-specific immunoglobulin superfamily (BT-IgSF), and has been identified recently as a ligand of the B7 family member VISTA/PD-1H, and interestingly its inhibitory effect on human T-cell functions through a novel VSIG-3/VISTA pathway has been unveiled. 104 It should be noted that, despite the structural similarity of the VISTA extracellular domain to PD-L1, there is no association of VISTA with the CD28-B7 family. Therefore, the pathways through which VISTA and PD-1 checkpoints govern their inhibitory functions are independent. 105 Finally, but not least, the coinhibitory receptor P-selectin glycoprotein ligand-1 (PSGL-1) has recently been shown to interact with multiple histidine residues in the extracellular domain of VISTA particularly at the acidic pH, a condition that mostly found in TMEs. At acidic pH conditions, the R chain of histidine looses protons and the subsequent post-translational modification facilitates binding to the ligand. 106 Importantly, VISTA-mediated immune suppression is inhibited in vivo by antibodies that block the associated interaction in acidic environments. 106 Remarkably, the insignificant VISTA expression on CD8+, CD4+, Treg, and TILs also deserves attention. 105 On CD4+, VISTA act like a co-inhibitory receptor and its action inhibits T-cell activation, propagation, and cytokines production via anti-CD3 activation. 107 Consistently, VISTA−/− CD4+ T cells have shown strong antigen-specific proliferation along with cytokine production as compared with naïve T cells and further VISTA-neutralizing monoclonal antibodies attenuate VSIG-3-induced T-cell inhibition via decreasing the binding of VSIG-3 and VISTA.104,108 In hematological malignancies, the potential roles of VISTA in immunotherapy also deserve attention. For example, in a study performed by Pagliuca et al., 109 the elevated VISTA expression can influence a patient’s response to chemotherapy by impeding the immune response and facilitating leukemia relapse. In this study, the elevated VISTA expression in myeloid neoplasia versus lymphoid subsets was seen by analyzing of human leukemia and lymphoma cell lines. In addition, the VISTA overexpression in AML cells of nucleophosmin 1 (NPM1) mutant was confirmed in both leukemic and CD3+ cells in short first remission versus long first remission cases according to the length of first remission. By observing these results, authors conclude that the VISTA high expression on AML cells might indicate an early reaction to immune activation-mediated tension for controlling tumor progression. Furthermore, this study highlighted the VISTA as a potential target for the treatment of patients with AML either in the prevention of disease recurrence or in patients with treatment refractory disease. In a murine brain glioma model, the VISTA-deficient animal highly resisted tumor induction, and in this model, depletion of CD4+ T cells enhanced tumor formation. 110 Consistently, elevated VISTA expression is also reported in glioma tissues in Gtex when compared with normal tissues. 111 With worthiness, in a recent study conducted by Zhang et al., 112 based on VISTA and CD8+ TILs in patients with hepatocellular carcinoma (HCC), they classified TMEs into four immune subtypes as follows: VISTA+/CD8+, VISTA+/CD8−, VISTA−/CD8+, and VISTA−/CD8+. 112 In this study, patients with VISTA-positive expression in tumor cells have shown prolonged OS as compared with those with VISTA-negative expression, and the tissue dual positive for VISTA and CD8+ TILs that were measured by various tissue microarray analysis was associated with favorable TME and better OS. In patients with high-grade serous ovarian cancer, VISTA expression on immune and endothelial cells was associated with pathologic type, while conversely its expression on tumor cells was associated with prolonged OS. 113 Furthermore, the association between VISTA expression in tumor cells and a favorable prognostic significance has recently been reported in patients with pancreatic cancer. 114 Consistent with these findings, in prostate cancer patients who were treated with anti-CTLA-4 (ipilimumab), the increased expression of PD-L1 and VISTA inhibitory molecules in independent subsets of macrophages deserves attention. 115 In OSCC, VISTA expression in tumor cells and lymphocytes is associated with IL-33 levels. 116 The IL-33 biology in various cancers has been described elsewhere. 117 In fact until recently and in patients with cancer, VISTA has mostly been described as a negative checkpoint regulator that suppresses T-cell activation with subsequently poor prognosis. Hence, the novel discovery of VISTA protein expression in tumor cells of patients with HCC or patients with high-grade serous ovarian cancer has underscored the importance of its newly found expression on tumor cells that results in a favorable prognosis. This finding may be of great interest to investigate the potentially precise role of VISTA expression in tumor cells and to design a rational combination for cancer immunotherapy. Consistently, a recent study indicates that VISTA expression was promoted in tumor cells following the chemotherapy through the HIF-2α transcription factor. 118 In the same study, the VISTA-blocking antibody 13F3 has shown therapeutic enhancement to carboplatin therapy. Importantly, in a recent study of patients with RCC along with venous tumor thrombus, the increased VISTA expression on immune cells was associated with T-cell exhaustion TOX marker expression and a worse prognosis of the disease. 119 These results further indicate a VISTA inhibitor may potentially have synergistic effects when combined with chemotherapy. Given the significant therapeutic potential of VISTA targeting approaches, some VISTA-specific antagonist agents along with monoclonal antibodies with their potential suppressing VISTA activity that have recently moved into the clinical trials are presented in Table 4.
