
Abstract
Ghrelin O-acyl transferase (GOAT; MBOAT4) catalyzes O-acylation at serine-3 of des-acyl ghrelin. Acyl ghrelin is secreted by stomach X/A-like cells and plays a role in appetite and metabolism. Therefore, GOAT has been expected to be a novel antiobesity target because it is responsible for acyl ghrelin production. Here, we report homogeneous time-resolved fluorescence (HTRF) and enzyme-linked immunosorbent assay (ELISA) methods utilizing human GOAT-expressing microsomes as a novel high-throughput assay system for the discovery of hit compounds and optimization of lead compounds. Hit compounds exemplified by compound A (2-[(2,4-dichlorobenzyl)sulfanyl]-1,3-benzoxazole-5-carboxylic acid) were identified by high-throughput screening using the HTRF assay and confirmed to have GOAT inhibitory activity using the ELISA. Based on the hit compound information, the novel lead compound (compound B, (4-chloro-6-{[2-methyl-6-(trifluoromethyl)pyridin-3-yl]methoxy}-1-benzothiophen-3-yl)acetic acid) was synthesized and exhibited potent GOAT inhibition with oral bioavailability. Both the hit compound and lead compound showed octanoyl-CoA competitive inhibitory activity. Moreover, these two compounds decreased acyl ghrelin production in the stomach of mice after their oral administration. These novel findings demonstrate that GOAT is a druggable target, and its inhibitors are promising antiobesity drugs.
Introduction
The prevalence rates of obesity are increasing worldwide, and obesity has been recognized as a major contributing factor in heart disease, diabetes, and fatty liver. Safe and effective treatment strategies for obesity have been urgently needed for a long time. 1
Several lines of evidence have suggested that ghrelin controls growth hormone release, energy balance, food intake, and long-term regulation of body weight in rodents and humans.2–6 Ghrelin is predominantly secreted by X/A-like enteroendocrine cells of the stomach. It comprises the 28 N-terminal amino acids of proghrelin, a precursor of 94 amino acids that is cleaved proteolytically to release ghrelin, and undergoes octanoylation on the hydroxyl group of the serine at position 3. 7
Ghrelin Ο-acyl transferase (GOAT; MBOAT4) was identified in 2008 by Gutierrez et al. and Yang et al.8,9 GOAT belongs to the membrane-bound Ο-acyltransferase family 10 and is expressed in X/A-like enteroendocrine cells.11,12 GOAT transfers an octanoyl group from octanoyl-CoA to the third residue of proghrelin. The acyl ghrelin produced by GOAT is a ligand of ghrelin receptor 1a (GHS-R1a), and antagonists of this receptor have been proposed as antiobesity drug targets. 13 However, only one GHS-R1a antagonist has progressed to a clinical trial, despite the perception that it is a traditional target. 14 Recently, a study on GOAT knockout mice revealed improvements in body fat mass, energy expenditure, and blood glucose compared with those in control mice. 15 Therefore, GOAT inhibitors have potential as antiobesity drugs, similar to GHS-R1a antagonists.11,15,16 While small molecules and peptide inhibitors of GOAT have been reported, their oral bioavailability and potency remain unknown.17–23
To screen for GOAT inhibitors, we have developed novel homogeneous microsomal GOAT activity assay methods that utilize biotinylated des-acyl ghrelin and octanoyl-CoA as substrates. Biotinylated acyl ghrelin was detected by the homogeneous time-resolved fluorescence (HTRF) method using an antibody specific for acyl ghrelin. Meanwhile, an enzyme-linked immunosorbent assay (ELISA) was established to confirm that the primary hit compounds do indeed inhibit GOAT. A diverse library of small compounds was screened, and some hit compounds were identified. Subsequently, a potent inhibitor, compound B, was newly synthesized in the lead generation stage based on the structure of one of the hit compounds. Both the hit compound and the lead compound decreased acyl ghrelin production in the stomach of normal mice after their oral administration. In summary, we have identified novel, potent, and orally bioavailable GOAT inhibitors as drug candidates.
Materials and Methods
Construction of GOAT-Expressing Plasmid
Human GOAT (accession no. EU518495) cDNA was amplified by PCR using a human stomach cDNA library (Takara Bio, Kusatsu, Japan) as a template and Pyrobest DNA Polymerase (Takara Bio). Cycling conditions were 1 min at 98 °C, 35 cycles (10 s at 98 °C, 15 s at 65 °C, and 80 s at 72 °C), and final extension for 5 min at 72 °C. The amplified fragments were treated with restriction enzymes, inserted into Stu I/Not I sites of pFastBac1 (Invitrogen, Waltham, MA) using Ligation High (Toyobo, Osaka, Japan), and transfected into ECOS JM109 (Nippon Gene, Tokyo, Japan) to construct pFastBac/human GOAT. The expression vector of mouse GOAT (accession no. EU721729) was prepared as described in a previous study. 17
Baculovirus Infection and Isolation of Microsomes
Baculovirus preparation was achieved with a BacToBac System (Thermo Fisher Scientific, Waltham, MA) using pFastBac/h MBOAT4 and pFastBac/m MBOAT4. The virus titer was measured by real-time PCR. Sf9 cells were infected with the baculovirus at a multiplicity of infection of 0.2 and grown for 70 h. Next, the Sf9 cells were diluted with suspension buffer containing 50 mM Tris-buffered saline (pH 7.5), 1 mM DTT, 1 mM EDTA, and Complete Protease Inhibitor Cocktail (Roche Molecular Systems Inc., Pleasanton, CA), and homogenized. After centrifugation of the cell suspension at 950g for 10 min, the supernatant was collected and ultracentrifuged at 95,680g for 30 min. The pellet was diluted with suspension buffer. The presence of the recombinant protein in the collected fractions was checked by staining with Coomassie Brilliant Blue (Thermo Fisher Scientific), and the activity was analyzed by comparing it with that of the empty vector membrane fraction. This protein solution was used for the microsomes.
