
Abstract
Narcolepsy type 1 (NT1) and, to a lesser extent, neurodegenerative diseases, such as Alzheimer’s disease, Parkinson’s disease, dementia with Lewy bodies and multiple system atrophy, entail the loss of the hypothalamic neurons that release the hypocretin/orexin (H/O) neuropeptides. NT1 has been associated with autonomic anomalies including alterations in temperature regulation and cardiovascular control, particularly during sleep. A spectrum of autonomic dysfunctions also characterizes neurodegenerative diseases. The central autonomic network (CAN) is an interconnected set of brain structures that are critical for the control of autonomic preganglionic neurons. The H/O neurons include pre-autonomic neurons that directly target preganglionic sympathetic neurons in the intermediolateral column of the spinal cord and parasympathetic neurons in the dorsal motor nucleus of the vagus nerve. The H/O neurons also project to and modulate the activity of other CAN structures that include pre-autonomic neurons, such as the rostral ventromedial medulla and caudal raphe nuclei, the rostral ventrolateral medulla and the hypothalamic paraventricular nucleus. In addition, the H/O neurons project to and modulate the activity of neurons in the nucleus of the solitary tract in the medulla, which receives and relays visceral afferent information, and in higher order structures of the CAN, such as the dorsomedial nucleus of the hypothalamus and the extended amygdala. The H/O neurons should, therefore, be regarded as a key component of the CAN. Functional alterations of the CAN due to H/O neuron deficiency might contribute to autonomic anomalies in patients with neurodegenerative diseases and are likely to underlie autonomic anomalies in patients with NT1.
Keywords
Introduction
Orexin A and B (O-A and O-B, also called hypocretin 1 and 2, H-1 and H-2, respectively) are two hypothalamic neuropeptides independently discovered in 1998 by two different research groups, which employed hypothesis-free molecular biology techniques on rats. 1,2 These neuropeptides are synthesized and released by neurons in a restricted area of the tuberal region of the hypothalamus, caudal to the paraventricular nucleus (PVN). 3 The loss of hypocretin/orexin (H/O) neurons has been associated with the pathophysiology of narcolepsy with cataplexy, 4 which is presently referred to as narcolepsy type 1 (NT1). 5 Less severe losses of H/O neurons occur in neurodegenerative diseases including Alzheimer’s disease (AD), 6 Parkinson’s disease (PD), 7 dementia with Lewy bodies (DLB) 8 and multiple system atrophy (MSA). 9 To a different extent, NT1, 10,11 AD, 12 PD 13 and DLB 13 have all been associated with alterations of the autonomic nervous system. The autonomic nervous system is controlled in the context of integrated behavioural responses by the central autonomic network (CAN), an interconnected set of brain structures that are also critical for the control of neuroendocrine, respiratory and sphincter motor neurons. 14 The aim of this narrative review is to discuss and put in a translational context the neuroanatomical and neurophysiological basic research that links the H/O neurons to the CAN.
The H/O neurons
