
Abstract
This study investigated neuroanatomic, genetic, cognitive, sociodemographic and emotional underpinnings of the Negative Urgency subscale of the Urgency, Premeditation, Perseverance, Sensation-Seeking and Positive Urgency Impulsive Behavior Scale in a healthy developmental sample. The goal of the investigation is to contribute to the harmonisation of behavioural, brain and neurogenetic aspects of behavioural self-control. Three domains – (1) Demographic, developmental, psychiatric and cognitive ability; (2) Regional brain volumes (neurobiological); and (3) Genetic variability (single nucleotide polymorphisms) – were examined, and models with relevant predictor variables were selected. Least absolute shrinkage and selection operator and best subset regressions were used to identify sparse models predicting negative urgency scores, which revealed that variables related to emotional regulation and right cingulate volume, as well as single nucleotide polymorphisms in CADM2 and SLC6A4, were associated with negative urgency. Our results contribute to the construct and criterion validity of negative urgency and support the hypothesis that negative urgency is a result of a complex array of influences across domains whose integration furthers developmental psychopathology research.
Negative urgency (NegUrg) is defined as impulsive behaviour in the context of unpleasant emotion (Whiteside et al., 2005). Construct validity of NegUrg is found across the lifespan, though less research focuses on paediatric populations (Geurten et al., 2018; Marmorstein, 2013). The present exploratory study considers the construct and criterion validity of the Urgency, Premeditation, Perseverance, Sensation-Seeking and Positive Urgency (UPPS-P) NegUrg scale through the examination of three domains in a sample of healthy children, adolescents and young adults: (1) Demographic, developmental, psychiatric, and cognitive ability; (2) Regional brain volumes (neurobiological); and (3) Genetic variability (single nucleotide polymorphisms (SNPs)).
Developmental, cognitive, and affective/psychiatric
Developmental, demographic, affective and cognitive variables each contribute to impulsivity, which increases in adolescence (Cyders and Smith, 2008) and declines in young adulthood (Green et al., 1999). ‘Fluid’ cognitive abilities, including executive function (EF) and attention processing of novel information, are developmentally similar (Cattell, 1963; Horn and Cattell, 1967) and measured by tasks such as the Dimensional Change Card Sort (DCCS) and Flanker Inhibitory Control and Attention Tests (Zelazo et al., 2013). Trajectories of these cognitive abilities coincide with maturation of the prefrontal cortex (PFC; Romer, 2010), with inverse association to impulsivity (Stanford et al., 2009). Other developmental factors such as moderate maternal alcohol consumption during pregnancy increase risk for offspring impulsivity (Flak et al., 2014) and are linked to attention-deficit/hyperactivity disorder (ADHD; Eichler et al., 2018; Eilertsen et al., 2017).
Gender and socioeconomic status (SES) may relate to impulsivity; however, sampling differences complicate interpretation. In one meta-analysis, women had higher NegUrg, with small effect size (d = –0.10; Cross et al., 2011). A study of 1372 undergraduates did not identify meaningful differences (Cyders, 2013). Children from low SES backgrounds performed worse on EF tasks (Arán-Filippetti and Richaud de Minzi, 2012), suggesting vulnerability to cognitive control issues.
Brain volumes
Neurological function forms much of the basis for behaviour, and regional brain volumes represent a readily available metric for investigation. Research has focused on development of self-regulation for people with psychiatric diagnoses (e.g. Fjell et al., 2012), though ratings of low impulse control predicted decreased volume in right ventromedial PFC in healthy boys (Boes et al., 2009). In healthy, educated young adults, NegUrg was negatively associated with frontal-striatal and temporal volume (Muhlert and Lawrence, 2015). Fjell et al. (2012) found right caudal anterior cingulate cortex (ACC) surface area negatively predicted cognitive control, especially in children 12 years or younger. Research shows associations of NegUrg and PFC suggestive of developmental psychopathology. In functional magnetic resonance imaging (fMRI) studies, NegUrg contributes to cingulate and orbital activation (Cyders et al., 2014, 2015; Joseph et al., 2009).