HTRF Microsomal Assay
The HTRF assay was established using an acyl ghrelin-selective antibody (mAb7; in-house antibody), Eu(k)-anti-mouse IgG PAb (Cisbio Bioassays, Bedford, MA), and Streptavidin-Xlent (Cisbio Bioassays). The excitation into the streptavidin-conjugated fluorophore and energy transfer into the europium-conjugated secondary antibody led to lanthanide phosphorescence. A microsomal reaction was performed with 30 μg/mL microsomes expressing human GOAT and the substrates 30 μM octanoyl-CoA (Chem-Implex International Inc., Wood Dale, IL), 3 μM biotinylated des-acyl ghrelin (Gly-Ser-Ser-Phe-Leu-Ser-Pro-Glu-His-Gln-Arg-Val-Gln-Gln-Arg-Lys-Glu-Ser-Lys-Lys-Pro-Pro-Ala-Lys-Leu-Gln-Pro-Arg-ε-aminocaproic acid-biotin [Greiner Bio One International GmbH, Kremsmünster, Austria]), and test compound diluted with the assay buffer (50 mM Tris-HCl [pH 7.5], 0.5% Tween 20, and Complete Protease Inhibitor Cocktail [Roche Molecular Systems]) in a reaction volume of 6 μL in Greiner 784075 384-well microplates for 15 min at room temperature. The reaction was stopped by the addition of 2 μL of 0.1 M citric acid/0.2 M Na2HPO4 buffer (pH 3.0), and 2 μL of 150 nM mAb7 diluted with assay buffer was added. Next, 2 μL of Eu(k)-anti-mouse IgG PAb and Streptavidin-Xlent diluted with HTRF detection buffer (Cisbio Bioassays) was added to the reaction and incubated at room temperature for 1 h. The biotinylated acyl ghrelin produced by human GOAT was detected by measuring fluorescence at 620 and 665 nm with an EnVision Multiplate Plate Reader (PerkinElmer, Waltham, MA). The results of this production are expressed as (665 nm/620 nm) × 104. We considered the signal ratio of the reaction without microsomes expressing human GOAT to indicate 100% inhibitory activity, and that of the complete reaction mixture to indicate 0% inhibitory activity. An increase in the detection of the signal of 620 nm fluorescence was considered to confirm GOAT activity, while the signal of 665 nm fluorescence remained constant. Curve fitting and calculation of 50% inhibitory concentrations (IC50) were performed with GraphPad Prism v5 (GraphPad Software, San Diego, CA). A mouse GOAT microsomal assay was conducted in the same way as the human GOAT microsomal assay.
The kinetic parameters for octanoyl-CoA and biotinylated des-acyl ghrelin were determined by incubating various concentrations of each substrate with 30 μg/mL microsomes, and the initial rate data were fitted to the Michaelis–Menten equation using GraphPad Prism v5. Competition assays were performed with the following four conditions: 5 and 500 μM octanoyl-CoA in the presence of 1 μM biotinylated des-acyl ghrelin, and 3 and 30 μM biotinylated des-acyl ghrelin in the presence of 10 μM octanoyl-CoA. The assay quality of high-throughput screening was analyzed using the Z′ factor. 24 This factor was calculated from the mean and standard deviation (SD) values of the maximum and minimum controls. The maximum control was defined as the highest activity under the assay conditions. The minimum control was defined as the activity in the absence of microsomes.
Microsomal ELISA
An ELISA was established on the basis of competition between horseradish peroxidase (HRP)–conjugated ghrelin and acyl ghrelin using 50 ng/mL mAb7-coated plates ( Suppl. Fig. S1A ). HRP conjugation of acyl ghrelin (Peptide Institute Inc., Osaka, Japan) was carried out with an HRP Labeling Kit (Dojindo, Kumamoto, Japan). The GOAT microsomal reaction was conducted using the same method as the HTRF assay, and the reaction mixture was subjected to the ELISA analysis. The microsomal activity was detected by luminescence using the SuperSignal ELISA Femto Substrate (Thermo Fisher Scientific).
Acetyl–Coenzyme A Acetyltransferase 1 Selectivity Assay
An assay in which the activity of human acetyl–coenzyme A acetyltransferase 1 (ACAT1) was detected by thin-layer chromatography (TLC) was performed as described previously. 25 In total, 5 μM [1- 14 C]-oleoyl-CoA (PerkinElmer) and 100 μM cholesterol were reacted with 30 μg/mL human ACAT1 enzyme for 40 min at 32 °C. The modified Bligh and Dyer method was applied to the sample preparations. 26 The organic phase was applied to a silica TLC plate (Merck KGaA, Darmstadt, Germany) and separated with a solvent system of hexane, ethyl ether, and acetic acid. The radioactivities incorporated into the lipids were measured with a Typhoon FLA 7000 (GE Healthcare UK Ltd., Little Chalfont, England) and analyzed with Image Quant TL software (GE Healthcare UK Ltd.).
In Vivo Pharmacological Assay and Tissue Sample Preparations
Male C57BL/6J mice were purchased from CLEA Japan Inc. (Tokyo, Japan) or Charles River Japan Inc. (Kanagawa, Japan). The mice were fed a standard chow of CE-2 (CLEA Japan) and orally administered vehicle or compound A at 30 mg/kg. At 1 h after administration, the mice were orally given a lipid emulsion (containing 20% volume of tricaprylin: a triglyceride of C8-length fatty acid). Stomach tissues were obtained at 4 h after lipid loading, boiled for more than 10 min, and acidified with a mixture of hydrochloric acid and acetic acid prior to analysis, as reported by Hosoda et al.27,28
Other mice received the oral administration of vehicle or compound B at 1, 3, or 10 mg/kg/day once daily for 3 days. After 3 days of treatment, blood samples were collected into tubes containing Na2EDTA (Dojindo) and aprotinin (Wako Pure Chemical Industries Ltd., Osaka, Japan) at final concentrations of 1.25 mg/mL and 500 U/mL, respectively, and centrifuged at 2000g for 2 min at 4 °C to separate the plasma. The obtained plasma samples were immediately treated with 1/10 volume of 1 mol/L hydrochloric acid and stored at −80 °C until measurement. Stomach samples were also obtained. These samples were washed with chilled phosphate-buffered saline (pH 7.4, Thermo Fisher Scientific), weighed, boiled in 1 mL of distilled water for 15 min, acidified with a mixture of 70 μL of acetic acid and 20 μL of 1 N hydrochloric acid, homogenized, and separated by centrifugation at 2000g for 5 min at 4 °C. Tissue-active ghrelin and des-acyl ghrelin were measured using Active Ghrelin ELISA and Desacyl-Ghrelin ELISA (Sceti Medical Labo K.K., Tokyo, Japan). All animal experiments were performed in compliance with the Guidelines for the Care and Use of Laboratory Animals of Takeda Pharmaceutical Co. Ltd (Kanagawa, Japan). All data are expressed as the mean and SD. The statistical significance of differences between the sample means was assessed by Student’s t test. For evaluation of dose–response effects, the one-tailed Williams’ test was employed.