The H/O neurons project to 3 and receive synaptic inputs from 15 a wide array of central nervous system structures. 3 The electrical activity of the H/O neurons is mainly restricted to active wakefulness, although sparse activity bursts also occur during the so-called phasic periods of rapid-eye-movement (REM) sleep that entail bursts of eye movements and muscle twitches. 16 However, direct measurements with brain microdialysis in cats 17 and rats 18 have shown that the interstitial H-1/O-A concentration depends loosely, if at all, on the states of sleep and wakefulness in these species. In humans, H-1/O-A concentration in the cerebrospinal fluid (CSF) obtained by spinal tap is higher during the night than during the day. 19 Thus, for reasons that are still unclear, there seems to be a temporal mismatch between the electrical activity of the H/O neurons and the brain interstitial concentration of H-1/O-A. Corresponding data are lacking for H-2/O-B concentration. Further complexity is added by evidence that H/O neurons co-release dynorphin and glutamate, and potentially also nociceptin/orphanin FQ, galanin, neurotensin and gamma-aminobutyric acid (GABA), together with the H/O neuropeptides. 20 H-1/O-A binds either H/O receptor 1 or H/O receptor 2, whereas H-2/O-B is selective for H/O receptor 2. 2
H/O neuron loss and autonomic dysfunction in NT1
The brain of patients with NT1 shows a generalized absence of H/O neuropeptides 4 as well as the loss of other markers of the H/O neurons, such as dynorphin. 21 This indicates the functional loss of the whole H/O neurons, and not just of the H/O neuropeptides, possibly due to an autoimmune reaction. 22 Accordingly, present guidelines for NT1 diagnosis indicate that a CSF concentration of H-1/O-A lower than one-third that expected in healthy subjects may substitute for findings of cataplexy, short sleep latency and sleep-onset periods of REM sleep. 5
NT1 has been associated with autonomic anomalies including abnormal pupillary reaction during darkness, impaired nocturnal penile tumescence and alterations in temperature regulation and cardiovascular control. 11 Concerning cardiovascular control, NT1 may entail a blunted fall of arterial pressure from wakefulness to sleep, and particularly to REM sleep, coupled to a variable decrease in arterial pressure during wakefulness (Figure 1), and a tendency towards an increase in heart rate, particularly during sleep. 10 A substantial variability has been noted in the occurrence of these alterations, pointing to the existence of as-yet unknown biological modifiers of the effects of H/O deficiency on cardiovascular control. 10 Nonetheless, recent data indicate that middle-aged and elderly patients with narcolepsy had more health contacts than control patients with respect to several disease domains, such as diabetes, cardiovascular and gastrointestinal disorders, 23 which may result, at least in part, from altered autonomic control. Moreover, higher values of diastolic pressure and heart rate were recently found in patients with NT1 treated with psychostimulants than in untreated patients, suggesting an increased long-term risk of cardiovascular diseases. 24
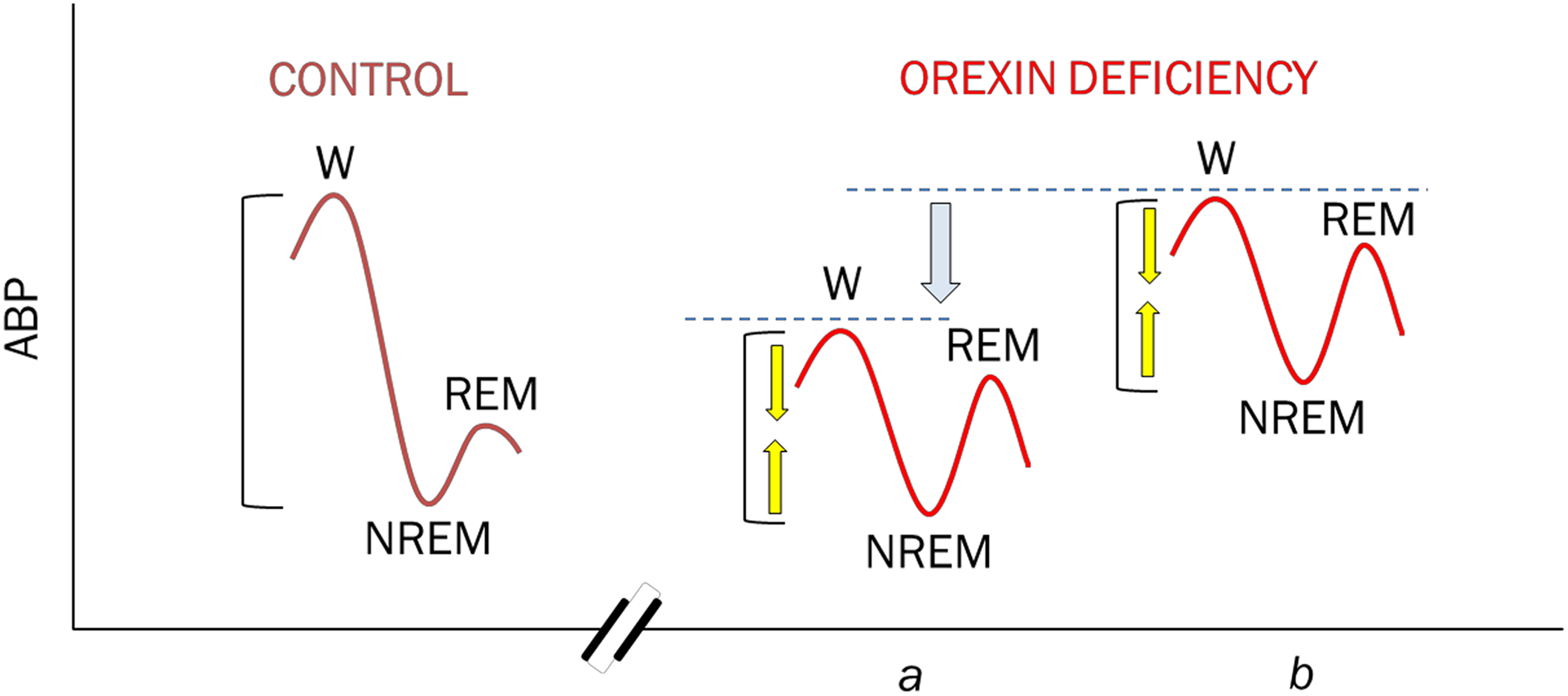
Diagram showing the alterations of the control of ABP, which have been associated with H/O deficiency. In control subjects (left), ABP physiologically decreases from wakefulness (W) to NREM sleep and rises again during REM sleep. Hypocretin/orexin deficiency (orexin, right) has been found to entail a variable decrease in ABP during W (single arrow) coupled to a blunted decrease in ABP from W to NREM and (particularly) to REM (double arrows). H/O deficiency may thus be associated with ABP values that are lower than normal in W and normal during sleep (a) or normal in W and higher than normal during sleep (b). Reproduced from the study by Berteotti and Silvani 10 with permission. ABP: arterial blood pressure; NREM: non-rapid eye movement; REM: rapid eye movement.