Current ongoing clinical trials for VISTA-specific monoclonal antibodies or antagonist agents.

PD-L1, programmed cell death-ligand 1; VISTA, V-domain Ig suppressor of T-cell activation.
New B7 family checkpoints molecules
The newly discovered representatives of the B7 family comprise five members which are B7-H3 (also known as CD276), B7-H4 [(also known as B7S1, B7x, or V-set domain-containing T-cell activation inhibitor 1 (Vtcn 1)], B7-H5 (also known as VISTA, platelet receptor Gi24 precursor or PD-1H), B7-H6 [NK cell cytotoxicity receptor 3 ligand 1 (NCR3LG1)], and B7-H7 [human endogenous retrovirus-H long terminal repeat-associating protein 2 (HHLA2)].120,121 The important features of B7-H5, known as VISTA, and its role in the regulation and suppression of immune reactions in cancer are shown in the previous section. Thus, in this section, apart from B7-H5, we discuss the novel features of other members of the type I membrane B7 family. In mice and humans, there is a sequence similarity between B7-H3 and the extracellular domain of PD-L1; however, in humans, there is also an alternative isoform with a tandem repeat of immunoglobulin variable (IgV) and immunoglobulin constant (IgC) domains (VCVC) that is the most frequently expressed isoform. 120 At the RNA level, the B7-H3 is generally expressed in both lymphoid and nonlymphoid cells while at the protein level, its expression is mainly found in T cells, B cells, monocytes, activated DC, and NK cells. Furthermore, its aberrant expression has been reported in various malignancies such as ovary, colorectum, liver, breast, prostate, brain, RCC, and NSCLC.120,122 The putative receptor which has been identified for B7-H3 is located on activated T cells; however, monocytes and macrophages that are influenced by various disease conditions are also recognized. 123 The B7-H3 co-stimulatory or co-inhibitory action on CD4+ and CD8+ via TCR signaling that enhances IFN-γ production to produce cellular immunity, or through the nuclear factor of activated T cells, nuclear factor kappa B (NFκB), and activator protein 1 factor that influences TCR regulation to inhibit the associated gene transcription has been reported, respectively.124,125 Since the precise B7-H3 receptors have not been identified, this controversial functionality of B7-H3 might be due to other possible binding ligands that need to be discovered. Last, but also important, the potential role of soluble B7-H3 (sB7-H3) released from cells into serum and its blockage of matrix metalloproteinase inhibitor, which consequently accumulates B7-H3 on the cell surface, should be mentioned. In this context, the high circulating serum B7-H3 levels have been associated with various malignancies in patients. 126 The association between the B7-H3 expression and the poor outcome in various human cancers further emphasized the feasibility of this immune checkpoint in the prediction and prognosis of various human cancers. For instance, B7-H3 in human BC is a direct target of microRNA (miR)-29c. 127 Nygren et al. 127 have found that the high expression of miR-29c is associated with a significantly reduced risk of death from BC. 127 Downregulation of miR-29c may play roles in BC progression through deregulating B7-H3 expression in BC. Interestingly, nearly 50 miRNAs that downregulate B7-H3 at protein levels have been identified, and among them, 13 miRNAs including miR-214, miR-363, miR-326, miR-940, miR-29c, miR-665, miR-34b, miR-708, miR-601, miR-124a, miR-380-5p, miR-885-3p, and miR-593 target B7-H3 via binding to its three prime untranslated region (3′-UTR). 127 Consistently, multiple adverse clinical symptoms have been found in association with elevated B7-H3 expression in either tumor cell or diffuse tumor vasculature in patients with ccRCC. 128 In particular, in higher grade and stage RCC, lower miR-187 expression levels are observed. 129 Downregulation of miR-187 may be relatively involved in RCC progression through its interfering influence on B7-H3 expression. In osteosarcoma cells, the expression level of B7-H3 is a direct target of miR-124, and overexpression of this tumor suppressor miRNA, inhibits cell proliferation through targeting B7-H3 in OS tumor tissue in vitro. 130 Interestingly, in recent years, miRNA profiling and sequencing revealed that its expression is dysregulated in cancer mainly via amplification or deletion of miRNA genes or transcriptional control changes. 131 Since miRNAs might act as either oncogenes or tumor suppressors in different conditions, further studies of their alterations and associated signatures can be a useful approach for tumor classification, diagnosis, prognosis, and even immunotherapeutic treatments. Among five major hematologic malignancies, the B7-H3 expression was reported to be highest in patients with AML and lowest in patients with acute lymphoblastic leukemia. 