
Abstract
The identification, in the late 20th century, of unexpectedly large families of G-protein-coupled chemosensory receptors revolutionised our understanding of the olfactory system. The discovery that non-selective olfactory sensory neurons express a single olfactory receptor type and project to a specific glomerulus in the main olfactory bulb provided fundamental insight into the spatial pattern of odour representation in the main olfactory bulb. Studies using head-fixed awake mice and optogenetics have revealed the importance of the timing of glomerular input in relation to the sniff cycle and the role of piriform cortex in odour object recognition. What in the 1970s had appeared to be a relatively simple dichotomy between odour detection by the main olfactory system and pheromone detection by the vomeronasal system has been found to consist of multiple subsystems. These mediate innate responses to odours and pheromones and to substances as diverse as O2, volatile urinary constituents, peptides and proteins.
The neurobiology of the sense of smell was a neglected research area for most of the 20th century, partly due to the perceived lack of importance of clinical disorders. However, a more fundamental reason for the lack of progress was the difficulties in organising, classifying and controlling olfactory stimuli. Other sensory stimuli can be easily varied along linear dimensions, such as spatial location, wavelength, frequency or intensity. But how can you organise complex molecular structures with differing sizes, shapes and multiple functional groups? How can the receptive field of an olfactory neuron be mapped in response to thousands of possible odorant stimuli? Moreover, the timing of stimulus delivery is less precise than for other senses, and olfactory stimuli are comparatively difficult to control. All these factors meant that by the 1980s, although there had been many attempts to characterize odorants, according to a limited number of primary odour characteristics (Amoore, 1963), none of these attempts to simplify odour space was successful and progress on understanding olfaction was slow.
Identification of olfactory receptor genes
By the later 1980s, progress had been made in identifying the transduction mechanism in olfactory sensory neurons (OSNs). It was known that odour stimulation was associated with guanosine triphosphate (GTP)-dependent increases in intracellular cyclic adenosine monophosphate (cAMP), suggesting the involvement of G-protein coupled receptors (GPCRs) (Pace et al., 1985). But the big breakthrough in the field came with the discovery of the olfactory receptor (OR) gene family by Buck and Axel (1991), work which subsequently led to the award of the 2004 Nobel Prize for Physiology and Medicine. Buck and Axel searched for GPCRs with diverse sequences and with expression restricted to the olfactory epithelium. What they found was the largest GPCR gene family in the mammalian genome, with the latest genomic studies suggesting that around 1100 functional ORs are expressed in mice (Saraiva et al., 2015) and around 370 functional ORs expressed in humans. This identification of the OR gene family paved the way for the discovery of a fundamental organising principle of the olfactory system that a mature OSN monoallelically expressed a single OR type (Chess et al., 1994). Yet despite expressing only a single OR type, OSNs typically responded to a range of structurally related odorants, and conversely, each individual odorant was found to activate OSNs expressing different OR types (Malnic et al., 1999). The combinatorial nature of this coding strategy means that the number of different odorants that can be detected and discriminated vastly exceeds the number of different OR types.
Is there a neural map of odorant responses in the main olfactory bulb?
The next step was the genetic labelling of OSNs expressing a specific OR gene. This revealed that OSNs expressing a specific OR were scattered widely across an expression zone in the main olfactory epithelium. But that axons of OSNs expressing the same OR converged onto individual glomeruli that received input solely from that specific OR type (Figure 1; Mombaerts et al., 1996). Various imaging techniques have since confirmed the basic coding principle that individual glomeruli respond to multiple odorants, but the overall pattern of glomerular activity is unique for each odorant (Friedrich and Korsching, 1997).

Basic principles of olfactory information processing. Olfactory sensory neurons expressing a particular olfactory receptor type (shown by colour) are distributed in the main olfactory epithelium, but converge their axonal projections to a specific glomerulus in the main olfactory bulb. Activity-dependent lateral inhibition between non-neighbouring mitral cells can enhance contrast in the spatiotemporal pattern of mitral cell activity.