H/O neuron loss and autonomic dysfunction in neurodegenerative diseases
AD is characterized by brain amyloid plaques, tau protein deposits forming neurofibrillary tangles and neurodegeneration. 25 Neurodegeneration in AD may also involve the H/O neurons, whose number was found decreased by 40% in post-mortem brain analyses. 6 However, this decrease is either inconstant or associated with increased activity of the remaining H/O neurons, as the CSF levels of H-1/O-A in patients with AD have been found only slightly decreased 6 or even increased. 26 This may be of pathophysiological relevance because of preclinical evidence that H/O signalling increases amyloid neuropathology in AD models. 27 The occurrence of autonomic dysfunction in patients with AD has received relatively little attention. Nonetheless, one study reported mild-to-severe impairments in the blood pressure response to the Valsalva manoeuvre and in the heart rate response to isometric contraction in patients with mild-to-moderate AD, suggesting sympathetic dysfunction. 28 Accordingly, the sympathetic response to the stress of standing was also found impaired in patients with AD. 12
PD and DLB are characterized by neurodegeneration and α-synuclein deposits forming neuronal Lewy bodies and Lewy neurites. 13 Neurodegeneration in PD and DLB was found to entail a massive loss of H/O neurons, which reached a 62% cell loss in Braak stage V of PD, 7 and approximately a 75% cell loss in DLB. 8 The number of H/O neurons was also found reduced to approximately one-third in patients with MSA, a synucleinopathy characterized by neuroglial rather than neuronal cytoplasmic inclusions. 13 Nonetheless, the H/O neurons loss in patients with PD, DLB and MSA is generally insufficient to entail significant decreases in CSF levels of H-1/O-A. 29 –31
There is a lack of information on the involvement of the H/O system in pure autonomic failure, a related neurodegenerative disorder that entails peripheral accumulation of synuclein as neuronal cytoplasmic inclusions. 13 The synucleinopathies entail a spectrum of autonomic dysfunctions including orthostatic hypotension, thermoregulatory dysfunction, gastrointestinal dysmotility and urogenital dysfunction. The contribution of central versus peripheral neurodegeneration to these dysfunctions is thought to be the highest for MSA, intermediate for PD and DLB and the lowest for pure autonomic failure. 13
The CAN
The CAN includes the pre-sympathetic neurons (S0N) that project directly to the sympathetic preganglionic neurons (S1N) in the intermediolateral cell group (IML) of the spinal cord (Figure 2). Most of these S0N are located in the rostral ventromedial medulla (RVMM), the caudal raphe nuclei (raphe obscurus and pallidus), the rostral ventrolateral medulla (RVLM), the A5 area of the pons, the hypothalamic PVN and also the lateral hypothalamic area, which includes H/O neurons. 32–34 Comparatively less is known on the pre-parasympathetic neurons (P0N) that send direct projections to the parasympathetic preganglionic neurons (P1N) in the nucleus ambiguous (NAmb) and the dorsal motor nucleus of the vagus nerve (DMNX) of the medulla and to the P1N in the laminae V-VII of the sacral spinal cord (Figure 2). The NAmb and DMNX provide the bulk of parasympathetic innervation to organs located above and below the diaphragm, respectively, whereas sacral P1N regulate lower gastrointestinal and urogenital functions. 35 Some of the P0N are localized in the PVN, the RVLM and the lateral and ventrolateral parts of the midbrain periaqueductal grey (PAG) matter. 36 –39 As it will be discussed in the following sections, there is evidence that P0N are also localized in the lateral hypothalamic area, which includes the H/O neurons.
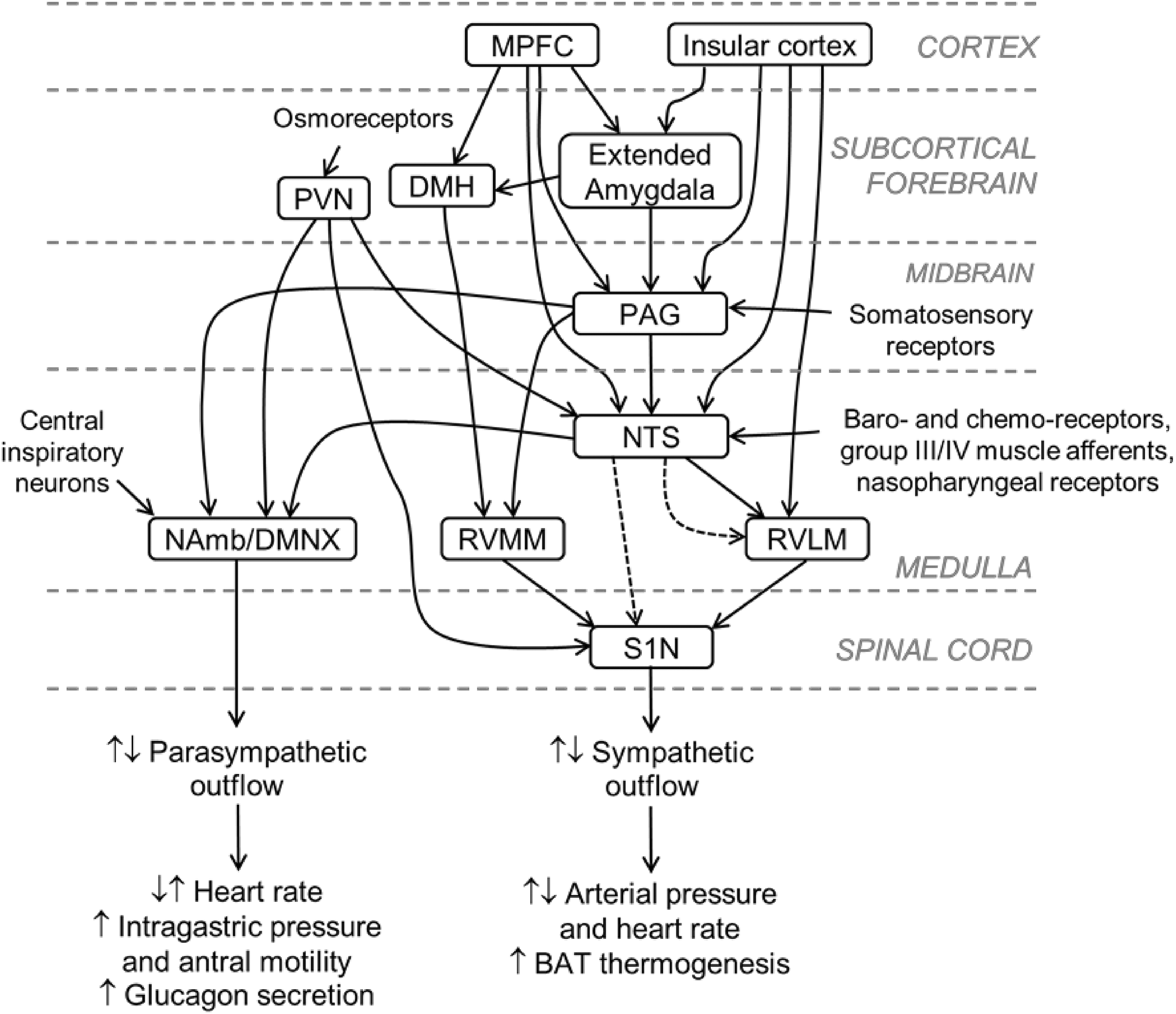