132 Importantly, analyzing B7-H3 expression via the TCGAseq and GSE10358 datasets showed that B7-H3 is associated with negative prognostic value in AML patients. Notably, positive correlation between B7-H3 expression and four genes belonging to the tumor necrosis factor family including TNFRSF4 (OX40), TNFSF9 (CD137L), TNFSF14 (LIGHT), and TNFRSF18 (GITR) was identified in patients with AML. In addition, in these patients, the VISTA (B7-H5) and CD70 genes also showed a positive correlation with B7H3 expression. Consistent with this study, other studies confirm blocking CD70 in patients with AML has effective therapeutic results. 133 Among various strategies in cancer immunotherapy, recently chimeric antigen receptor T (CAR-T) cell therapy in prostate cancer tissues and cells also deserves attention. 134 The high expression of B7-H3 on the surface of PC3, DU145, and LNCaP cells and prostate cancer tissues was efficiently inhibited in vitro and in vivo by B7-H3 CAR-T cells in an antigen-dependent manner. Furthermore, in vitro application of B7-H3 CAR-T cells into tumor cells produced high levels of IFN-γ and TNF-α that indicates B7-H3 can be a potential target for specific CAR-T cells therapy in prostate cancer. There are already studies that explain the precise role of those cytokines in the treatment of cancers. 135 Consistently, in NSCLC treatment with dihydroartemisinin (DHA), the B7H3 expression was also actively involved with the positive effect of the antitumor agent. 136 In this study, DHA treatment largely inhibited the B7-H3 overexpression while it expanded the infiltration of CD8+ T Lymphocytes in the xenografts. This result further highlights the potential effect of B7-H3 blockade in cancer immunotherapy. Another type I membrane B7 family member is the B7-H4 that contains one IgV and one IgC domains with 87% amino acid similarity between humans and mice. 120 Recent studies indicated the broad spectrum of patients’ tumors that have been associated with aberrant B7-H4 expression across a wide variety of cancers, including colorectal, craniopharyngioma, RCC, BC, prostate cancer, pancreatic cancer, esophageal squamous cell carcinoma (ESCC), and cervical cancer.137–144 Furthermore, serum sB7-H4 may be a valuable prognostic marker in patients in this wide spectrum of cancers. In patients with non-metastatic ccRCC, the high levels of sB7-H4 along with elevated PB neutrophil count were associated with either poor progression-free survival (PFS) or OS. 145 Recently, Emaldi and Nunes-Xavier 146 have found increased B7-H4 gene expression in Caki-1 and 786-O renal cancer cells that were treated either with tyrosine kinase inhibitors (axitinib, cabozantinib, and lenvatinib) or mTOR inhibitors (everolimus and temsirolimus). 146 In this study, knocking down the expression of B7-H4 by small interfering RNA (siRNA) reduced renal cancer cell viability while increasing sensitivity to drug treatment. These findings highlight the therapeutic potential of B7-H4 in immune checkpoint-targeted therapy. Although the activated T cells and myeloid-derived suppressor cells (MDSCs) express the B7-H4 putative receptor, its counter receptor has not been identified yet. The contact between B7-H4 and its associated receptors inhibits TCR-mediated T-cell proliferation and induces IL-2 production which negatively regulate T-cell responses. Consistently, in a study conducted by Xu et al., 147 the B7-H4 expression at both mRNA and protein levels was measured to be upregulated by IL-2, IFN-α, and IFN-γ in a ccRCC cell line which was obtained from patients with RCC. 147 In this study, the low efficacy of IL-2, IFN-α, and IFN-γ in metastatic RCC could be due to the pathway in which the B7-H4 escapes from the immune response. Finally, it should be considered that immunohistochemical analysis of cervical cancer tissues has also revealed a negative correlation between B7-H4 expression and IL-2 cervical cancer patients. 148 Since this research proposes that the putative B7-H4 receptor could be induced upon activation, the discovery of more specific receptors, even at highly differentiated levels, could improve our understanding of B7-H4 antitumor immunity. It should be taken into account that the B7-H4 mRNA is extensively spread in peripheral tissues but it has low or absent expression at protein level in normal tissues. Hence, its high protein expression in tumor tissues exposes the B7-H4 as a crucial target for immunotherapy. Consistently, the preclinical study with B7-H4-specific CAR-T cells or its antibody-mediated blockade has revealed promising therapeutic results in vitro and in vivo.149,150 Inconsistent with the findings mentioned above, the positive prognostic role of sB7-H4 in patients with NSCLC treated with pembrolizumab should not be neglected. 