
Abstract
Study Design
Translational animal study.
Objective
To investigate the therapeutic effects of needle-knife therapy in cervical spondylotic radiculopathy (CSR) rat models. This study also aims to determine whether these effects are mediated via inositol-requiring enzyme 1 alpha (IRE1α)-X-box binding protein 1 (XBP1) axis.
Methods
A CSR rat model was established via spinal canal insertion. The IRE1α activator IXA4 was administered to enhance IRE1α-XBP1 signaling activity. Motor function and pain sensitivity were evaluated using paw withdrawal mechanical threshold (PWMT), paw withdrawal thermal latency (PWTL), and gait scoring. Microglial activation was evaluated by immunofluorescence staining for ionized calcium-binding adapter molecule 1 (Iba1) and tumor necrosis factor-alpha (TNF-α). Neuronal apoptosis was assessed via TUNEL/NeuN double immunofluorescence, qRT-PCR, and Western blotting (WB). Key proteins in the IRE1α-XBP1 signaling were analyzed using WB.
Results
CSR rats exhibited spinal cord inflammation, neuronal apoptosis, and aberrant activation of the IRE1α-XBP1 pathway. Needle-knife therapy significantly mitigated spinal cord pathology. The treatment suppressed inflammatory cytokines, inhibited pro-inflammatory microglial activation, and inhibited activation of IRE1α-XBP1 signaling. Administration of IXA4 partially reversed these protective effects.
Conclusion
Needle-knife therapy ameliorates inflammation and neuronal apoptosis in CSR. These effects are associated with downregulation of the IRE1α-XBP1 signaling.
Keywords
Introduction
Cervical spondylotic radiculopathy (CSR) is one of the most prevalent subtypes of cervical spondylosis, characterized by compression or irritation of cervical nerve roots. 1 Clinically, CSR manifests with neck and shoulder pain, radiating upper limb numbness, sensory abnormalities, and motor dysfunction, significantly impairing patients’ quality of life. 2 Growing evidence suggests that inflammation plays a central role in the pathophysiology of CSR. 3 Mechanical compression of nerve roots triggers an inflammatory cascade, marked by increased production of pro-inflammatory cytokines such as tumor necrosis factor-alpha (TNF-α) and interleukin-6 (IL-6), which aggravate neural tissue injury and contribute to pain sensitization. 4 In parallel, neuronal apoptosis has been identified as a critical pathological event in CSR progression, where excessive cell death in spinal cord and dorsal root ganglion neurons exacerbates neurological deficits and delays functional recovery. 5
Conventional treatments for CSR include pharmacological therapies (eg, non-steroidal anti-inflammatory drugs, corticosteroids), physical rehabilitation, and surgical interventions. However, these approaches often have limited long-term efficacy or are associated with notable side effects and complications.6,7 Therefore, alternative or complementary therapies with both symptom-relieving and neuroprotective properties are urgently needed. Needle-knife therapy is a minimally invasive intervention derived from traditional Chinese medicine that combines the principles of acupuncture and modern surgical techniques. It involves releasing soft tissue adhesions and improving local circulation by inserting a specialized blade-like needle into targeted sites. 8 In recent years, needle-knife therapy has been widely applied in the treatment of musculoskeletal and cervical spine disorders, including CSR, with demonstrated clinical efficacy in reducing pain and improving mobility. 9 Despite these promising results, the underlying molecular mechanisms of its therapeutic effects remain largely unexplored.
Among the intracellular signaling pathways implicated in neuroinflammation and apoptosis, the inositol-requiring enzyme 1 alpha (IRE1α)-X-box binding protein 1 (XBP1) axis has emerged as a key regulator. IRE1α, an endoplasmic reticulum (ER) stress sensor, initiates an adaptive response via unconventional splicing of XBP1 mRNA under pathological conditions. 10 Activation of the IRE1α-XBP1 pathway is closely associated with pro-inflammatory cytokine production and neuronal apoptosis in various models of spinal cord injury and neurodegeneration.11,12 Although the IRE1α-XBP1 signaling axis has been studied before, its involvement in CSR pathogenesis and therapeutic modulation by needle-knife therapy has not been systematically investigated.
Given this background, we hypothesize that needle-knife therapy exerts its beneficial effects in CSR by modulating the IRE1α-XBP1 signaling to affect inflammation and neuronal apoptosis. This study aims to explore the protective role and underlying mechanism of needle-knife therapy in CSR rat models, with a particular focus on the regulation of the IRE1α-XBP1 axis. The findings may offer new insights into the molecular basis of needle-knife therapy and provide experimental evidence to support its application in clinical management of CSR.
Materials and Methods
Experimental Animals
A total of 48 specific pathogen-free-grade male Sprague-Dawley (SD) rats weighing 200 ± 10 g were purchased from SJA Laboratory Animal company (Changsha, China; Approval No. SCXK [Xiang] 2025-0004). Rats were housed under standard laboratory conditions with a temperature of 22-25°C, relative humidity of 50-60%, and a 12 h light/dark cycle, with ad libitum access to food and water. All animal procedures were approved by the Ethics Committee of Hunan University of Chinese Medicine (Ethics Number: HNUCM21-2512-17) and were conducted in accordance with institutional guidelines.