Pharmacokinetics Analysis of Mouse Cassette Dosing
Test compounds were administered intravenously (0.1 mg/kg) or orally (1 mg/kg) by cassette dosing to nonfasted mice. After administration, blood samples were collected from the caudal vein at 0.083, 0.167, 0.25, 0.5, 1, 2, 4, and 8 h and 0.25, 0.5, 1, 2, 4, and 8 h after intravenous and oral administrations, respectively. The samples were centrifuged at 13,000g for 30 min at 5 °C to obtain the plasma fraction. Aliquots (5 μL) of the plasma samples were mixed with acetonitrile containing the internal standards, and the mixtures were centrifuged at 3600g for 7 min at 25 °C. The compound concentrations in the resulting supernatants were measured by AB Sciex API5000 (McKinley Scientific, Sparta, NJ) coupled with Shimadzu UFLC (Shimadzu, Kyoto, Japan).
Results
Evaluation of GOAT Microsomal Activity
To evaluate GOAT enzyme activity, the microsomal fraction of Sf9 cells expressing human GOAT was purified and the HTRF assay system was established in accordance with the method shown in Figure 1A . From a reaction progression curve obtained by mixing various concentrations of microsomes and substrates, the reaction time and the concentration of microsome-dependent activities were confirmed ( Fig. 1B ). As a result, the initial linear portion of the enzyme reaction was determined and subsequent experiments were conducted within this linear range. The apparent Km values of biotinylated des-acyl ghrelin and octanoyl-CoA were determined by the Michaelis–Menten equation, and their values were 3.6 and 15 μM, respectively ( Fig. 1C , D ). Figure 1C appears to show systemic deviation in the fitted curve determining the Km value of biotinylated des-acyl ghrelin. This reaction was performed with incubation at various concentrations of biotinylated des-acyl ghrelin, and the other components were used at fixed concentrations. Therefore, the balance between biotinylated des-acyl ghrelin and Streptavidin-Xlent was different at each concentration of biotinylated des-acyl ghrelin. We suppose that this imbalance is one of the factors for the systematic deviation. The Km values of biotinylated des-acyl ghrelin and octanoyl-CoA detected by ELISA were 1.0 and 9.2 μM ( Suppl. Fig. S1B,C ), and similar values could be confirmed. Moreover, the signals of the reaction mixture with microsomes expressing empty vector and without microsome were evaluated, and we considered that the reaction without microsome could be set as a sample with 100% inhibitory activity ( Suppl. Fig. S2 ).
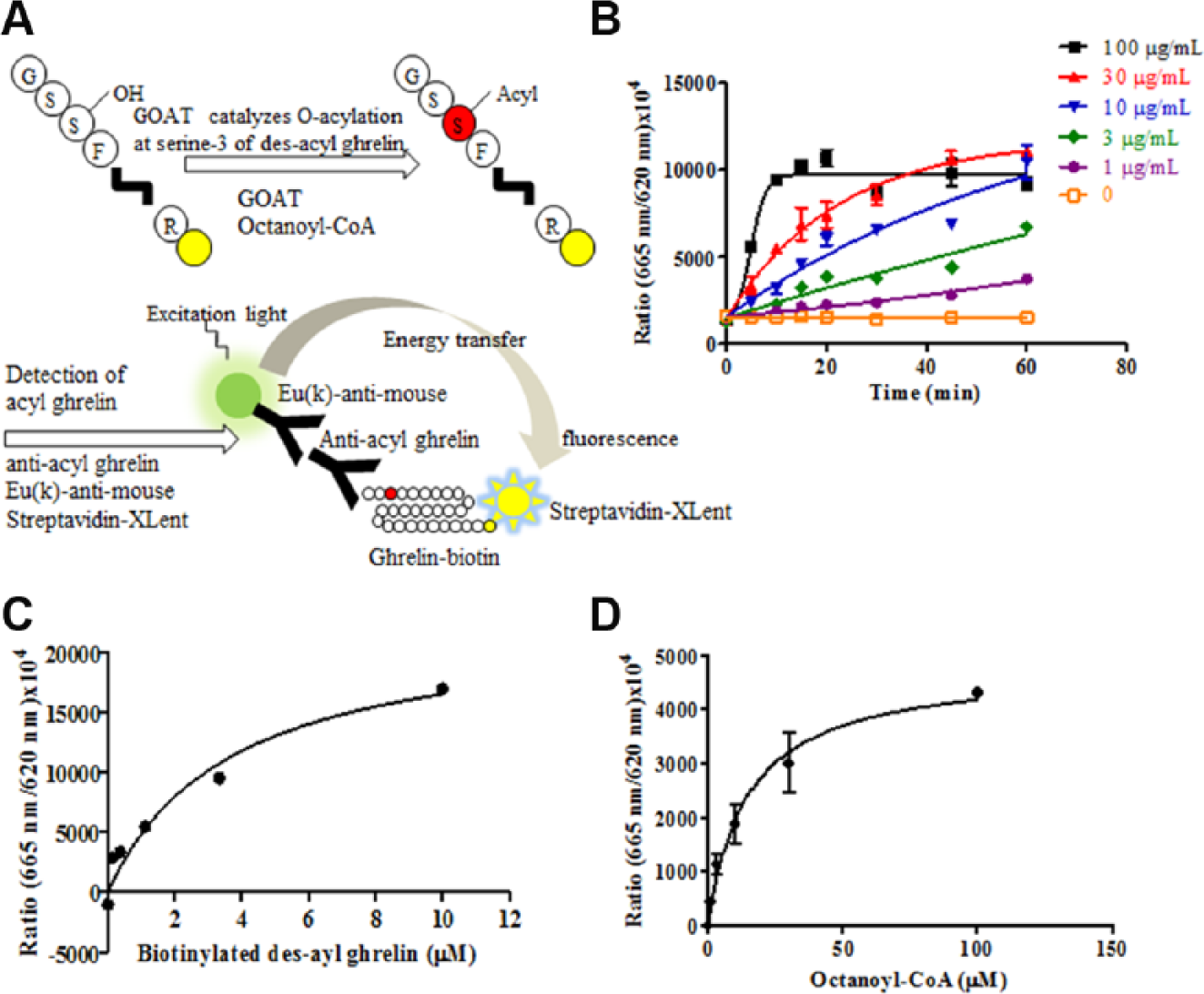
Evaluation of human GOAT microsomal activity. (
The substrate concentrations in the screening condition were set at 30 μM octanoyl-CoA and 3 μM biotinylated des-acyl ghrelin based on the above results. The reason for these selections was that the concentrations of substrates should be set around the apparent Km values to obtain a diversity of screening hits. 29 The inhibitory activity of octanoylated [Dap3]-ghrelin 17 was evaluated in this assay condition, and this peptide showed an IC50 value of 15 nM ( Suppl. Fig. S2D ). Furthermore, the HTRF assay was selected as the primary screening assay in order to ensure high throughput because of its homogeneous method.