151 For example, patients with elevated levels of sB7-H4 (>63.9 pg/mL) have shown longer OS and PFS. In this context, unveiling the precise interaction of sB7-H4 with the cell protein that consequently enhances T-cell-mediated immune reaction deserves particular attention. Among the B7 family members identified until now, B7-H6 and B7-H7 are the most recently distinguished proteins. B7-H6 activates NK cells through NKp30 has two Ig domains including IgV and IgC with homologous sequences as well as the other B7 family members. 152 In contrast with the B7-H4, the B7-H6 mRNA is not broadly found in normal tissues while its high expression has been reported in various subsets of human HCC and chronic myeloid leukemia.152,153 Similarly, the expression of human B7-H6 protein is not seen in healthy tissues while its expression is elevated on various human tumors such as cervical cancer, ESCC, BC, small-cell lung cancer, and posterior lipopolysaccharide (LPS) stimulation of glioma cells.154–158 In this context, the negative prognostic value of B7-H6 in human cancers deserves also particular attention. In patients with pancreatic cancer, the B7-H6 expression on tissues has been associated with tumor progression and metastasis while notably the soluble form of it has shown the same profile in these patients. 159 Shorter OS was seen also at high expressed levels of both the B7-H6 cell surface and sB7-H6. In this study, the in vitro knocking out of B7-H6-induced NK cells increased cytokine production and increased the PC tumor cells’ interaction with NK-mediated cytotoxicity. Apart from these findings, the B7-H6 may show opposing functions based on its opposite effects on NK cells. In this context, some reports indicate that the B7-H6-expressing tumor cells can be eliminated by the immune system through cytotoxicity or secretion of various cytokines. Several approaches applied to tumor therapies including radiotherapy, chemotherapy, and immune mediator therapy using cytokines such as TNF-α, have been shown to upregulate B7-H6 expression in tumor cells and enhance their sensitivity to NK cell cytolysis. 160 Although tumor-induced B7-H6 cells stimulate innate immunity, the mechanism by which these cells escape from the immune system should not be overlooked. For example, the B7-H6 induces anti-apoptosis through signal transducer and activator of transcription 3 (STAT3) pathway activation and promotes tumor proliferation. 161 In addition, to speed up tumorigenesis, B7-H6 secretes TNF-α, IFN-γ, and B7-H6-specific bispecific T-cell engager (BiTE) triggers T. 161 In this context, the specific interaction of the B7-H6 on the surface of transformed cells with NKp30 that produces IFN-γ also deserves attention. 162 Notably, low IFN-γ levels in various cancers including H22 hepatoma and B16 melanoma induced PD-L1, PD-L2, CTLA-4, and facilitated tumor immune escape. 163 In human oral squamous carcinoma, PD-L1 surface expression is increased by IFN-γ through the protein kinase D isoform 2 (PKD2) signal pathway while inhibition of PKD2 activity prevents PD-L1 expression along with antitumor effect enhancement of tumor antigen-specific T cell. 164 It should be taken into account that both type IFN I and II regulate a multigenic PD-L1-dependent and PD-L1-independent resistance to ICB and their precise molecular mechanisms have been explained elsewhere. 165 Several factors that influence tumor cells can also regulate B7-H6 expression. For example, its surface expression can be upregulated by endoplasmic reticulum (ER) stress. Protein kinase R-like ER kinase (PERK) that phosphorylates eukaryotic initiation factor-2α (eIF2α) plays a pivotal role in B7H6 induction via ER stress. 166 Obiedat et al. 166 have revealed that nelfinavir and lopinavir enhance eIF2α phosphorylation and subsequently provoke B7-H6 expression, a condition in which the enhanced B7-H6 expression improved melanoma targets for CAR-T cells directed against this immune checkpoint. 166 Furthermore, the class I histone deacetylase inhibitors (HDACi) or siRNA-mediated knockdown of the class I histone deacetylases (HDAC) 2 or 3, downregulates the B7-H6 expression either at transcription or at translation levels, respectively. 162 Such a downregulation at the translation level reduces the NKp30-dependent effector functions of NK cells. 