Genetic
Impulsivity is moderately heritable, accounting for 20% to 62% of variance (Bevilacqua and Goldman, 2013; Congdon and Canli, 2008; Hur and Bouchard, 1997; Pederson et al., 1988; Seroczynski et al., 1999). Family genetics and environment influence performance-based impulsivity (Dougherty et al., 2003). Multivariate association of impulsivity measures and SNP data have identified various genetic components of pathways associated with impulsivity and related neural constructs, including brain development, nervous system signal generation, amplification or transduction, calcium signalling and immune response (Bevilacqua and Goldman, 2013; Khadka et al., 2014; Whelan et al., 2012).
GABRA2 alleles, encoding the alpha subunit of the GABA-A ‘inhibition’ receptor, explained 2.6% to 4.8% of variance in impulsivity (Villafuerte et al., 2012). Genetic contributions are also mediated by serotonin (5HT) and dopamine (DA) (Bevilacqua and Goldman, 2013; Congdon and Canli, 2008). 5HT and DA neuromodulation in amygdala and orbitofrontal cortex (OFC) contribute to urgency (Cyders and Smith, 2008). DA levels are associated with emotional dysregulation, ADHD and behavioural disinhibition/impulsivity (Cardinal and Everitt, 2004; Friedel, 2004; Winstanley et al., 2004a). DA D2 and D4 receptors, within PFC–amygdaloid–OFC circuitry, are genetically linked to these effects (Floresco and Tse, 2007; Winstanley et al., 2004b).
CADM2, DBH, AVPR1A, DRD2, SLC6A3 and SLC6A4 are implicated in executive dysfunction, emotional reactivity and impulse-control-based psychopathology (Albayrak et al., 2013; Golds et al., 2020; Kieling et al., 2008; Levin et al., 2009; Mick et al., 2010:; Zald and Treadway, 2017), and FOXP2, ATP2C2 and ROBO1 are implicated in speech and language, among others (see Koomar and Michaelson, 2020).
Methods
Whiteside and Lynam (2001) originally developed a four-factor (Urgency, Premeditation, Perseverance and Sensation-Seeking Impulsive Behavior (UPPS)) model of impulsivity. Subsequent validation research indicated that Urgency is the UPPS impulsivity factor most associated with psychopathology (Whiteside et al., 2005). Urgency involves difficulty coping with urges in response to unpleasant emotion, possibly reflecting the intersection of impulsivity and negative affect (Whiteside et al., 2005). A revision of the UPPS scale, the UPPS-P, added on a fifth subscale – Positive Urgency – to reflect a distinction between impulsive behaviour aimed at sustaining positive emotions versus impulsive behaviour aimed at avoiding negative emotions (NegUrg; Lynam et al., 2006).
Based on existing literature on their demonstrated and theoretical relevance to impulsivity and negative affect, we selected demographic, cognitive, neurobiological and genetic variables that we hypothesised would be predictive of NegUrg. Cognitive variables hypothesised to be less related to impulsivity (those measuring ‘crystallised abilities’) were also included in the model. We then conducted exploratory analyses to determine the most optimal combination of variables in predicting self-reported NegUrg. Through the determination of salient markers related to NegUrg, high-risk children can be identified early to reduce subsequent risk for the development of associated psychopathology.
Cognitive, sociodemographic, imaging and genetic data for this study were obtained from the Pediatric Imaging, Neurocognition and Genetics (PING) study (Jernigan et al., 2016). The PING data repository consists of multimodal, multisite, cross-sectional data collected from a community-dwelling (i.e. non-clinical) sample of 1493 children ranging in age from 3 to 20 years. See Jernigan et al. (2016) for a more complete description of the entire PING cohort, exclusionary criteria and data collection methods.
Participants
Data from a subset of 225 participants, ages 7 to 21, for whom data from the PhenX Toolkit (https://www.phenxtoolkit.org; which includes the UPPS-P) were available, were selected for this study. Descriptive data for the participants are presented in Table 1.
Participant characteristics.

Data collection and measures
Criterion variable
The NegUrg 12-item self-report subscale of the UPPS-P (Lynam et al., 2006) was used to measure difficulty coping with urges in response to unpleasant emotion. The measure used a Likert-type scale, ranging from 1 (agree strongly) to 5 (disagree strongly). All items except for one were reverse coded and included statements, such as ‘I have trouble controlling my impulses’ and ‘When I feel bad, I will often do things I later regret in order to make myself feel better now’. Item scores were averaged to get a total NegUrg score, with higher scores reflecting higher self-reported NegUrg.