High-Throughput Screening to Identify Novel GOAT Inhibitors
A diverse library of more than 500,000 small-molecule compounds from Takeda Pharmaceutical Co. Ltd. was screened at a concentration of 1 μM by the HTRF assay, as shown in the workflow ( Fig. 2A ). The assay quality was judged acceptable because the Z′ factor was 0.83 and the signal-to-background ratio was 6.3. A histogram of the primary screening showed a normal distribution curve ( Fig. 2B ), with a median value of 7.4 and SD of 12. The hit rate was 0.05% when the threshold was set at 33% or more inhibitory activity of the median and 3 SD through the deconvolution. A total of 301 compounds were selected from the mother plates and tested for their inhibitory potencies by the ELISA as a confirmatory assay to eliminate compounds with autofluorescence or reactivity toward the antibodies in the HTRF assay. Next, the inhibitory activities of 190 compounds against ACAT1 were tested by ELISA to examine their selectivity. ACAT1 is a member of the MBOAT family, and ACAT1 inhibitors were reported to induce adrenal toxicities.30,31 Therefore, we chose 39 compounds that showed no activity against ACAT1. The remaining hit compounds were tested by the mouse GOAT assay because we planned to confirm any in vivo activity using high-fat diet-induced obese mice. Similar inhibitory activities of these hit compounds for mouse GOAT were confirmed ( Fig. 3B ). After this step, 39 active compounds were selected and clustered into 26 groups with similar structural features based on Ward’s method. 