Imaging studies from the late 1970s onwards had suggested that the pattern of glomerular response to an odorant was relatively stereotyped, both between right and left main olfactory bulbs (MOBs) within an individual and between individuals of the same species. This raised the question of whether odour information was mapped on the surface of the MOB in a similar way to the neural mapping of stimulus properties in other sensory modalities. The emerging view by the turn of the century was that there was a broad chemotopic mapping, with glomerular responses to different classes of odorants mapping to different regions of the MOB (Mori et al., 2006). But when individual glomeruli were identified on the basis of their odorant response profile, it was found that glomeruli that responded to similar odorants were intermingled with glomeruli that responded to completely different classes of odorant (Soucy et al., 2009). This suggested that the large-scale chemotopic map across the MOB is not apparent at the level of individual glomeruli and is unlikely to perform the same role in neural processing as the neural maps in other sensory modalities. Instead, the highly stereotyped glomerular position could be explained as a developmental map in which the position of glomerular convergence on the MOB depended on variations in basal, unstimulated G-protein activity among OR types (Imai et al., 2006). This developmental principle explained how ORs sharing a high level of sequence homology have similar odorant binding characteristics and similar basal levels of G-protein activation, which resulted in them converging to similar areas of the MOB. This stereotyped, self-organising pattern of glomerular convergence may be important in enabling developing OSN axons to converge onto the correct glomerulus during development and during the continuous process of re-innervation in the adult.
How is the olfactory code decoded?
A unique feature of the vertebrate olfactory bulb, which has long intrigued neuroscientists, is the presence of reciprocal synapses between the MOB projection neurons and inhibitory interneurons (Figure 1). These inhibitory connections undoubtedly play a variety of roles in shaping the spatiotemporal pattern of mitral cell responses to odours, including the fast synchronisation of mitral cell ensembles. However, these inhibitory connections are not as dense or spatially uniform as those mediating lateral inhibition in other sensory systems, which raised the question of how such sparse and irregular inhibitory connections could play a role in neural processing. A potential explanation for this organisation has come from recent evidence that reciprocal synapses can mediate activity-dependent lateral inhibition. This could perform contrast enhancement of the pattern of mitral cell activity, irrespective of their local spatial organisation (Arevian et al., 2008), and therefore overcome the lack of spatial mapping of stimulus features across the MOB. Furthermore, pattern separation of newly learned odours in the MOB may be enhanced by neurogenesis and the incorporation of new inhibitory interneurons, with more plastic synapses, into the MOB network (Lledo and Valley, 2016).
By the 1980s, our understanding of the role of the piriform cortex in olfaction was based largely on anatomical tracing studies. These revealed that individual mitral cells send divergent projections to pyramidal cells across large areas of piriform cortex. Conversely, piriform cortex pyramidal cells receive input from mitral cells sampling different OR inputs from glomeruli across the MOB. The neural architecture of the anterior piriform cortex and its convergent pattern of inputs suggested that it could correlate the odourant features represented at the level of the MOB and associate them into representations of odour objects. Direct evidence for this hypothesis was eventually provided by recording responses from individual pyramidal cells in piriform cortex while optically stimulating different patterns of glomeruli in the olfactory bulb. Not only did multiple glomeruli need to be activated to cause spiking in piriform cortex pyramidal cells, but different individual pyramidal cells were activated by different patterns of glomerular activity (Figure 2; Davison and Ehlers, 2011).