Predictor variables
As described in the introduction, important relationships with NegUrg have been implicated in three domains: (1) Demographic, developmental, psychiatric, and cognitive ability; (2) Regional brain volumes (neurobiological); and (3) Genetic variability (SNPs). For each domain, predictor variables were selected based on the literature supporting their relevance to NegUrg. See Table 2 for a complete list of predictor variables examined.
Predictor variables examined by model.
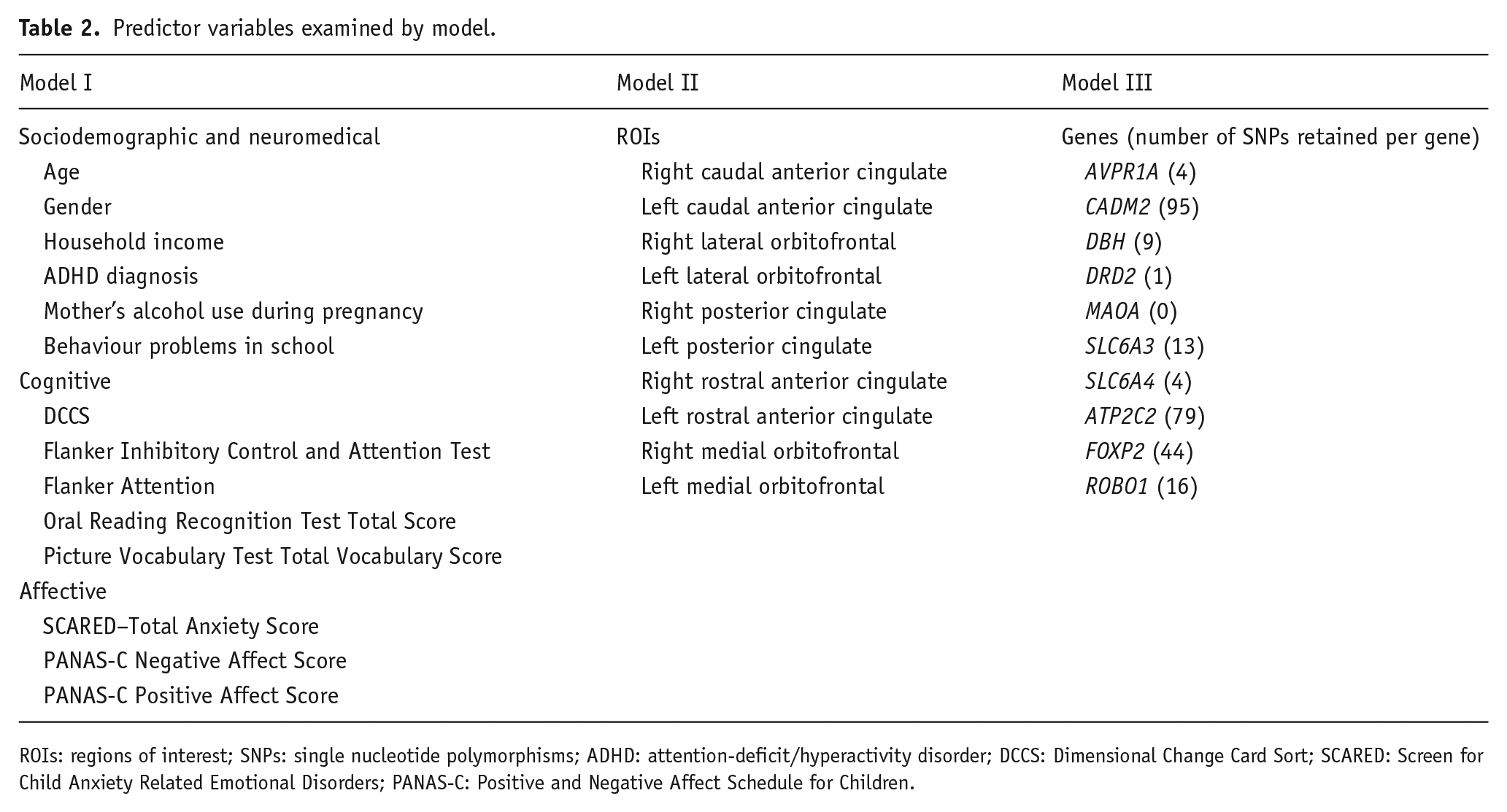
ROIs: regions of interest; SNPs: single nucleotide polymorphisms; ADHD: attention-deficit/hyperactivity disorder; DCCS: Dimensional Change Card Sort; SCARED: Screen for Child Anxiety Related Emotional Disorders; PANAS-C: Positive and Negative Affect Schedule for Children.