32 Finally, certain compounds represented by compound A were chosen ( Fig. 3A , Suppl. Tables S1 and S2 ) because of chemical tractability. Subsequently, compound A was further optimized to compound B (Supplementary Information), 33 yielding improvements in the biochemical potency and bioavailability ( Fig. 3A , C , Suppl. Table S2 ).
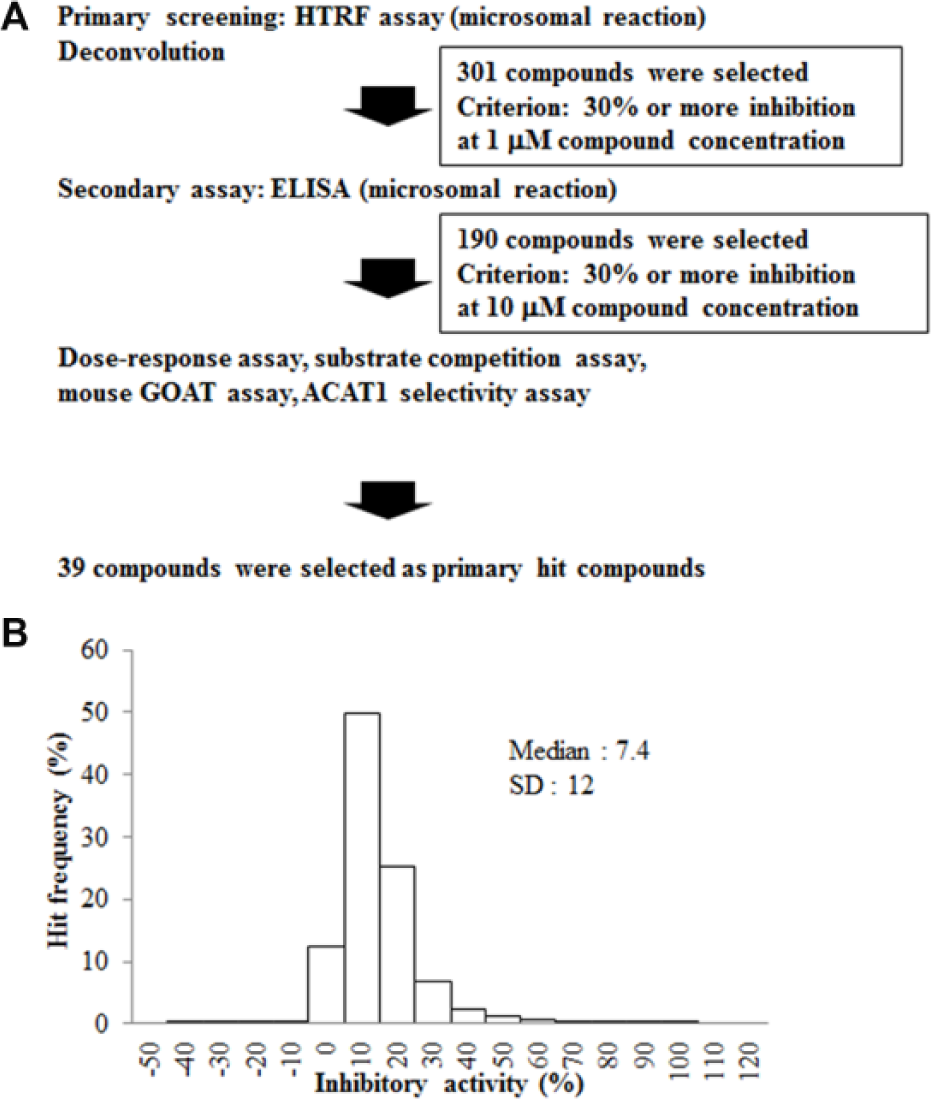
Results of the primary screening. (
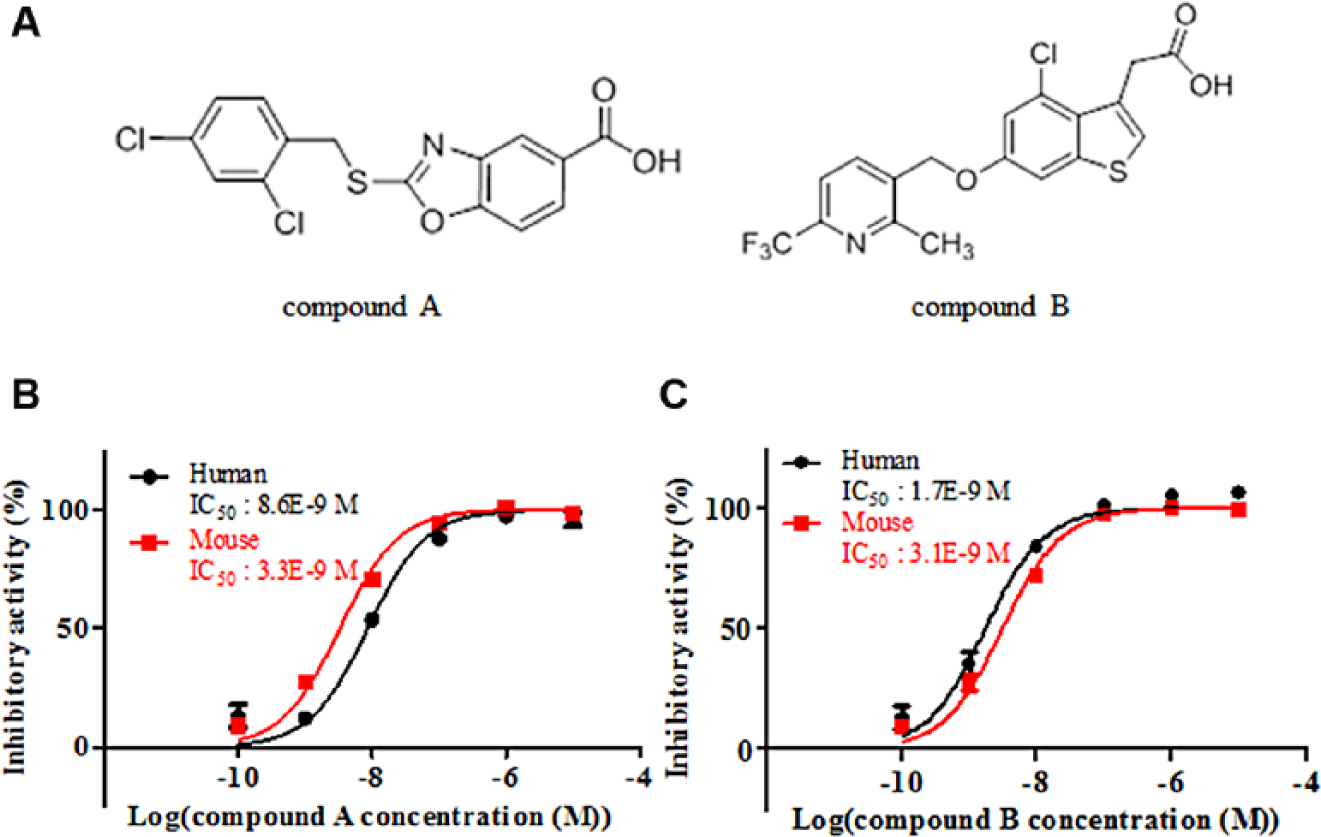
Inhibitory activities of the hit and lead compounds. Structures of the compounds. (
Characterization of the Hit and Lead Compounds
To understand the mechanism of inhibition, substrate competition assays were performed. The GOAT inhibition rates of the compounds were examined using four reaction conditions. The representative hit compound (compound A) and lead compound (compound B) showed a right shift in the 50% IC50 values at the high concentration of octanoyl-CoA. Meanwhile, neither compound caused a shift at the high concentration of des-acyl ghrelin. These results indicated that our compounds possessed competitive inhibitory activity against octanoyl-CoA and noncompetitive inhibitory activity against des-acyl ghrelin ( Fig. 4 ).