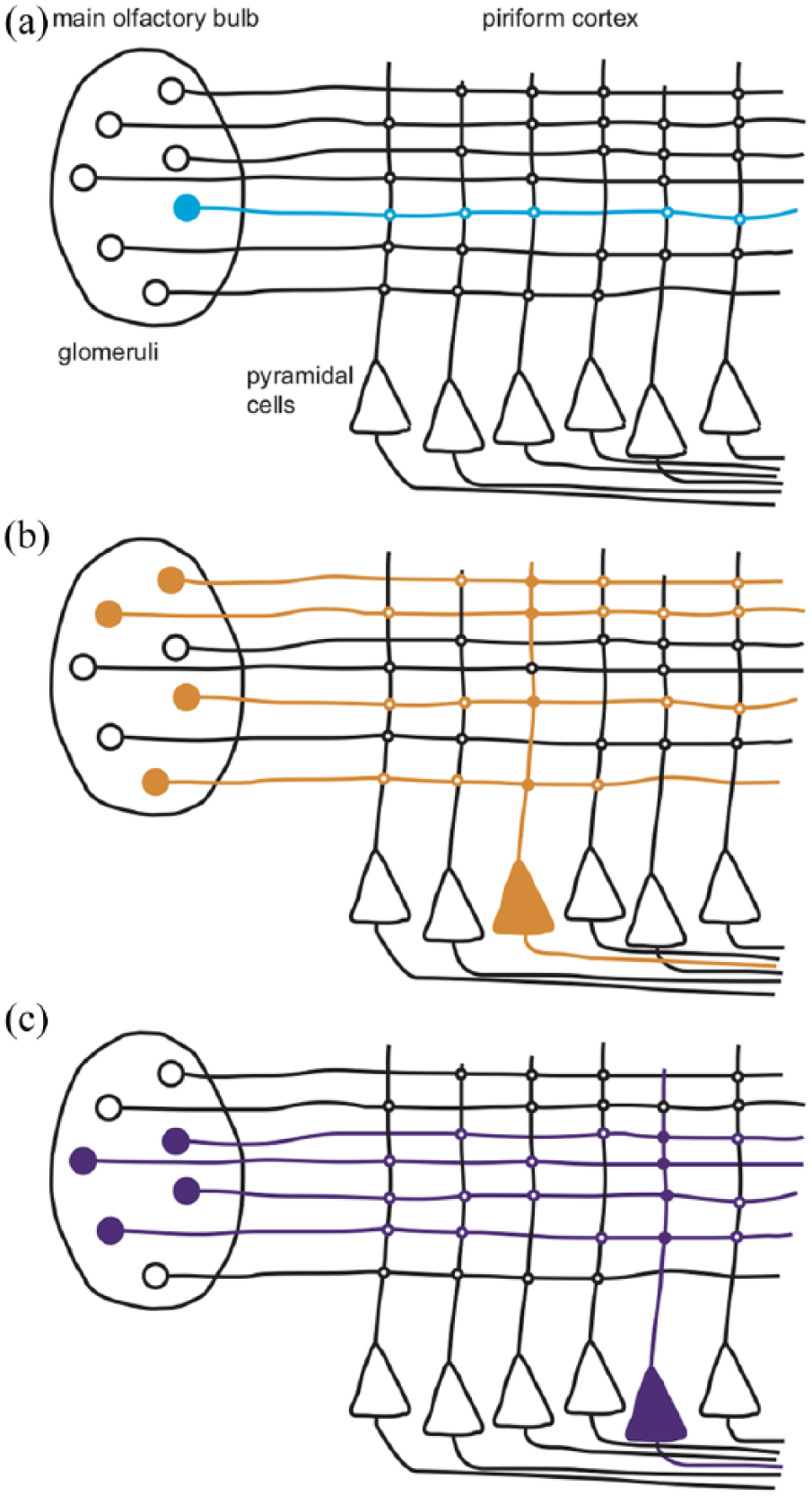
Pyramidal neurons in the piriform cortex associate patterns of mitral cell activity to form odour objects. (a) Stimulation of a single glomerulus is not sufficient to drive pyramidal neuron activity. (b) Simultaneous stimulation of at least four glomeruli drives pyramidal neuron activity. (c) Different patterns of glomerular stimulation activate different pyramidal neurons.