Needle-Knife Therapy
Following model establishment, needle-knife therapy was performed at five acupuncture points: the interspinous spaces of the left C6, C7, and T1 vertebrae, and bilateral Jiaji points (located approximately 0.5 cm lateral to the midline spinous processes of the cervical vertebrae). A Guxi-branded needle-knife (0.35 × 25 mm, Bond Medical Instruments Co., Ltd., Maanshan, China; Registration No. Anhui Medical Device 2017220054) was used. The skin around the insertion point was stabilized with the left hand, while the needle-knife was held in the right hand with the blade parallel to the rat’s longitudinal axis and inserted perpendicularly through the skin. The blade was advanced layer by layer, and the tensioned tissue layer was released with 3-5 cutting actions at the site of tightness. Gentle compression was applied after withdrawal to avoid bleeding. Treatment was administered once every 3 days, for a total of three sessions.
CSR Rat Modeling and Grouping
For efficacy evaluation, thirty-two rats were randomized into 4 groups (n = 8 per group): the Normal (untreated rats), the Sham (rats received the same surgical procedure as the CSR group but without insertion into spinal canal), the CSR (rats with CSR induced by spinal canal insertion), and the CSR + NK (CSR rats received needle-knife therapy) groups.
For pathway validation, forty rats were randomized into 5 groups (n = 8 per group): the Sham, the CSR, the CSR + NK, the CSR + IXA4 (CSR rats received intraperitoneal injection of the IRE1α activator IXA4 [HY-15845, MedChemExpress, Monmouth Junction, NJ, USA] at a dose of 10 mg/kg), 13 and the CSR + NK + IXA4 (CSR rats received intraperitoneal injection of IXA4 followed by needle-knife therapy 20 min later) groups. After completion of needle-knife therapy, behavioral assessments were performed at predetermined time points. Rats were subsequently anesthetized and sacrificed, and intact spinal cord tissues were harvested for storage or immediate experimental analysis as needed.
CSR modeling was performed using the spinal canal insertion method, as previously described by Su et al. 14 All rats were fasted and water-deprived for 12 h prior to surgery. Anesthesia was induced by intraperitoneal injection of sodium pentobarbital (30 mg/kg), and rats were fixed in the prone position. After standard skin preparation and disinfection of the cervical back region, the T2 vertebral level was palpated, and a 3-cm midline incision was made toward the cranial side. Subcutaneous tissues and posterior neck muscles were bluntly dissected to expose the left laminae of C6-T2. Superficial tissues, connective tissue, and ligamentum flavum were cleared from the interlaminar space. A nylon thread (1.5 cm length, ∼0.5 mm diameter) was implanted beneath the nerve root along the spinal axis to apply compression. The wound was closed in layers, followed by routine postoperative analgesia and antibiotics. Model success was confirmed on postoperative day 3 by signs such as curled inward posture of the left forepaw, limping, and non-weight-bearing movement. Behavioral tests, including mechanical withdrawal threshold, thermal nociceptive threshold, and gait score analysis, were performed in CSR rats at designated time points from baseline (prior to needle-knife intervention) through 12 days post-treatment. Upon completion of behavioral testing, rats were euthanized via intraperitoneal administration of an overdose of pentobarbital sodium (150 mg/kg), and spinal cord tissues were collected for subsequent analyses.
Assessment of Mechanical and Thermal Pain Thresholds
Pain-related behaviors, including paw withdrawal mechanical threshold (PWMT) and paw withdrawal thermal latency (PWTL), were evaluated on days 0 (pre-treatment), postoperative days 3, 6, 9, and 12, in a quiet environment by the same operator to minimize variability. PWMT was measured using von Frey filaments (0.4-15 g), applied perpendicularly to the center of the left forepaw plantar surface. A rapid paw withdrawal, shaking, or licking within 8 s or immediately upon removal was considered a positive response. Each test was separated by at least 10 s. The 50% PWMT was calculated using the up-down method as described by Su et al. 15
Thermal sensitivity was measured using the BME-410C automated thermal stimulator (Institute of Biomedical Engineering, Chinese Academy of Medical Sciences, Beijing, China). Rats were placed in a perforated transparent chamber to acclimate for 20-30 min. A focused beam of light was directed onto the left forepaw plantar surface for no longer than 40 s to avoid burns. The latency to paw withdrawal, licking, or escape was recorded as PWTL. Each rat was tested five times with ≥10 min intervals, and the average value was taken as the final thermal threshold. A shorter PWTL indicates higher pain sensitivity and greater nociceptive response.
Gait Scoring
Gait was evaluated on postoperative days 0, 3, 6, 9, and 12. Scoring was performed according to the method of Kawakami et al. 16 to assess gait abnormalities related to paw contracture from pain. Score 1: no paw deformity, normal gait; Score 2: obvious paw deformity (eg, plantar flexion or foot inversion), but normal gait; Score 3: impaired gait with motor paralysis of the left forelimb. A gait score ≥2 indicated significant motor dysfunction and confirmed successful CSR modeling.