Model I: sociodemographic, medical history, cognitive and affective data
Data on basic sociodemographic information, medical, developmental and behavioural history, as well as family history of medical and neuropsychiatric disorders, were collected using the self- or parent/guardian-reports from the PING Study Demographic and Child Health History Questionnaire. Cognitive assessments were conducted using the NIH Toolbox Cognition Battery (NTCB; Weintraub et al., 2013). The NTCB is a computer-based assessment tool with seven tests measuring cognitive performance in the domains of attention, EF, processing speed, working memory, episodic memory and language. Additional information on the NTCB can be found at http://www.nihtoolbox.org/. Cognitive tasks were selected based on evidence suggesting relationships with impulsivity (Hoaken et al., 2003; Romer et al., 2009). The DCCS score and the Attention and total scores from the Flanker Inhibitory Control and Attention Test were used as measures of EF and were predicted to be related to NegUrg. The total score from the Oral Reading Recognition Test (a measure of overall reading ability) and the total vocabulary score from the Picture Vocabulary Test (measure of vocabulary knowledge) were used as measures of crystallised intelligence and were predicted to be less related to NegUrg (Spinella, 2004).
To examine psychiatric and affective constructs, assessments were conducted using the PhenX Toolkit (https://www.phenxtoolkit.org). The Total Anxiety Score from the Screen for Child Anxiety Related Emotional Disorders (SCARED; Birmaher et al., 1999) and the Negative and Positive Affect Scores from the Positive and Negative Affect Schedule for Children (PANAS-C; Laurent et al., 1999) were included in the current study.
Model II: brain volumes of regions of interest
A standardised acquisition protocol for structural neuroimaging data was utilised at nine PING study sites across the United States (Jernigan et al., 2016). Scanner type (3T GE, Philips or Siemens) varied by site. T1-weighted scans were used for automated cortical and subcortical segmentation using FreeSurfer software (https://surfer.nmr.mgh.harvard.edu/). Cortical volumes (cubic millimetres) were calculated for each brain region using the Desikan–Killiany parcellation atlas, which includes a total of 34 regions of interest (ROIs) per hemisphere (Desikan et al., 2006). Additional information on the specific PING protocols for each scanner manufacturer can be found at http://pingstudy.ucsd.edu/resources/neuroimaging-cores.html. Ten ROIs (five regions of homologous pairs from both hemispheres) were chosen for analysis based on their relevance to emotion regulation or cognitive control and were predicted to be related to NegUrg (see Table 2; Boes et al., 2009; Chester et al., 2016; Cyders et al., 2015).
Model III: genetic data
Genetic information was collected via saliva samples and derived through genome-wide sequencing using the Illumina Human660W-Quad BeadChip, which contains over 550,000 genetic markers (including SNPs and copy number variations); the Illumina Human660W-Quad BeadChip was designed on Hapmap release 21 reference data (see http://hapmap.ncbi.nlm.nih.gov/; additional information can be found at https://www.ncbi.nlm.nih.gov/variation/news/NCBI_retiring_HapMap/). Genotyping assignments were made, and reproducibility was examined using a clustering algorithm in Illumina’s Genome Studio software. SNPs with <99.9% reproducibility were flagged and investigated further (Jernigan et al., 2016). For this project, quality control was conducted on the final genetic dataset, including filtering for minor allele frequency <1% and Hardy–Weinberg Equilibrium, p < 0.0001. Additional information on the specific PING genetics protocol can be found at http://pingstudy.ucsd.edu/resources/genomics-core.html. We selected 265 ACGT-coded SNPs from nine genes associated with dopaminergic transmission and metabolism pathways, language development and psychopathology using PLINK (Version 1.07; Purcell et al., 2007) for model building (see Table 3).
SNPs organised by gene and predictive function.

SNPs: single nucleotide polymorphisms.