162 Regarding these findings and as a potential cancer treatment, combined immunotherapy with HDACi should be further investigated. Finally, regarding the therapeutic potential of bispecific antibodies targeting B7H6, recent results obtained from a novel B7-H6-targeted IgG-like T cell-engaging antibody in gastrointestinal tumors deserve attention. This study revealed that B7-H6/CD3 IgG-like T-cell engager (ITE) induced redirection of T cells into B7H6 expressing tumor cells yielded various results including proliferation of T cells along with B7-H6-dependent lysis of tumor cells. In addition, in in vitro coculture assays and in vivo colorectal cancer models, cytokine secretion and infiltration of T cells into tumor tissues were observed. 167 These results highlight the importance of additional clinical investigations in B7-H6 targeted therapy. Until now, the regulation of B7-H6 expression in tumor cells is not adequately explored. However, a broad understanding of the actual mechanisms that govern B7-H6 expression is also pivotal for the evaluation of this immune checkpoint as a crucial target in tumor therapies. Last but not least, in the B7 family, the B7-H7 which is known as HHLA2 with its associated receptor CD28H [transmembrane and immunoglobulin domain-containing protein 2 (TMIGD2) or immunoglobulin containing and proline-rich receptor-1 (IGPR1)] is only found in humans. 168 Despite B7-H7 expression in various epithelia of human organs, its expression has also been reported on human monocytes and macrophages. Although there is no expression of B7-H7 on resting T or B cells, IFN-γ or other inflammatory signals like LPS or polyinosinic:polycytidylic acid (poly I:C) upregulate B7-H7 expression on mature monocytes and dendritic cells.121,169 Furthermore, a high expression of CD28H has been reported on naïve T cells, pDCs, and NK cells. 170 Regarding the B7-H7 functionality as a co-stimulatory molecule, the B7-H7 interaction with CD28H on the NK cells activates them via selective synergy with receptors NKp46 and 2B4 that are located on the surface of NK cells. This interaction further induced pro-inflammatory cytokine secretion along with degranulation and lysis of B7-H7+ tumor cells. 171 The high expression of B7-H7 on tumor cells intensifies NK cell functions either via natural or antibody-dependent cellular cytotoxicity. 171 Hence, the antitumor activity of NK cells through the interaction between B7-H7 and its associated receptors CD28H as a potent activator of NK cells should not be neglected. Importantly, the inhibitory function of the B7-H7 on T-cell activation and proliferation via TCR and CD28 signaling pathways also deserves attention. 121 The simultaneously combined TCR and CD28 stimulation increased B7-H7 co-inhibitory action on T cells as much as PD-L1 co-inhibitory activity. 172 Accordingly, there is also a study indicating that B7-H7 prevents the proliferation of CD4+ and CD8+ T cells in the presence of TCR signaling. 121 Interestingly, the blockade of B7-H7 boosted T-cell activity and proliferation, which further emphasized the role of its blockage as a new therapeutic approach. The elevated expression level of B7-H7 in breast, lung, ccRCC, colorectal carcinoma, intrahepatic cholangiocarcinoma, and malignant glioma that is associated with poor prognosis or metastatic disease in patients has drawn the attention of the clinical significance of the B7-H7 expression in human cancer therapy.173–176 In in vivo experiments of human gallbladder cancer, the HHLA2 overexpression promoted tumor progression while its knockdown reduced the sizes of the GBC tumors. 177 Conversely, the HHLA2 ablation inhibited both TGF-β1- and long noncoding RNA H19-induced GBC progression in vitro. However, in a recent study on human ccRCC that evaluated the HHLA2 prognostic value, a positive correlation between increased HHLA2 and survival rates is reported. 178 In this context, neither costimulatory nor co-inhibitory roles of the B7-H7 and CD28H pathways in T-cell activation or for other immune cells have been fully understood. Therefore, the identification of unknown receptors on activated T cells or other immune cells that could interact with tumor-expressed B7-H7 cells should be further investigated. Finally, given the above findings, targeting the interaction between tumor-expressed B7-H7 and endothelial-expressed CD28H, which may enhance angiogenesis in TME, could be a novel therapeutic approach in antitumor immunity. Interestingly, in a recent study, the either stimulatory or inhibitory role of HHLA2 expression in colorectal cancer has been also addressed. 179 Currently ongoing clinical trials for the new B7 family checkpoint molecule-specific antagonists are presented in Table 5.
Current ongoing clinical trials for the new B7 family checkpoint molecule-specific antagonist agents.