Diagram showing the major central structures regulating autonomic outflow that receive projections from H/O neurons. No distinction is made between excitatory and inhibitory connections; dashed lines indicate indirect connections. The A5 area, the parabrachial nucleus of the pons and the sacral parasympathetic preganglionic neurons are omitted for the sake of clarity. Functional studies highlighted the potential for complex bidirectional effects (increases: ↑ or decreases: ↓) of H/O neurons on sympathetic and parasympathetic outflows (cf. the text for details). Only the key functional outcomes highlighted in the text are reported here. Reproduced with modification from the study by Silvani et al. 40 with permission. H/O: hypocretin/orexin; DMH: dorsomedial nucleus of the hypothalamus; DMNX: dorsal motor nucleus of the vagus nerve; MPFC: medial prefrontal cortex; NAmb: nucleus ambiguous; NTS: nucleus of the tractus solitarius (nucleus of the solitary tract); PAG: periaqueductal grey; PVN: hypothalamic paraventricular nucleus; RVLM: rostral ventrolateral medulla; RVMM: rostral ventromedial medulla; S1N: sympathetic preganglionic neurons.
The S0N and P0N are themselves the target of synaptic projections from higher order structures of the CAN, which include the dorsomedial nucleus of the hypothalamus (DMH), the central nucleus of the amygdala, the bed nucleus of the stria terminalis (BNST), the medial prefrontal cortex (MPFC) and the insular cortex (Figure 2). 14,34,40 In addition to the S0N and P0N and their higher order controlling structures, the CAN includes the nucleus of the solitary tract (NTS) of the medulla, which is the main gateway for visceral afferents to the brain, and the parabrachial nucleus (PBN) of the pons, which contributes to relay the information from the NTS to the other structures of the CAN (Figure 2). 14,34
The H/O neurons include S0N
The H/O neurons have been reported to send long projections to all levels and all laminae of the spinal cord in mice, rats and human subjects, with a robust innervation of the IML in the thoracic and lumbar spinal cord, where S1N are localized. 41 A later study on rats confirmed H/O projections to the IML of spinal segments T1-L3, but found a progressive rostro-caudal decrease in the content of H-1/O-A and H-2/O-B measured with a radioimmunoassay on spinal cord tissue. 42 A more recent study on rats again confirmed H/O projections to S1N in the IML of the spinal segments T1 and T2, which are mainly involved in the control of the pupil size through the superior cervical ganglion, but reported that H/O projections were rare to segments T4-L2. 43
From a functional perspective, intrathecal administration of either H-1/O-A or, to a lesser extent, H-2/O-B was found to increase arterial pressure and heart rate by enhancing sympathetic activity on α1 and β1 adrenergic receptors. 44 Patch clamp recordings on spinal cord slices demonstrated that antidromically identified S1N could be directly activated by H-1/O-A or H-2/O-B. 44 This finding was later confirmed and expanded by the demonstration that the majority of H/O-sensitive S1N in the spinal cord express mRNA for both H/O receptors 1 and 2. 45 However, a more recent study that performed triple immunolabelling on rat spinal cords reported that most S1N express H/O receptor 1, but not H/O receptor 2. 46
Taken together, these results indicate that H/O neurons include S0N that send direct projections to the S1N in the IML of the spinal cord. The H/O neurons are, therefore, a source of the descending projections to S1N previously recognized to originate from the lateral hypothalamic area (Figure 2). 34 While studies agree that H/O projections target S1N in the upper thoracic segments of the spinal cord, the extent to which H/O projections target more caudal segments may be lower, and is still debated. The available evidence also indicates that H/O neurons control S1N at least in part by releasing H/O neuropeptides, as opposed to their co-transmitters. The evidence suggests that H-1/O-A binds to H/O receptor 1 in the spinal cord S1N, whereas the functional role of H-2/O-B and the expression of H/O receptor 2 on these S1N are less clear.