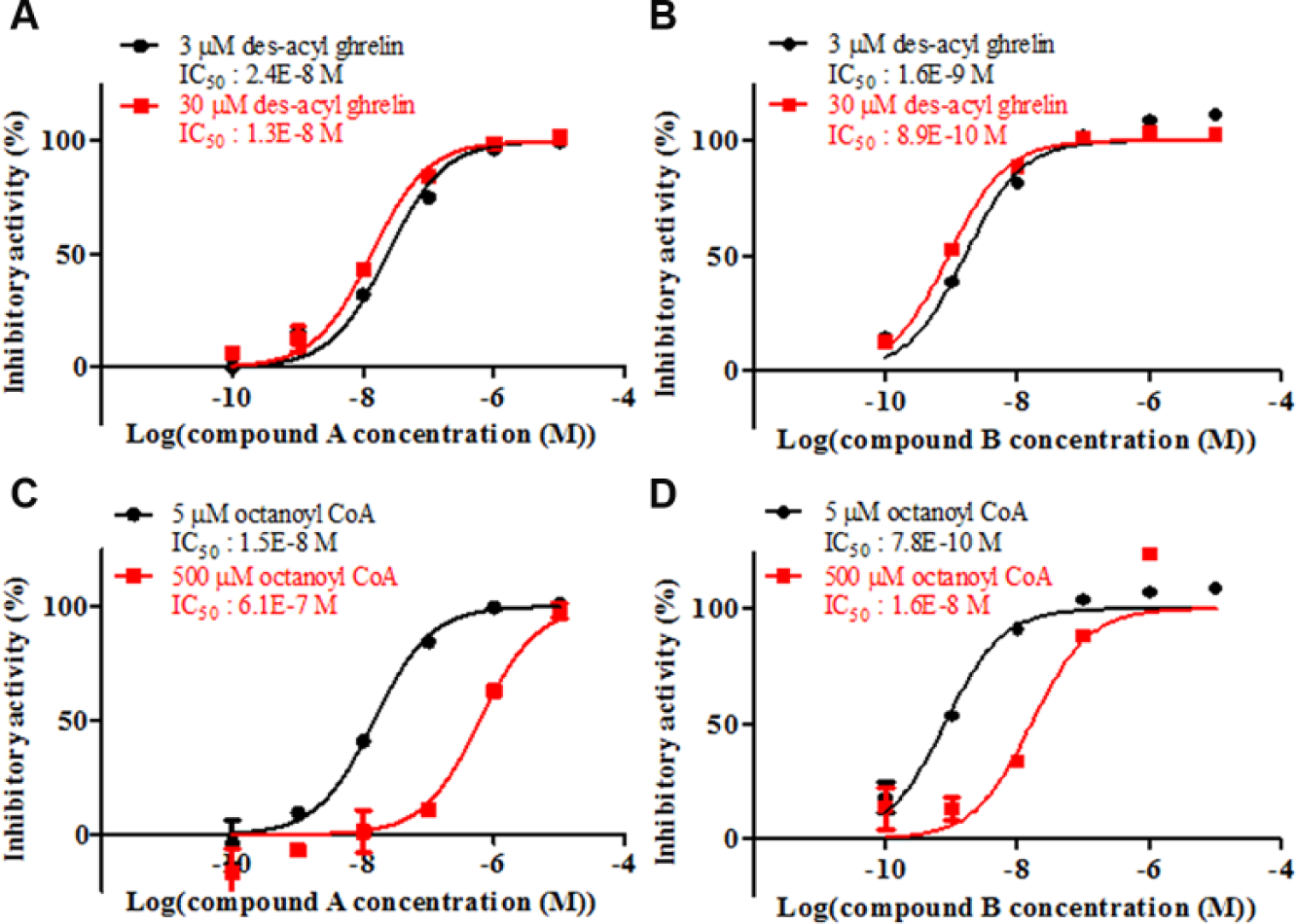
Substrate competition assays. (
In Vivo Efficacies of Hit Compound and Lead Compound
Since compounds A and B had mouse GOAT inhibitory activities ( Fig. 3B , C ), the efficacies of these compounds were evaluated in normal mice. Both compounds decreased the level of acyl ghrelin in the stomach. While compound A orally administered as a single dose at 30 mg/kg decreased acyl ghrelin and total ghrelin (acyl ghrelin plus des-acyl ghrelin) in the stomach ( Fig. 5D , F ), compound A administered at the same concentration had no effects on plasma ghrelin content ( Fig. 5A–C ). A possible reason for the results of compound A is that it did not continue for a sufficient amount of time. The ghrelin peptide is produced in the stomach and secreted into the plasma. A continuous inhibitory effect against GOAT might be necessary to affect the plasma ghrelin concentration. In contrast, compound B with higher activity and bioavailability showed efficacy in the plasma and stomach in the mice after oral administration for 3 days. Decreases in acyl ghrelin in the plasma and stomach were detected in a dose-dependent manner at 1 mg/kg and above ( Fig. 6A ,D). The concentrations of total and des-acyl ghrelin increased in the plasma and stomach in response to the decrease in acyl ghrelin ( Fig. 6B , C , E , F ). It was supposed that compounds A and B showed different phenotypes because of differences in their bioavailability and method of administration. The phenotype of an increasing concentration of des-acyl ghrelin was also observed in GOAT knockout mice. 15 While the reason for the increase in total ghrelin after the administration of compound B remains unclear, we suggest that the mice exhibit activity to compensate for the decrease in acyl ghrelin as a result of strong inhibition by the compound. In the present study, we did not examine the impact of GOAT inhibition on the gene expression level of ghrelin in the stomach. Both compounds did not decrease the total ghrelin content in the stomach; in particular, compound B significantly increased the total ghrelin content in the stomach. These results suggested that a decrease in acyl ghrelin was not due to the regulation of ghrelin gene expression. However, further analyses would be needed to conclude the mechanism of inhibition.