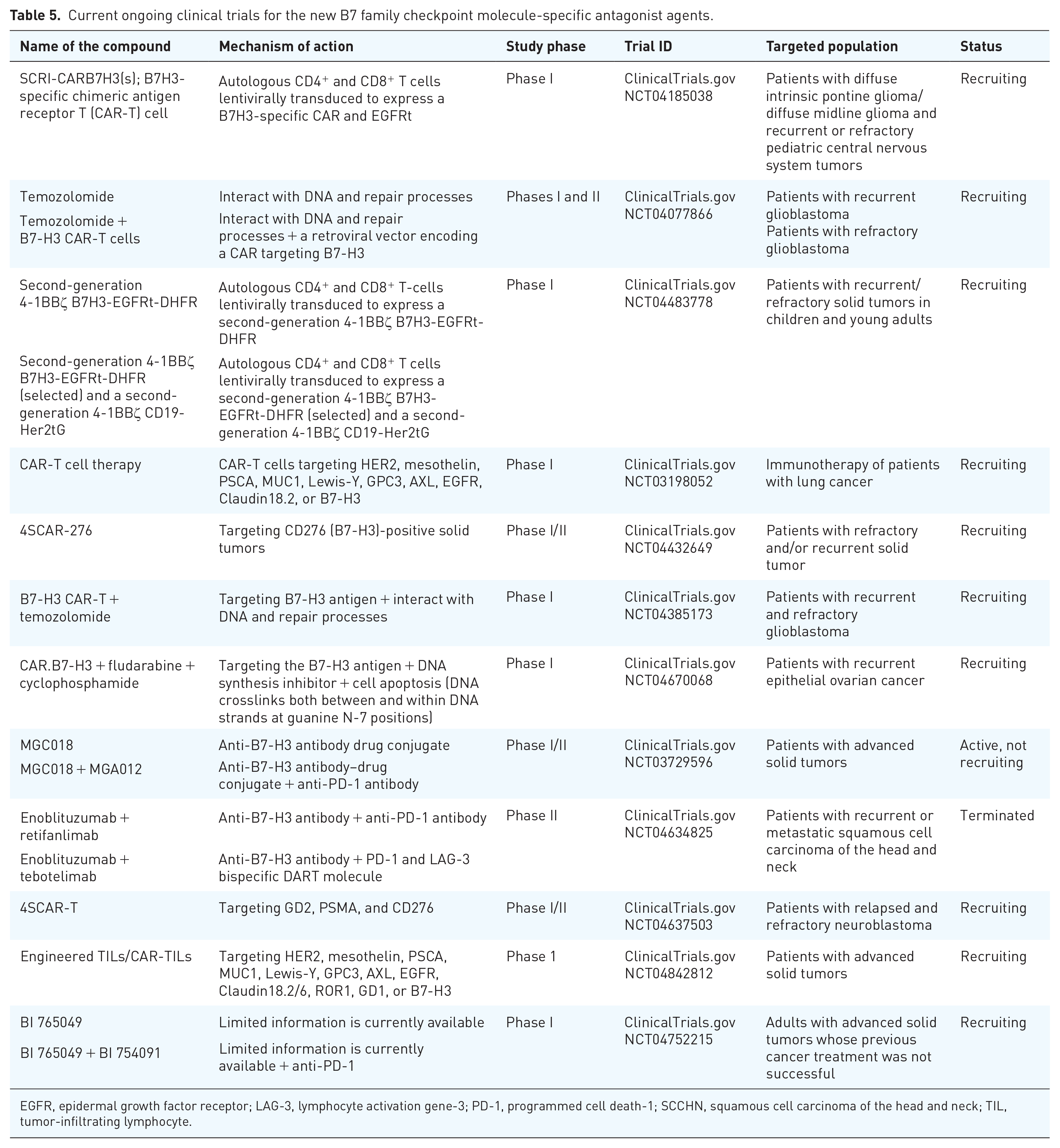
EGFR, epidermal growth factor receptor; LAG-3, lymphocyte activation gene-3; PD-1, programmed cell death-1; SCCHN, squamous cell carcinoma of the head and neck; TIL, tumor-infiltrating lymphocyte.