The H/O neurons include P0N
H/O projections to the NAmb
Fibres immunopositive to H-1/O-A were reported in the NAmb together with the expression of H/O receptor 1 within the neuropil, suggesting H/O modulation of presynaptic axonal terminals rather than of the NAmb neurons themselves. 47 This may explain the discrepancy with the previous report of no detectable expression of mRNA for H/O receptor 1 in the rat NAmb. 48 Accordingly, microinjection of H-1/O-A in the rat NAmb was found to cause vagally mediated bradycardia and to potentiate the bradycardia driven by the baroreceptor reflex in response to a rise in arterial pressure. 47 These results were later confirmed and expanded by the observation of their enhancement by oestrogen in female rats. 49
The potential for H/O projections to modulate P1N in the NAmb was also supported by patch-clamp recordings on brainstem slices from rats during early postnatal development. In particular, application of H-1/O-A to the bath of slices from rats at postnatal days 2–7 enhanced GABAergic and glycinergic inhibitory postsynaptic currents on P1N. 50–52 This effect was mediated by H/O receptor 1 51,52 and confirmed on a similar preparation by a different group. 53 Moreover, bath application of H-1/O-A was found to inhibit glutamatergic excitatory postsynaptic currents on the NAmb P1N. 50 However, focal (as opposed to bath) application of H-1/O-A with a micropipette in the vicinity of NAmb P1N inhibited the GABAergic inhibitory postsynaptic currents in slices from rats at postnatal days 20 or 30, but not from rats at postnatal day 5. 52
Taken together, these results indicate that H-1/O-A may decrease the activity of NAmb P1N by activating GABAergic and glycinergic neurons and/or by inhibiting glutamatergic neurons in other brainstem structures that project to the NAmb. H-1/O-A may also increase the activity of NAmb P1N by inhibiting GABA release from presynaptic terminals. The mechanisms whereby H-1/O-A, which is strongly neuroexcitatory, 54 would inhibit glutamatergic excitatory postsynaptic currents 50 or presynaptic GABA release 52 have not been clarified. Nevertheless, it seems fair to conclude that H-1/O-A has the potential for indirect, bidirectional effects on the P1N of the NAmb (Figure 2).
H/O projections to the DMNX
The DMNX is targeted by a moderate number of H/O projection fibres. 55 Evidence obtained in different laboratories indicates that H-1/O-A and H-2/O-B directly excite P1N in the DMNX, 56–58 likely acting on both H/O receptors 1 and 2. 59 The H/O neurons, therefore, include P0N involved in the control of P1N of the DMNX (Figure 2). However, and somewhat paradoxically, H/O projections also enhance inhibitory inputs to the P1N of the DMNX that control the stomach. 57,58 In this respect, relatively few of the DMNX P1N that project to the antrum/pylorus, duodenum or caecum were reported as H/O responsive. 57 H/O projections may thus exert complex, bidirectional effects on the vagal control of gastric function, particularly at the level of the gastric fundus or corpus. Nonetheless, microinjection of either H-1/O-A or H-2/O-B in the DMNX was reported to elicit only increases in intragastric pressure and antral motility. 59
Evidence obtained with trans-synaptic tracers indicates that the P1N of the DMNX that innervate the pancreas also receive synaptic projections from H/O neurons. 60 This innervation may be excitatory and functionally relevant, as microinjection of H-1/O-A in the DMNX dramatically increased vagal pancreatic nerve firing, whereas microinjection of an antagonist of H/O receptor 1 in the DMNX or chemical ablation of the lateral hypothalamic area, which includes H/O neurons, inhibited the increase in vagal pancreatic nerve firing elicited by insulin-induced hypoglycaemia. 61 Accordingly, microinjection of an antagonist of H/O receptor 1 in the DMNX prior to acute insulin administration decreased plasma glucose and suppressed glucagon secretion in rats. 62
Although the DMNX P1N mainly target subdiaphragmatic structures, a subset of DMNX P1N with non-myelinated C-fibre axons target the heart and may regulate coronary blood flow and cardiac contractility to some extent. 40 Recent evidence indicates that these neurons are also directly targeted by H/O neuron projections, which may be excitatory or inhibitory depending on whether the H/O co-transmitter released is glutamate or GABA. 63
H/O projections to sacral P1N
H/O projections to the sacral spinal cord have been described by two different reports, 41,55 suggesting a role of H/O peptides also in the control of sacral P1N. Electrophysiological and functional evidence of this role is lacking, however, and would be a relevant target of investigation.