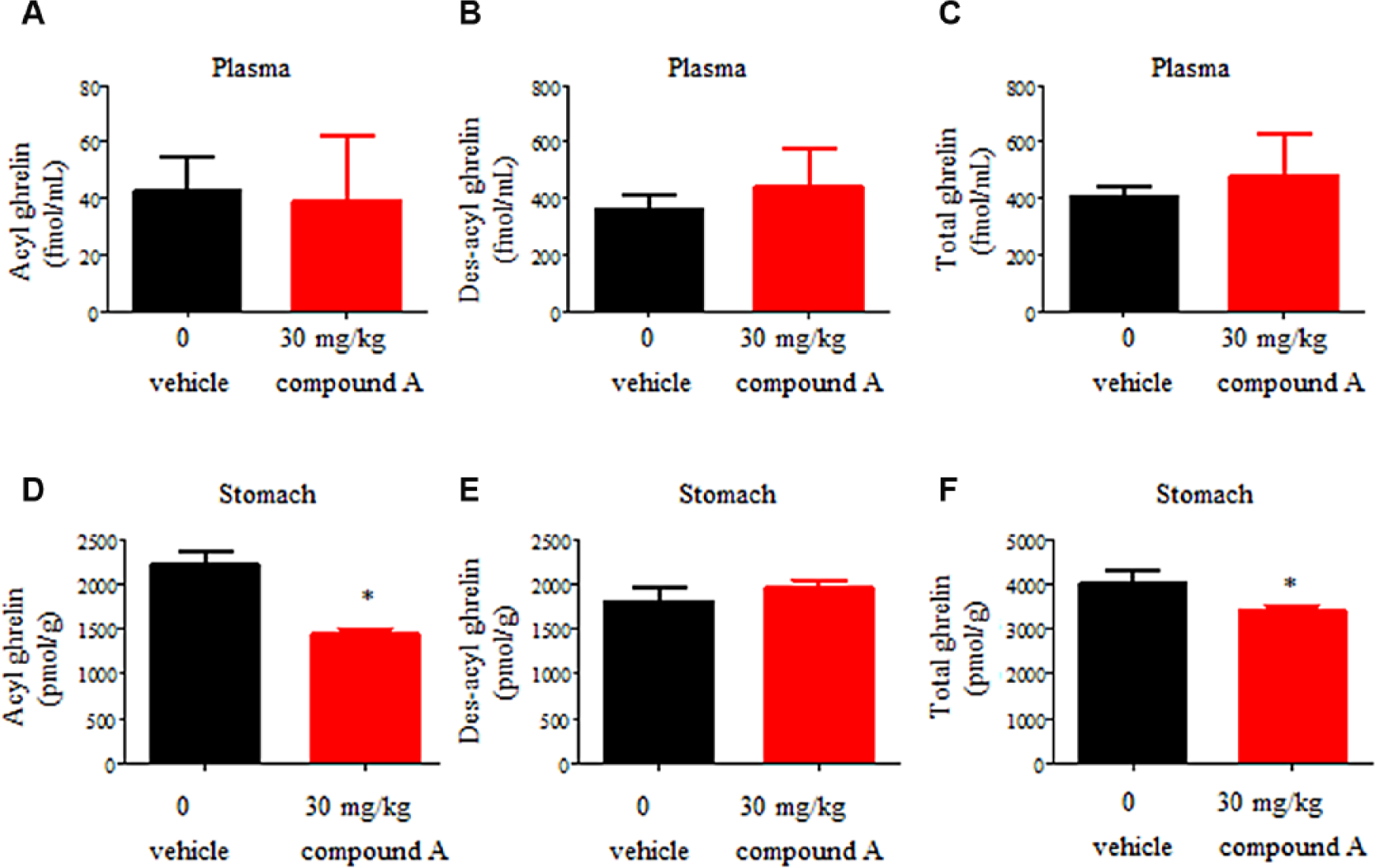
Effects of oral administration of 30 mg/kg compound A on acyl ghrelin and des-acyl ghrelin levels in the plasma and stomach. Plasma levels of acyl ghrelin (
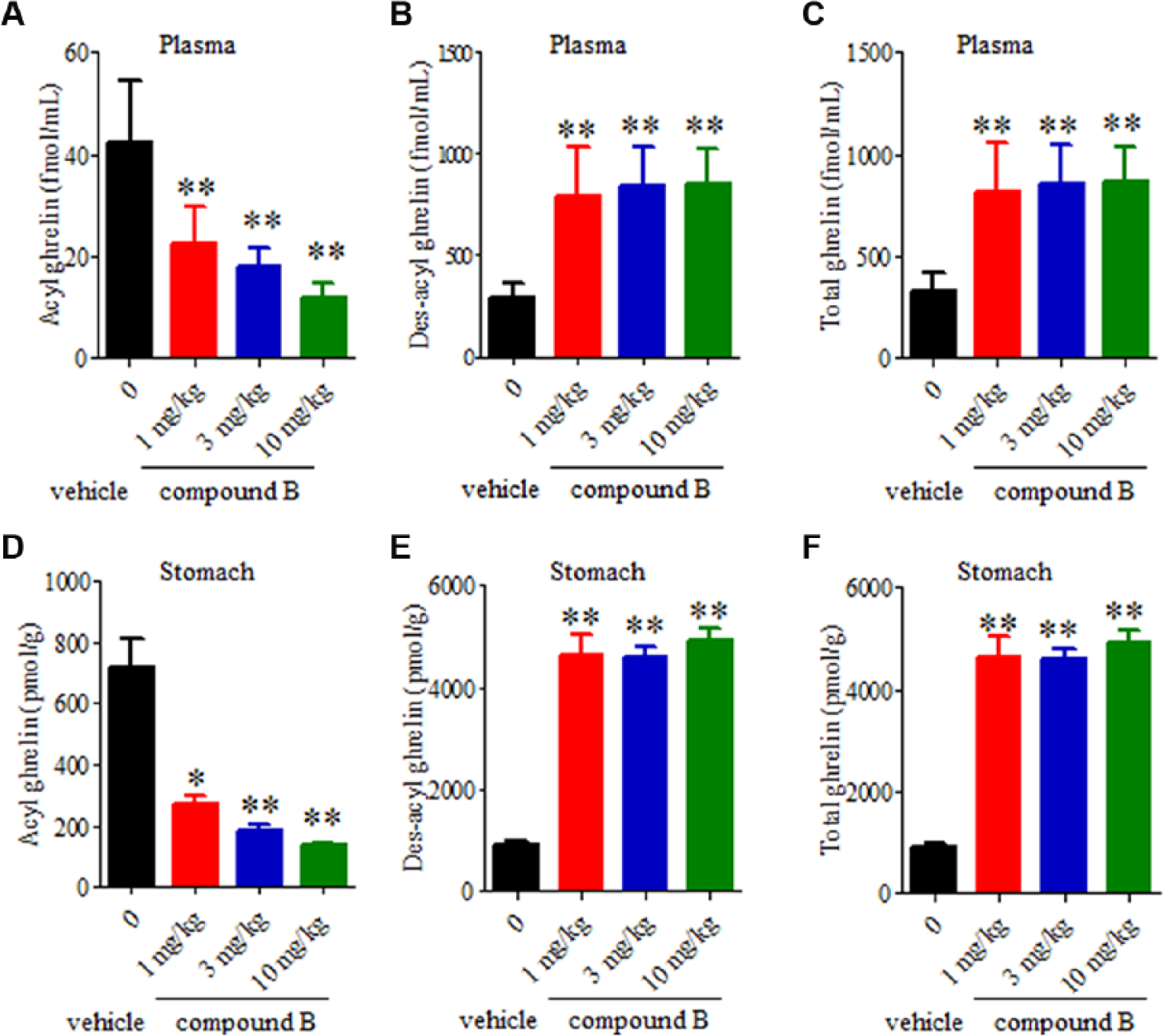
Effects of oral administration of 1, 3, or 10 mg/kg compound B once daily for 3 days on acyl ghrelin and des-acyl ghrelin levels in the plasma and stomach. Plasma levels of acyl ghrelin (
Moreover, the esterase inhibitors were not used when the blood and stomach samples were collected. However, in our experiments, we collected the samples with utmost care and attention to prevent acyl ghrelin from converting into des-acyl ghrelin. Although more investigation, such as the treatment with esterase inhibitors, is needed to determine the precise concentration of tissue ghrelin levels, our current protocol is relatively appropriate to maintain tissue acyl ghrelin concentration because tissue enzymes are expected to be inactivated by acidification and boiling.27,28
Discussion
We successfully developed a homogeneous and robust GOAT biochemical assay method, identified hit compounds by high-throughput screening, and optimized a lead candidate compound. Some in vitro GOAT assays using microsomes were reported previously. Yang et al. 17 established a mouse GOAT assay using 3H-labeled octanoyl-CoA and proghrelin-His8. Meanwhile, Garner and Janda 20 developed a fluorescence GOAT assay based on cat-ELISA. Most in vitro assay systems have limitations of high cost and low throughput, which need to be overcome for them to be used in large-scale screening. Our newly established HTRF assay system requires only a couple of hours to confirm human and mouse GOAT activities, and thus makes screening of a large compound library possible. Biotinylated des-acyl ghrelin was used as a substrate in the assay because we expected that the difference in C-terminal peptide lengths between proghrelin and ghrelin would minimally affect the reaction with GOAT. Des-acyl ghrelin and octanoyl-CoA are considered to be endogenous substrates of GOAT because it is localized in the endoplasmic reticulum (ER) membrane and the octanoyl reaction occurs in the ER lumen.34,35 Indeed, the apparent Km value of biotinylated des-acyl ghrelin was similar to the data obtained using proghrelin, as reported by Yang et al. 17 In the process of optimization in the HTRF assay, the lower concentration of mAb7 was set in comparison with that of biotinylated des-acyl ghrelin. The high concentration of mAb7 decreased the fluorescence signal of 620 nm ( Suppl. Fig. S2C ). The balance between mAb7 and HTRF components affected the signal intensity.
Some GOAT inhibitors were identified by high-throughput screening. The HTRF assay performed for the primary screening and the ELISA carried out for the secondary screening exhibited reasonable assay performance. Compound A, as one of the hit compounds, possessed high activity with an IC50 value of 8.6 nM, competitive activity against high concentrations of octanoyl-CoA, no inhibitory activity against ACAT1, and similar activity for mouse GOAT in the HTRF assay. The results for the octanoyl-CoA competition assays in this study provide new insights. Small-molecule inhibitors, such as our hit and lead compounds, are reasonably thought to occupy the octanoyl-CoA-binding site rather than the des-acyl ghrelin-binding site. Although there are no available data for the octanoyl-CoA concentrations in mouse plasma, the reported concentrations in human plasma were 70 36 and <770 nM. 37 As these concentrations are lower than the apparent Km value of human GOAT, octanoyl-CoA competitive GOAT inhibitors are predicted to be effective in humans.