B-cell and T-cell lymphocyte attenuator
B- and T-cell lymphocyte attenuator (BTLA) also termed CD272 is an extracellular Ig domain-containing glycoprotein that belongs to B7/CD28 superfamily, whereas its associated ligands HVEM (herpesvirus entry mediator) which is also termed as TNFRSR14 belongs to TNF/TNFR superfamily. 180 Interestingly, the TNF superfamily that interacts with HVEM also includes two ligands such as lymphotoxin alpha (LTalpha) and LIGHT (TNFSF14). 181 The HVEM, as a complex cosignaling molecule, further interacts with the Ig superfamily BTLA and CD160. Based on the HVEM ligand engagement, the HVEM has both stimulatory and inhibitory functions. In this context, following its interaction with TNF members on T and B cells, the HVEM induces stimulatory signals, whereas its interaction with BTLA or CD160 conversely induces an inhibitory signal on T cells. 182 There is a study that indicates the interaction of LIGHT with the HVEM in lymphoid malignancy stimulates a significant elevation in the expression of IL-8 and the upregulation of apoptotic genes. 183 By contrast, the interaction of BTLA with HVEM recruits the protein tyrosine phosphatases Src-homology phosphatase type-1 (SHP-1) and Src-homology phosphatase type-2 (SHP-2) and suppresses TCR activation. 184 Notable, recruitment of SHPs to its cytoplasmic domain blocks also TCR signal transduction after ligation to CTLA-4 or PD-1. 184 Last but not least, ligation of BTLA with HVEM inhibits the secretion of cytokines including IL-2, IFN-γ, IL-4, and IL-10 and downregulates the proper immune response.185,186 Consistently, in a murine TC-1 cervical cancer model, blocking BTLA-HVEM interactions with psBTLA [eukaryotic expression plasmid that expressed the extracellular domain of murine BTLA (soluble form of BTLA)] significantly enhanced antitumor immunity of heat shock protein 70 (HSP70) vaccine by increasing the expression of Th1 cytokines, IL-2, and IFN-γ in the TME. 187 Despite the BTLA expression on T cells and B cells, its expression has been seen further on other critical regulators of pro- and antitumor immunity like NK cells and APCs.188,189 During the past decade, various studies on tumor cells and their microenvironment have revealed the irregular expression of both BTLA and HVEM in effector lymphocytes against tumor cells. For example, the tumor cells immunohistochemistry analysis of patients with stage I–III NSCLC has shown high BTLA expression which was further associated with a positively high level of PD-L1 expression, lymphatic invasion, a shorter relapse-free survival, and poor prognosis. 190 Similarly, the BTLA expression was significantly upregulated on leukemic cells and NK cells from patients with CLL, and further soluble BTLA was found to be increased in the sera of these patients and this was associated with shorter survival time. 191 Consistent with this, in patients with CLL, increased expression of BTLA on the surface of CD4+ and CD8+ T lymphocytes was correlated with poor outcomes and T-cell-mediated immune exhaustion. 192 Notably, blocking BTLA combined with bispecific anti-CD3/anti-CD19 antibody enhanced anti-leukemic responses through CD8+ T cells. In this context, the BTLA/HVEM binding disruption further stimulated IFN-γ+ CD8+ T lymphocytes secretion. The anti- or pro-tumorigenic activities of IFN-γ and its associated receptors (IFN-γ receptor 1/2) have been defined elsewhere. 193 Likewise, in gastric biopsy and PB samples that were taken from patients with GC, an increased BTLA mRNA and protein were identified in advanced stages. Interestingly, in these patients, the HVEM was higher only at the protein level. 194 These recent findings highlight that the BTLA/HVEM/sHVEM (soluble herpes virus entry mediator) inhibitory pathways may serve as potential therapeutic strategies in patients with GC. In an animal model study of epithelial ovarian carcinoma, the combined chemotherapy along with anti-BTLA antibody significantly decreased peritoneal tumor volume and prolonged survival along with increased activated CD4+ and C8+ T lymphocytes in affected mice. 195 In this study, the BTLA expression was identified predominantly on B lymphocytes, particularly on CD19hi B cells, and the elevated expression might be due to the increased secretion of anti-inflammatory cytokines including IL-6 and IL-10 through phosphorylation of STAT3 and serine/threonine kinase (AKT) in B lymphocytes. Notably, the influence of increased anti-inflammatory cytokines like IL-6, IL-10, and TGF-β on tumor progression in ovarian cancer has been reported elsewhere. 196 The synergistic therapeutic value of anti-PD-1 and anti-BTLA against murine glioblastoma also has been reported. 197 A combined anti-PD-1 and anti-BTLA therapy for tumor-bearing mice demonstrated increased OS (up to 60%) compared either with anti-PD-1 (20%) or anti-BTLA (0%) individually (p = 0.003). Furthermore, in this preclinical study, an increased expression of CD4+ IFN-γ (p < 0.0001) and CD8+ IFN-γ (p = 0.0365) was seen along with decreased levels of CD4+ FoxP3+ Treg in the brain (p = 0.0136) on this combined therapy. Another recent study of patients with cutaneous melanoma [skin cutaneous melanoma (SKCM)] has shown that BTLA expression levels increased in metastatic melanoma compared to normal skin tissues and primary melanoma. In this study, increased BTLA expression correlates with improved prognosis in SKCM and was positively associated with enhanced immune cell responses. 