Statistical analyses
The study sample was divided into two subsets: (1) an initial model-building sample (80%) for training and cross-validation and (2) a test sample (20%) to check the external validity of the final model. Statistical analyses were conducted in R Studio (Version 1.0.44; R Core Team, 2016) using best subset and least absolute shrinkage and selection operator (LASSO) regressions (glmnet package; Tibshirani, 1996). Each set of variables was used to predict the criterion, NegUrg. R2 and mean squared error (MSE) were examined to evaluate model fit. Comparison of resultant R2 and MSE values from the model-building and test samples was used as an indicator for the validity and generalisability of the final models. Beta weights were used to evaluate the contribution of individual predictors.
LASSO regression
Separate LASSO regressions were conducted on Model I and on Model III. The LASSO method of variable selection was chosen due to its ability to yield sparse models in which predictors can be ‘zeroed’ out of the model. Ten-fold cross-validation was used to optimise a single regularisation parameter (lambda).
Best subset regression
Best subset regression was conducted on Model II. The best subset method of variable selection was chosen to derive the most robust and useful combination of predictors in the context of a more limited array of predictors. Separate least squares regressions were performed for each possible combination of predictors, with subset sizes ranging from a single predictor to 10, the full number of predictors. Test error was estimated using 10-fold cross-validation for each possible number of predictors. The model for which the number and set of predictors minimised the mean cross-validation error was chosen.
Results
Model I: sociodemographic, medical history, cognitive and affective data
Seven cognitive/demographic predictors were selected through LASSO regression. Fit indices for the model-building sample (MSE = 0.35, R2 = 0.25) and the test sample (MSE = 0.35, R2 = 0.30) were similar, indicating that the model fit was appropriate. Non-zeroed coefficients for the selected predictors are presented in Table 4.
Model I non-zeroed coefficients.

PANAS-C: Positive and Negative Affect Schedule for Children; SCARED: Screen for Child Anxiety Related Emotional Disorders.
A number of affective, behavioural and demographic variables predicted NegUrg. The most efficient model included affective variables but neither fluid (i.e. measures of EF) nor crystallised cognitive ability. The strongest positive predictors of NegUrg were self-reported total anxiety, as well as negative affect and behaviour problems in school. NegUrg declined with age, while higher household income and mother’s alcohol use during pregnancy were associated with higher NegUrg. Our model indicated an association between male gender and lower risk of self-reported NegUrg. Females (M = 2.36, SD = 0.64) had 9.31% higher NegUrg scores than males (M = 2.15, SD = 0.67).
Model II: ROI data
Best subset regression revealed that the optimal (lowest mean cross-validation error) model consisted of three right-sided ROIs (right caudal anterior cingulate, right posterior cingulate and right rostral anterior cingulate) which best predicted NegUrg. Fit indices for the model-building sample (MSE = 0.41, R2 = 0.04) and the test sample (MSE = 0.47, R2 = 0.05) were similar, indicating that the model fit was appropriate. Coefficients for the selected predictors are presented in Table 5. Greater volume of the right posterior cingulate and right rostral anterior cingulate was associated with lower self-reported NegUrg, while greater volume of the right caudal anterior cingulate was associated with higher self-reported NegUrg.
Model II non-zeroed coefficients of selected ROIs.

ROIs: regions of interest.
Model III: genetic data
Three SNPs from two genes were selected through LASSO regression. Fit indices for the model-building (MSE = 0.43, R2 = 0.16) and the test samples (MSE = 0.38, R2 = 0.11) were similar, indicating that the model fit was appropriate. The final model indicated that rs17022545 and rs9829960 from CADM2, as well as rs3794808 from SLC6A4, were predictive of NegUrg. Non-zeroed coefficients for the selected predictors are presented in Table 6.
Model III non-zeroed coefficients of selected SNPs.

SNPs: single nucleotide polymorphisms.
Follow-up a posteriori analyses
To further examine the extent of the relationship between variables that were zeroed out of the final models and NegUrg, two follow-up LASSO analyses were conducted. First, because the measures of EF (DCCS, Flanker) were expected to be predictive, but were not selected in the LASSO regression results, a subset of the Model I variables was used in a follow-up LASSO regression (see Table 7). Results of this analysis zeroed out all of the coefficients entered, again indicating that they were not predictive in the context of a model without all the variables in the original LASSO regression.
Subset of Model I variables for follow-up LASSO.

LASSO: least absolute shrinkage and selection operator; ADHD: attention-deficit/hyperactivity disorder; DCCS: Dimensional Change Card Sort.