The H/O neurons project to multiple CAN structures that include S0N and P0N
H/O projections to the RVMM and the caudal raphe nuclei
The RVMM and the caudal raphe nuclei include more S0N than any other brain region (Figure 2). These S0N mostly contain serotonin and are activated during arousal and stress, whereas they do not generate cardiac responses to baroreceptor or chemoreceptor inputs. 33,40 The S0N in the raphe pallidus are involved in the control of heat loss and brown adipose tissue thermogenesis. 64 The S0N in the raphe obscurus may play a role in the differential changes in regional sympathetic nerve activity (i.e. increases to skeletal muscles and decreases to kidneys) that occur during REM sleep. 65,66 These diverse functions may be modulated by H/O neurons through their projections to the RVMM, 3,67,68 the raphe pallidus 3,68,69 and the raphe obscurus, 3,68 often with direct contacts with the S0N. 68,69 There is evidence for expression of H/O receptor 1 in the RVMM, 67 and at least of its mRNA in the raphe obscurus. 48 Accordingly, H-1/O-A microinjection in the RVMM of rats was found to increase heart rate by increasing sympathetic activity and, at the same time, decreasing parasympathetic activity to the heart. 67 Nanoinjection of H-1/O-A in the rostral raphe pallidus was reported to increase sympathetic activity to and thermogenesis by the brown adipose tissue. 69 This evidence indicates that the H/O projections to the RVMM and raphe pallidus may be functionally relevant. Corresponding functional evidence is lacking for H/O action on the raphe obscurus and would be an interesting topic of research, also considering the alterations in cardiovascular control often reported during REM sleep in patients with NT1. 10
H/O projections to the RVLM
The S0N in the RVLM are mainly glutamatergic and adrenergic neurons of the C1 cell group (Figure 2). These S0N play a key role in the expression of cardiovascular reflex responses to baroreceptor, chemoreceptor and other inputs. 40 Recent evidence indicates that C1 cells of the RVLM also play important roles in non-homeostatic, adaptive responses to stress. 70 In addition, some C1 cells of the RVLM are P0N that control P1N in the DMNX. 38
H/O neurons project to the RVLM. 3,71 Many putative C1 neurons of the RVLM express H/O receptors 1 and 2 72 and are in close proximity to H/O synaptic terminals. 71,72 On the other hand, C1 cells of the RVLM may also synapse with, and presumably excite, H/O neurons. 73
Several lines of evidence suggest the functional relevance of these connections between H/O neurons and the RVLM. Microinjection of H-1/O-A in the rat RVLM was reported to increase arterial pressure, heart rate and splanchnic sympathetic nerve activity. 71,72,74 Microinjection of an antagonist of H/O receptor 1 in the RVLM decreased these responses to H-1/O-A partially, indicating that they were mediated by both H/O receptor types. 72 A similar conclusion was reached based on in- vitro recordings of RVLM neurons, suggesting, however, that H/O receptor 2 played the greater role. 75 Accordingly, H/O receptor 2 in the RVLM was reported to mediate the activation of adrenal nerve activity caused by H/O release in response to neuroglucoprivation. 76,77 Moreover, there is evidence that in the RVLM, signalling mediated by H/O receptor 2 contributes to hypertension in spontaneously hypertensive rats, a genetic model of essential hypertension. 78
Experiments on brainstem slices indicated that H-1/O-A increased the frequency of spontaneous excitatory and inhibitory postsynaptic potentials in RVLM neurons, suggesting that H-1/O-A may also modulate transmitter release from presynaptic terminals in the RVLM. 79 This may contribute to explain why microinjection of H-1/O-A in the RVLM could exert bidirectional effects on cardiovascular reflex responses in anesthetized rats, increasing baroreflex sensitivity and sympathetic activity in conditions of hypoxia and decreasing the somato-sympathetic reflex and sympathetic activity in conditions of hypercapnia. 72
H/O projections to the A5 noradrenergic cell group of the pons
The S0N in the A5 area synthesize noradrenaline, give rise to one of the most extensive projections to the spinal cord IML and may contribute to cardiovascular and cardiorespiratory integration, although their precise functional role is still unclear. 34
The A5 area receives dense descending H/O projections, 3 which, however, may be largely fibres of passage. 80 Nonetheless, at least H-1/O-A has the potential to modulate A5 neurons, as expression of H/O receptor 1 was documented in the A5 cell group at the mRNA 48 and protein levels. 81 Functional studies are needed to understand whether H/O neuropeptides actually play a role in modulating A5 cells.