198 Notably, the accurate BTLA anticipation in the outcome of melanoma patients that were treated with melanoma-associated antigen 3 (MAGE-A3) blocker or first-line anti-PD-1 further highlights the pivotal role of BTLA as a predictive biomarker in melanoma. Although this study did not support an inhibitory role of BTLA for immune escape and cytotoxic activity of effector lymphocytes, its expression as a single gene and independent good prognostic factor should not be overlooked and deserves to be systematically investigated in vivo and in vitro in other metastatic cancers. In this context, there is a recent study that revealed the tumor cell-intrinsic BTLA receptor could prevent tumor cell proliferation through the protein kinase (ERK1/2, extracellular signal-regulated protein kinase) signaling pathway. 199 Importantly, the interaction of tumor cell-intrinsic BTLA and HVEM prevented extracellular regulation of protein kinase (ERK1/2) and consequently resulted in the inhibition of tumor cell proliferation. This study further highlighted that tumor cell-intrinsic BTLA/HVEM inhibition can be a potential target in cancer immunotherapy. Table 6 summarizes the characteristics of anti-BTLA-specific monoclonal antibodies currently under clinical trials.
Current ongoing clinical trials for BTLA-specific monoclonal antibodies or antagonist agents.

BTLA, B- and T-cell lymphocyte attenuator.
Conclusions and future perspectives
In recent years, cancer immunotherapy and particularly immune checkpoints inhibitors (ICIs) including blocking CTLA-4/CD28 or PD-L1/PD-1 axis are being investigated in various preclinical experiments or clinical studies of human malignancies at various stages of disease. Despite some of their therapeutic success, a large proportion of patients indicates inadequate response to ICI due to the adaptive, primary or acquired resistance against immunotherapy. In TME, the PD-L1 overexpression in tumor cells is considered one of the crucial mechanisms involved in tumor immune escape and several factors, including cellular or intracellular oncogenic signals, transcriptional factors, and posttranscriptional modifiers, could modulate its over expression. The soluble PD-L1 (sPD-L1) can also effectively predict metastasis and poor prognosis in various cancers through recruiting the suppressive immune cells into the affected sites. Hence, to increase the effectiveness of the ICI, combined CTLA-4 and PD-1 blockers like ipilimumab and nivolumab have been examined. Furthermore, the anti-PD-1/PD-L1 antibodies (pembrolizumab and nivolumab/atezolizumab and durvalumab) as the most broadly prescribed anticancer therapies have been also investigated in various clinical trials. However, both approaches indicate some degree of limiting immune-related adverse effects along with induction of autoimmunity in treated patients. Importantly, inhibition of some B7:CD28 pathways, such as PD-1 inhibitor, has been found to induce the expression of other immune checkpoints that consequently weakens the influence of the ICI. Based on these observations, there is a need to discover novel targets or ligands that enhance ICI for various checkpoints, along with the feasibility of their combinational therapy in each individual cancer. In this context, those immune checkpoints that were discussed in this review and summarized in Figure 1, such as Tim-3, LAG-3, TIGIT, VISTA, new B7 family proteins, and BTLA are currently investigating to unveil either their potential costimulatory or coinhibitory characteristics as biomarkers in the course of immunotherapy and for the advancement of targeted agents. Furthermore, the complete picture of the checkpoint biology on the surface of various immune cells rather than T cells that are also infiltrating into TME, including macrophages, B cells, neutrophils, DCs, NK cells, cancer-associated fibroblasts, and MDSCs, may open new horizons for the next generation of immunotherapies. Undoubtedly, the discovery of the precise molecular pathways regulated by each of them and their explicit roles in immune homeostasis, along with the identification of their novel ligands could support the innovative approaches to immunotherapy. Finally, due to the plasticity of tumors and to enhance the curative effect of ICIs, additional factors including TME composition, tumor mutation burden, and microsatellite instability, together with the individual properties of the patient’s immune response, should be taken into account for the development of future sequential therapeutic strategies ICIs.

A summarized overview of mechanisms of action of various immune checkpoints and immune cell components. Signals induced by each immune checkpoint and the interaction via immune mediators are schematically depicted.