Second, the left hemisphere ROIs from Model II were examined, as they were not selected by the best subset regression in the original analysis. Variables entered into the model are presented in Table 8. Best subset regression revealed that the optimal (lowest mean cross-validation error) model consisted of two left hemisphere ROIs out of the five entered into the model (MSE = 0.45). However, the model-building sample produced an R2 = 0.021 and the test sample of that model produced an R2 = 0.0036, indicating that the model fit might not generalise. Taken together, our neuroanatomical results suggest that right cingulate ROIs represent the most efficient neural architectural model to predict NegUrg.
Subset of Model II variables for follow-up best subset.
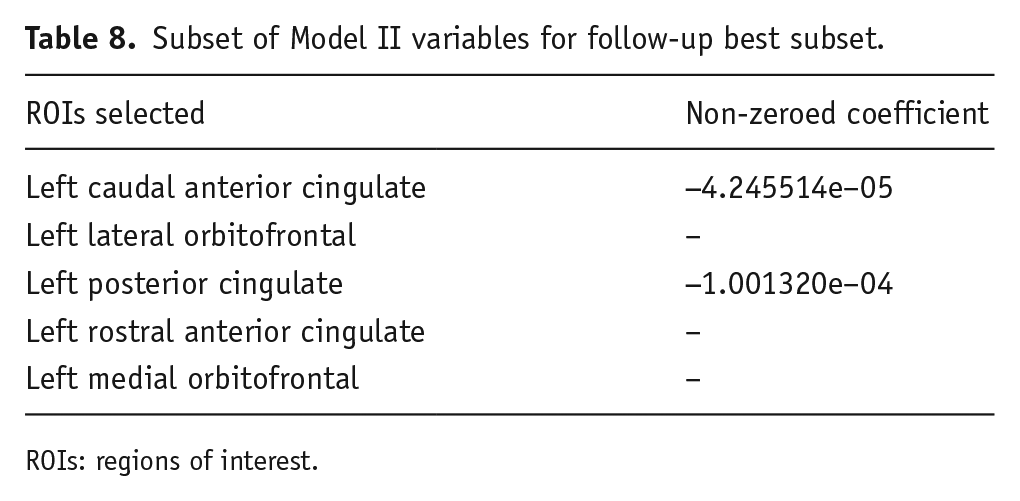
ROIs: regions of interest.
Discussion
Results demonstrate that data from an array of domains including brain and neurogenetic mechanisms can be harmonised in a community-dwelling sample with relevance to developmental psychopathology research. Sociodemographic factors, as well as psychological, genetic and right hemisphere fronto-limbic neuroanatomical variables related to emotional regulation (as opposed to cognitive control), predicted NegUrg in a cross-sectional, childhood to early adulthood, sample. Results support construct and criterion validity of the UPPS-P NegUrg Scale and elucidate the developmental bases of the NegUrg component of impulsivity, which has been linked to neuropsychiatric and externalising disorders and academic underachievement (Muhlert and Lawrence, 2015; Spinella, 2004).
NegUrg implies difficulties with decision-making and emotion regulation, suggesting a role for cognitive (i.e. executive) dysfunction in the development of impulsivity. However, EF measures (Flanker, DCCS, etc.) were not selected in our analyses, emphasising emotional and behavioural factors (i.e. emotional regulation vs. low cognitive ability) in NegUrg aetiology. Interestingly, age may contribute to differential ACC recruitment, as older children recruited dorsal (‘cognitive’) while younger children recruited ventral (‘emotional’) areas during emotional regulation (Perlman and Pelphrey, 2010).
Self-reported anxiety, negative affect and school behaviour problems were the strongest NegUrg predictors. Findings support previous research demonstrating NegUrg’s negative relationship to emotional regulation and intersection of impulsivity and negative affect (Cyders and Smith, 2008; Whiteside and Lynam, 2001). Previous research identified an association between anxiety and ACC independent of age, including preferential recruitment of ventral regions in more fearful children (Perlman and Pelphrey, 2010). During processing of high arousal stimuli, NegUrg has been negatively associated with activation in ACC and anterior medial OFC, regions associated with emotional control (Joseph et al., 2009). The distinct emotional dysregulation characterisation of NegUrg found in our study is consistent with the five UPPS-P factors contributing unique components of impulsivity (Lynam et al., 2006), as well as NegUrg being related to NEO-Five Factor Inventory’s Neuroticism dimension but not Conscientiousness or Extraversion (Whiteside and Lynam, 2001). NegUrg is associated with maladaptive behaviours and psychopathology associated with problematic risk-taking and disinhibitory behaviours, including alcoholism, borderline personality disorder, and dysregulated eating disorders (Anestis et al., 2009; Coskunpinar et al., 2013; Cyders and Smith, 2008; Fischer et al., 2008; Smith and Cyders, 2016). Relative UPPS-P elevations and predictors revealed here may help identify at-risk youth for early intervention.