Is there a temporal dimension to the odour code?
One of the major advances over the last couple of decades has been the introduction of the head-fixed awake recordings in mice. As well as avoiding the direct effects of anaesthesia during electrophysiological recordings, this approach enabled animals to engage in active odour sampling via sniffing. This revealed that mitral cell responses from awake mice appeared to be more sharply tuned to a narrower range of odorants, compared to recordings performed under anaesthesia. Moreover, when mitral cell spike trains were aligned to the sniff cycle rather than to stimulus delivery, the peak firing rate was found to be higher and more precisely timed. Peak responses occurred at different phases of the sniff cycle in the same mitral cell in response to different odours and at different times in different mitral cells in response to the same odour (Shusterman et al., 2011). Along with earlier studies using voltage-sensitive dye imaging, these findings demonstrated that the spatial pattern of glomeruli and mitral cells evolves during the odour response.
However, the question of whether the olfactory system could use this temporal information to provide a further dimension for discrimination of similar odours could not be addressed using odour stimuli, as they are too dispersed in time for studies requiring millisecond precision. Smear et al. eventually overcame this problem using optogenetic activation of OSN input to the MOB, which could be delivered at precise times in relation to the sniff cycle. They showed that mice could be trained to reliably discriminate the same optogenetic stimulus delivered at different times during the sniff cycle, even when the timing difference was as short as 25 ms (Smear et al., 2011). The importance of timing could also be observed at the level of the piriform cortex, where pyramidal neurons appeared to be tuned to respond to certain lags of glomerular activation and therefore potentially capable of converting the MOB temporal code into a frequency code (Haddad et al., 2013).
What does the vomeronasal system do?
The term pheromone was coined over 50 years ago to describe a substance released by an individual that has an innate effect on another individual of the same species. Pheromonal effects on mouse female reproductive state had been established by 1970. These were found to be mediated by the vomeronasal organ (VNO), which was regarded as a parallel and independent chemosensory system to the main olfactory system. However, progress in identifying potential pheromonal effects in vertebrates was slow, relying on behavioural bioassays to identify active constituents that were typically present at low concentrations in complex biological secretions. Once again, it was the identification of the sensory receptors that transformed the field and that has led to the rapid increase in understanding of the mouse vomeronasal system over the last 20 years.
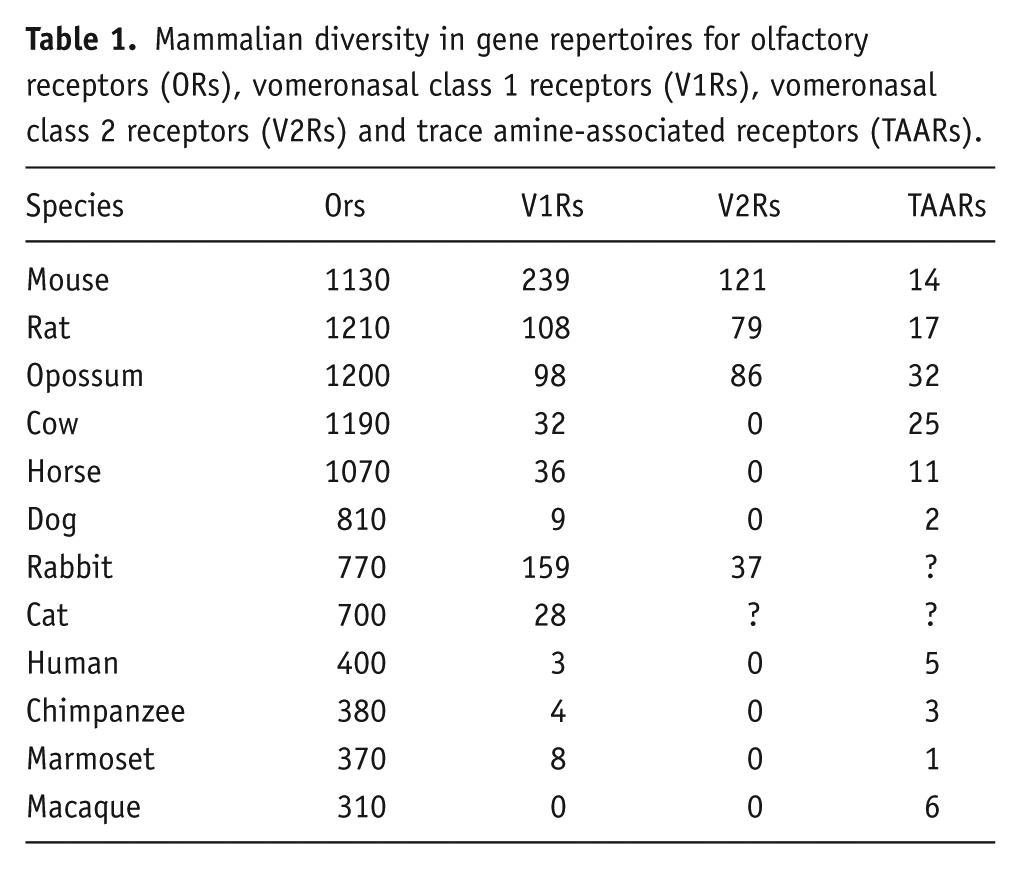