H/O projections to the PVN
The PVN is a master controller of the autonomic nervous system, which integrates neuroendocrine, homeostatic and stress responses. The S1N in the PVN, many of which contain oxytocin, corticotropin-releasing factor, vasopressin or angiotensin, may be essential for generating cardiac sympathetic responses to changes in blood volume or plasma osmolality. 40
A high H/O fibre density was reported in the PVN with immunohistochemistry, 3,55 and H/O content in the PVN was confirmed by radioimmunoassay. 82 The PVN was found to express H/O receptor 2 at mRNA 48 and protein 83 levels. Accordingly, electrophysiological evidence indicates that H-2/O-B, which is selective for H/O receptor 2, 2 depolarizes PVN neurons. 84 However, the expression of H/O receptor 1 was also reported in the PVN, 85 including on spinally-projecting PVN neurons. 86 Overexpression of H/O receptor 1 in the PVN of a rat model of obesity was inferred to play a causal role in increasing renal sympathetic nerve activity in that model. 86 Microinjection of H-1/O-A into the rat PVN was recently found to increase splanchnic and renal sympathetic nerve activity acting, at least in part, on H/O receptor 1. 87 On the other hand, optogenetic experiments indicate that H/O neurons target spinally projecting PVN neurons by co-releasing glutamate or GABA with H/O neuropeptides, which facilitate glutamatergic transmission at this level. 88 Thus, functional evidence supports the view that H/O neurons may exert a complex modulation of PVN S0N, at least in part through H/O receptor 1. Conversely, the role of H/O projections, if any, in modulating the P0N of the PVN that target the DMNX 36,39 is unknown and warrants further investigation (Figure 2).
H/O projections to the PAG
The lateral and ventrolateral PAG projects to the DMNX and NAmb. 40 A high density of H/O fibres, 3 high H/O neuropeptide content 82 and moderate expression of mRNA for both H/O receptors 1 and 2 48 were reported in the PAG. The ventrolateral and, to a much lower extent, the lateral PAG also send projections back to H/O neurons. 15 Non-ingestive effects of H/O neuropeptides resemble in many respects the effects mediated via the lateral and dorsolateral columns of the PAG, which consist in somatomotor activation, vigilance, tachycardia, hypertension and non-opioid analgesia. 89 In light of the multiple connections of the PAG with other CAN structures, it is therefore possible, but unproven, that the PAG is involved in H/O neuron modulation of parasympathetic as well as sympathetic activity (Figure 2).
The H/O neurons target CAN structures that receive and relay visceral afferent information
H/O projections to the NTS
The NTS relays visceral afferent information to the CAN through two parallel pathways: one targets the PBN and a number of other circumscribed brain regions and the other starts from the PBN, and has projections that mirror those from the NTS. 34
A high density of H/O fibres was found in the NTS, 3,55,90,91 which represent, in part, collaterals of projections to the NAmb. 92 The NTS also showed moderate H/O neuropeptide content, 82 moderate expression of mRNA for H/O receptor 1 and low expression of mRNA for H/O receptor 2. 48 Accordingly, either H-1/O-A 93,94 or H-2/O-B 91 was found to directly depolarize NTS neurons, 93,94 and H-2/O-B was also found to enhance excitatory synaptic activity on NTS neurons. 91 However, H-1/O-A microinjection in different NTS regions caused either pressor/tachycardic or depressor/bradycardic responses, the latter partially due to increased cardiac vagal activity, and potentiated the bradycardia to activation of arterial baroreceptors. 90 A potential explanation for these bidirectional effects is that H/O projections to the NTS may co-release H-1/O-A and leucine-enkephalin, which would enhance the depressor and bradycardic responses elicited by H-1/O-A at this level. 95 Another explanation points to a link with different H/O doses and mechanisms of action. 96 In particular, lower H/O doses in the NTS decreased heart rate and arterial pressure due to the activation of the soluble guanylate cyclase signalling pathway by the nitric oxide produced by neuronal nitric oxide synthase. Higher H/O doses in the NTS increased heart rate and arterial pressure by activating GABAergic or enhancing nitrergic neurotransmission. 96 These complexities notwithstanding, the available evidence strongly supports a modulatory role of H/O projection on the NTS (Figure 2).
H/O projections to the PBN
Descending H/O fibres have been reported to the PBN, 3 including the Kolliker-Fuse subnucleus, 97 and the lateral PBN projects back to the H/O neurons. 15 However, no significant expression of mRNA for H/O receptor 1 and just a slight expression of mRNA for H/O receptor 2 above background were reported in the PBN. 48 Functional evidence on the role of H/O projections to the PBN is still lacking.
H/O projections to higher order structures of the CAN
H/O projections to the DMH
The DMH receives input from the amygdala and the insular cortex, projects to the caudal midline raphe and the RVMM and is critical for the expression of the cardiovascular response to stress (Figure 2). 40 The DMH receives abundant H/O projections 3 and includes neurons that project back to the H/O neurons, relaying information from the suprachiasmatic nucleus. 55 The DMH also includes some H/O neurons, 3 which contribute projections to the NTS/DMNX region of the dorsal medulla. 98 The DMH has a moderate expression of mRNA for H/O receptor 2 and a low-density expression of mRNA for H/O receptor 1. 48 Accordingly, immunohistochemical studies reported expression of both H/O receptor 1 85 and H/O receptor 2 83 in the DMH, and a recent functional study indicated that injection of either H-1/O-A or H-2/O-B in the DMH increased arterial pressure and heart rate. 99
H/O projections to the MPFC
The MPFC comprises the anterior cingulate cortex and the prelimbic and infralimbic areas and is involved in both cognitive and visceromotor functions. The MPFC may increase parasympathetic activity and decrease vasomotor sympathetic activity by indirect pathways that involve the amygdala, PAG and NTS (Figure 2). 40 H/O projections target the MPFC, 55,100 which shows a mild expression of H/O receptor 1 at the mRNA 48 and protein 85 levels. H-2/O-B released by H/O projections to the thalamus also excites neurons of the thalamic paraventricular nucleus which send excitatory projections to the MPFC. 101 H/O thus have the potential to modulate the activity of the MPFC both directly and indirectly, but the functional consequences of this modulation still await investigation.