Regarding the generation of a lead compound, compound B was newly synthesized based on the structures of the hit compounds using the highly potent and bioavailable chemotype containing compound A, and showed improved activity with an IC50 value of 1.7 nM and bioavailability ( Suppl. Table S2 ). The area under the plasma concentration curve value for the mouse cassette dosing by oral administration was 120-fold higher than that for the hit compound. The lead compound showed decreased acyl ghrelin concentrations in the plasma and stomach and increased des-acyl ghrelin concentration in the mice after oral administration at 1 mg/kg once daily for 3 days. In the field of metabolic disease, a variety of beneficial functions of des-acyl ghrelin have been reported to improve insulin sensitivity and glucose homeostasis in hepatocytes 38 and adipose tissue 39 and to suppress food intake. 40 These results suggest that des-acyl ghrelin could play a key role in metabolism, although the signal mechanism remains to be identified. In vivo efficacy studies were performed and will be the subject of a subsequent publication, based on the fact that our compounds show on-target activity in the mice as well. Moreover, the reason for the change of the concentration of des-acyl ghrelin and total ghrelin by the compounds could be clarified.
Several molecules in the ghrelin system, including GOAT, are expressed in a wide variety of tissues and play crucial roles by regulating multiple biological processes other than those in metabolic disorders, such as in aging and neurodegenerative disorders.16,41,42 Analyses using GOAT inhibitors can improve our understanding of the molecular mechanisms that are particularly important in several therapeutic areas.
In summary, we have successfully developed a homogeneous assay system and identified small-molecule inhibitors of GOAT that showed activity both in vitro and in vivo.
Footnotes
Acknowledgements
We thank Mr. Shoichi Okubo for providing help with performing DNA cloning, Mr. Takashi Ito and Mr. Yukiya Tanaka for protein expression and purification, and Dr. Nobuyuki Suzuki for antibody construction. We also thank Ms. Kumiko Hazama and Dr. Ryo Fujii for initial exploration of the target, Ms. Michiyo Tokuhara and Mr. Kazutoshi Kawakami for evaluation of the compounds, and Dr. Minoru Ikoma for synthesis of the lead compounds. We would like to thank Mr. Shinichi Niwa for help with the DMPK evaluation and Mr. Motoo Iida for the structural determination with nuclear magnetic resonance experiments. Finally, we acknowledge Dr. Naoki Tarui for supervision of the research and valuable suggestions.
Supplementary material is available online with this article.
Declaration of Conflicting Interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The authors received no financial support for the research, authorship, and/or publication of this article.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.