In the mid 1990s, Axel’s group used differential screening of complementary DNA (cDNA) libraries from individual vomeronasal sensory neurons to identify the V1R class of vomeronasal receptors (Dulac and Axel, 1995). A year later, Buck’s group used a similar approach to identify a completely separate class of V2R vomeronasal receptors (Matsunami and Buck, 1997). Recent analysis of the mouse genome has revealed around 239 functional Vmn1rs, which detect small volatile molecules and sulphated steroids; 121 functional Vmn2rs, which detect proteins and peptides and a further five formyl peptide receptors that detect bacterial and immune-related peptides. This unexpectedly large number of receptor types suggests a previously unsuspected level of complexity in the role of the mouse VNO mediating the effects of pheromones and conveying information about conspecifics, including their sex, hormonal status, genetic individuality and infection status. However, this is not typical of most mammals and considerable species differences exist (Table 1).
Mammalian diversity in gene repertoires for olfactory receptors (ORs), vomeronasal class 1 receptors (V1Rs), vomeronasal class 2 receptors (V2Rs) and trace amine-associated receptors (TAARs).
For most of the 1990s, the commonly held view was that there was a clear dichotomy between the function of the main olfactory system to detect general odours and the vomeronasal system to detect pheromones. However, the mouse VNO has since been found to mediate not only pheromonal effects but also innate responses to predator odours (Table 2; Isogai et al., 2011). Moreover, chemosensory subsystems have been found within the main olfactory system, such as OSNs expressing trace amine-associated receptors, which mediate innate responses to chemosensory stimuli, including predator odours and pheromones (Dewan et al., 2013). The main distinction between these two systems now appears to be that the main olfactory system is adapted to respond to airborne stimuli, whereas the vomeronasal system is adapted to respond to stimuli in the liquid phase following direct contact with the stimulus source.
Mouse olfactory subsystems, their receptors and their functional significance.

Looking forward
What appeared, 50 years ago, to be a relatively simple, if mysterious, sense has been revealed as a far more complicated set of sensory receptors and pathways, mediating both innate and learned responses to chemosensory cues. There is still an uncharted world of chemosignals and receptors to be explored, along with central olfactory areas such as the nucleus of the lateral olfactory tract and olfactory tubercle about which very little is known, apart from their anatomical connectivity. But we now have the experimental tools to ensure that the recent fast pace of progress of our understanding of olfaction will not be slowing down anytime soon.
Footnotes
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship and/or publication of this article.
Funding
The author(s) received no financial support for the research, authorship and/or publication of this article.