Predictors of NegUrg included age, gender, household income, mother’s alcohol use during pregnancy, school behaviour problems, anxiety and negative affect. Older participants had lower NegUrg, consistent with known developmental trajectory (Cyders and Smith, 2008; Green et al., 1999) and brain development processes (Romer, 2010). From approximately the age of 11 into early adulthood, the thinning of cortical grey matter has been associated with behavioural changes, including increased impulsivity (Romer, 2010), as well as improved information processing efficiency, a component of EFs (Breukelaar et al., 2017). Similarly, the association between NegUrg and behavioural problems in school is consistent with existing research showing the relationship between NegUrg and externalising behaviours, such as disordered conduct and problematic alcohol use (Settles et al., 2012).
Our model indicated that female gender predicted greater NegUrg (females had 9% higher scores); similar results have been demonstrated for the impulsiveness facet of the NEO-Personality Inventory-Revised (Costa et al., 2001), but this finding is not entirely consistent with previous work (Cyders, 2013). Socialisation and sociocultural factors may contribute to gender differences of emotional expression, with girls demonstrating more internalising emotions than boys (Chaplin and Aldao, 2013). The role of emotional expression and socialisation in NegUrg presents an avenue for further research.
Our results link higher parental family income, a component of SES, to increased NegUrg. This finding is in contradiction to our prediction that low SES would be associated with higher NegUrg, based on the relationship between poor EF and low SES (Arán-Filippetti et al., 2012). It is possible that children from higher SES families may be under greater emotional stress due to increased pressure to perform academically; however, future research should parse apart relationships between specific components of SES and more cognitively versus emotionally mediated forms of impulsivity.
Our results indicated that in a healthy sample, maternal alcohol use during pregnancy is associated with higher NegUrg. These results support previous findings that maternal alcohol consumption may confer behavioural risk (e.g. child behavioural problems, symptoms of attentional and impulse disorders) for offspring, even at non-clinical levels (Flak et al., 2014). These results highlight the importance of maternal alcohol use as a contributing factor to preclinical traits, such as NegUrg.
Our results show an association between the rostral subregion of the right ACC with NegUrg. This is consistent with previous findings examining neural correlates of NegUrg and related psychopathology (e.g. Boes et al., 2009; Fjell et al., 2012; Hoptman et al., 2014). Research shows associations of NegUrg and PFC suggestive of developmental psychopathology and is broadly consistent with a right hemisphere lateralisation of inhibitory processes (Aron et al., 2004). Furthermore, our results indicate the cingulate gyrus is important for the neural representation of NegUrg and support the existence of a normal-to-pathological continuum for the relationship between individual variation in cortical morphology and self-regulatory problems (Fjell et al., 2012). Nevertheless, the picture that emerges across studies is nuanced. While Boes et al. (2009) interpreted the reduced volume in the right ventromedial prefrontal cortex to indicate cortical difficulties in regulation of hedonic subcortical input, the positive and negative associations of cortical volume to NegUrg in this investigation suggest the literature will need to develop to embrace more complex relationships.
Consistent with previous research demonstrating a genetic association with impulsivity and related constructs, including brain development (Khadka et al., 2014), our results indicated that the rs17022545 and rs9829960 SNPs from CADM2 (encoding cell adhesion molecule 2) were predictive of higher and lower NegUrg, respectively. CADM2 is associated with synaptic organisation and has been linked to individual differences in processing speed (Ibrahim-Verbaas et al., 2016). It has been identified as a candidate gene for autism spectrum disorder, given its role in synapse formation in early postnatal development (Casey et al., 2012), and has been linked to hyperactivity and impulsivity in a sample of children and adolescents, aged 6 to 18 years (Albayrak et al., 2013). Consistent with our neuroanatomical findings, CADM2 is expressed more robustly in frontal and ACC regions (Ibrahim-Verbaas et al., 2016). These results link neurodevelopment and individual differences in impulsive behaviour (Khadka et al., 2014).