H/O projections to the insular cortex
The insula is a viscerosensory and visceromotor region, which can increase or decrease heart rate depending on the stimulated site. It is not completely clear whether these changes in heart rate are due to sympathetic or parasympathetic effects, which may be exerted through indirect pathways, involving the amygdala, PAG and NTS, as well as through direct projections to the RVLM (Figure 2). 40 H/O neurons project to the insular cortex, 55 but the functional significance of these projections remains unclear. The insular cortex does not express mRNA for H/O receptor 2 and shows only a light expression of mRNA for H/O receptor 1, 48 which is consistent with other results on receptor protein expression. 85
H/O projections to the extended amygdala
The central nucleus of the amygdala and the BNST constitute an anatomo-functional unit called the extended amygdala. 14 The insular cortex and the extended amygdala constitute a corticostriatal–pallidal circuit that processes emotional information with autonomic responses (Figure 2) and projects to the hypothalamic behaviour control column. 40
H/O neurons target the amygdala, including its central nucleus, as well as the BNST. 3,55 The central nucleus of the amygdala and the BNST project back to the H/O neurons. 15 Experiments on brain slices indicate that H/O neuropeptides excite neurons in the central medial nucleus of the amygdala through H/O receptor 2. 102 Although an early report did not show any expression of mRNA for H/O receptor 2 in the central nucleus of the amygdala, 48 expression of this receptor at protein level was reported by a later study. 83 The involvement of H/O neuropeptides in mediating the cardiovascular responses elicited by the amygdala and the BNST is supported by the finding that stimulation of these structures induces long-lasting cardiorespiratory excitation in wild-type mice, but not in H/O-neuron ablated mice. 103 However, the results of a more recent study 104 highlighted the complexity of autonomic cardiovascular control by the BNST. In particular, chemical stimulation of the BNST was found to decrease arterial pressure and heart rate, and this effect was attenuated by H-1/O-A. Pharmacological experiments indicated that H-1/O-A facilitated noradrenaline release in the BNST. Noradrenaline in the BNST would activate local GABAergic neurons through α-2 adrenergic receptors, and GABA released by these neurons would, in turn, inhibit a sympathoinhibitory pathway starting from the BNST by activating GABAA receptors. 104
Conclusions
The evidence discussed in the previous sections indicates that H/O neurons include S0N and P0N, as they send direct projections to S1N in the spinal cord and to P1N in the DMNX. The H/O neurons also project to other CAN structures that include S0N and P0N, such as the RVMM and caudal raphe nuclei, the RVLM and the hypothalamic PVN. While it is not always clear whether the H/O neurons directly target S0N and P0N in these structures or, rather, target neurons in their vicinity, the latter may nonetheless modulate S0N and P0N through local circuits. Therefore, the H/O neurons have the potential to control S1N and P1N both directly and indirectly, through other pre-autonomic neurons of the CAN. There is also abundant evidence that H/O neurons target the NTS, which is a key CAN structure involved in relaying visceral afferent input, and higher order CAN structures including the DMH and the extended amygdala. Significant gaps in our knowledge on the anatomic and/or functional links between H/O neurons and structures of the CAN persist, particularly regarding lower thoracic and lumbar S1N, sacral P1N, the medullary raphe obscurus, the pontine A5 group and PBN, the midbrain PAG, the MPFC and the insular cortex. Nonetheless, the available evidence appears sufficient to regard the H/O neurons as a key component of the CAN (Figure 2).
While a loss of H/O neurons has been reported in the brain of patients with different neurodegenerative diseases, such a loss is far from complete and may even be accompanied by hyperactivity of residual H/O neurons in patients with AD. Moreover, neurodegeneration in PD, DLB and MSA is known to involve the CAN and/or the peripheral autonomic nervous system, thus providing a parsimonious explanation of autonomic dysfunction. Nonetheless, given the substantial role of H/O neurons in the CAN, alterations of H/O signalling have the potential to contribute, at least in part, to autonomic dysfunction in patients with neurodegenerative diseases. On the other hand, it appears biologically plausible that functional alterations of the CAN due to the dramatic H/O neuron deficiency in patients with NT1 underlie the autonomic anomalies reported in these patients, such as alterations in autonomic cardiovascular control during sleep (Figure 1).
Footnotes
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) received no financial support for the research, authorship, and/or publication of this article.