Our results also indicated that the rs3794808 SNP from SLC6A4 (encoding the 5HT transporter Solute Carrier Family 6 Member 4) was related to lower NegUrg. The SLC6A4 gene encodes an integral membrane protein responsible for cross-membrane transport of 5HT, a neurotransmitter that contributes to emotion-based action, including urgency (Cyders and Smith, 2008). The role of DA and 5HT pathways across several forms of impulsivity reflects functional heterogeneity, with an emerging trend for association of reduced 5HT levels and premature response tendency (Dalley and Roiser, 2012). Greater expression of SLC6A4 in the brain has been demonstrated in areas representing emotional aspects of behaviour, such as cortical and limbic areas (Lesch et al., 1996). A polymorphic region of SLC6A4, 5-HTTLPR, has also been associated with anxiety-related personality traits, as well as the Harm Avoidance trait on the Tridimensional Personality Questionnaire (Lesch et al., 1996). Taken together, these factors provide support for a genetic basis to NegUrg that is in line with the sociodemographic and neuroanatomical findings of the construct.
Limitations/future directions
The cross-sectional nature of the study did not permit an evaluation of the interaction between developmental phase and NegUrg prediction. Our participants’ ages span three crucial neurodevelopmental periods, making it difficult to interpret associations in cortical volume of adjacent brain regions in a developmental sample, as stage of brain maturation likely has implications.
UPPS-P is a self-report measure of NegUrg, which may have different correlates than a behavioural measure (Whelan et al., 2012), particularly in a community-dwelling sample. Nevertheless, self-reported NegUrg is associated with a range of psychopathology (e.g. Berg et al., 2015); in our sample, it was predicted by school behaviour problems, negative affect and anxiety. The model using ROIs indicated that right hemispheric regions best predicted NegUrg, consistent with research linking fearful temperament, emotion-regulation impairment and right-lateralised frontal electroencephalogram (EEG) asymmetry (Fox et al., 1995). Future research should examine brain laterality of NegUrg, given the LASSO method of zeroing out variables that may have predictive value but do not fit sparse models. The role of the genetics of NegUrg, including both dopaminergic and serotonergic genetic pathways, remains an avenue for future work. While research involving SNPs has increased understanding of the interaction between genetics and phenotypes, the use of SNPs has limitations. SNPs do not necessarily tell us about expression levels or functional differences for a particular protein or gene. Therefore, it is difficult to understand the functional impact of a specific SNP. For example, it is not clear exactly how the polymorphisms in SLC6A4 relate to 5HT transport or 5HT levels within synaptic compartments, or how those levels relate to impulsivity. Furthermore, each region of the brain has specific transcription factors and protein modification systems, so regionally specific implications of individual SNPs cannot be ascertained from genotyping alone.
Despite these limitations, this study furthers understanding of the NegUrg construct and provides additional support for its intersectionality between impulsivity and negative affect. Although results do not negate the relevance of variables not selected in the models, they point to salience of the affective, neuroanatomical and genetic variables that were selected in typically developing individuals from childhood to early adulthood. The combination of increased negative affect, heightened anxiety and school behavioural problems, as well as areas associated with emotional processing in the right hemisphere, and genetic factors linked to impulsivity and anxiety appear to be of particular importance and may serve as markers for early NegUrg identification.
Footnotes
Authors’ note
Data used in preparation of this article were obtained from the Pediatric Imaging, Neurocognition and Genetics (PING) study database (http://ping.chd.ucsd.edu). As such, the investigators within PING contributed to the design and implementation of PING and/or provided data but did not participate in analysis or writing of this report. A complete listing of PING investigators can be found at ![]() .
.
Author contributions
We attest that all authors made meaningful scientific contributions to the investigation.
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: Data collection and sharing for this project was funded by the Pediatric Imaging, Neurocognition and Genetics (PING) study (National Institutes of Health Grant RC2DA029475). PING is funded by the National Institute on Drug Abuse and the Eunice Kennedy Shriver National Institute of Child Health and Human Development. PING data are disseminated by the PING Coordinating Center at the Center for Human Development, University of California, San Diego. The content is solely the responsibility of the authors and does not necessarily represent the official views of the National Institutes of Health, the US Department of Veterans Affairs or the US Government.