Hematoxylin-Eosin (HE) Staining
Spinal cord tissues were fixed in 4% paraformaldehyde for 24-48 h, dehydrated, cleared, and embedded in paraffin. Sections (4 μm thick) were cut in sagittal or transverse planes and baked at 60°C for 2 h. After dewaxing twice in xylene (10 min each), tissue was rehydrated in graded ethanol (100%, 90%, 80%, 70%; 2-5 min each) and rinsed in distilled water. Sections were stained with hematoxylin for 5-10 min, differentiated in 1% acid-alcohol for 1 s, and blued in running water for 10 min. After eosin counterstaining for 5 min, sections were dehydrated again in graded ethanol, cleared in xylene, and mounted with neutral resin. Morphological changes in tissue structure and neurons were examined and photographed under a light microscope.
Double Immunofluorescence Staining
Paraffin-embedded spinal cord sections were permeabilized with 0.3% Triton X-100 for 15 min and blocked with 5% bovine serum albumin (BSA) for 30 min. Sections were incubated overnight at 4°C in the dark with the following primary antibodies: mouse anti-ionized calcium-binding adapter molecule 1 (Iba1, 1:200, Cat# ab283319, Abcam, Cambridge, UK) and rabbit anti-TNF-α (1:200, Cat# ab205587, Abcam). After three washes with phosphate-buffered saline (PBS), sections were incubated at room temperature in the dark for 1 h with secondary antibodies: Alexa Fluor 488-conjugated goat anti-rabbit IgG (green, 1:400) and Alexa Fluor 594-conjugated goat anti-mouse IgG (red, 1:400). Cell nuclei were counterstained with 4′,6-diamidino-2-phenylindole (DAPI, blue) for 5 min. After mounting with anti-fade medium, sections were observed and imaged under a fluorescence microscope.
Enzyme-Linked Immunosorbent Assay (ELISA)
ELISA was performed to measure the levels of IL-6 and TNF-α in spinal cord tissue. Tissues from each group were rinsed with pre-chilled PBS to remove residual blood, blotted dry, and weighed. Pre-chilled RIPA lysis buffer (containing 1% protease inhibitor; Beyotime Biotechnology, Shanghai, China) was added at a ratio of 1:10 (g:mL) based on tissue weight. Homogenization was carried out on ice, followed by centrifugation at 12 000×g for 15 min at 4°C. The supernatants were collected for analysis. The concentrations of IL-6 and TNF-α were determined according to the instructions of commercial ELISA kits (IL-6, Cat# PI328; TNF-α, Cat# PT516; both from Beyotime Biotechnology).
Nissl Staining
Paraffin-embedded spinal cord sections were dewaxed in xylene three times (5-10 min each), followed by rehydration in 100% ethanol (5 min), 90% ethanol (2 min), 70% ethanol (2 min), and distilled water (2 min). Nissl staining was performed according to the manufacturer’s instructions (Nissl Staining Solution, Cat# C0117, Beyotime Biotechnology). Briefly, the sections were stained with Nissl solution for 10 min, rinsed in distilled water twice, followed by 95% ethanol for 5 s, and 70% ethanol twice. After final dehydration in 95% ethanol (2 min × 2) and clearing in xylene (5 min × 2), slides were mounted with neutral resin. Neurons, exhibiting characteristic blue-purple granular staining, were visualized and photographed under a microscope. Neuronal counts were performed in 3 randomly selected fields per section for statistical analysis.
Quantitative Real-Time Polymerase Chain Reaction (qRT-PCR)
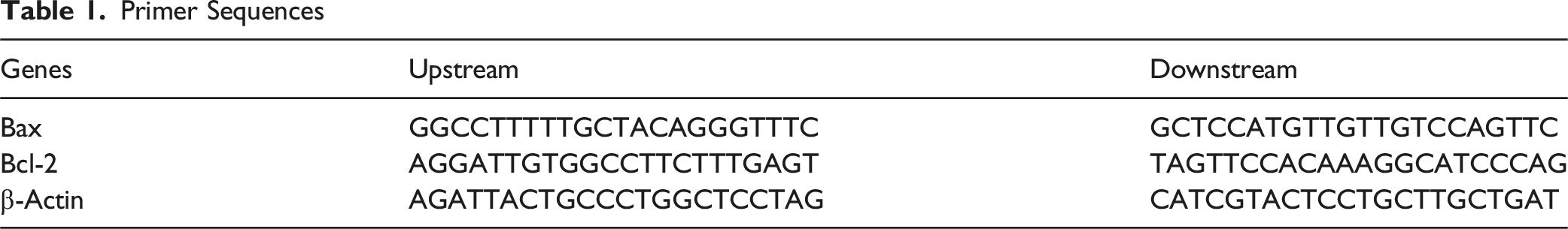
Primer Sequences
NeuN and TUNEL Double Staining
Neuronal apoptosis was assessed by co-staining with NeuN and TUNEL, using the TUNEL Apoptosis Detection Kit (Cat# C1088, Beyotime Biotechnology). After dewaxing in xylene and dehydration through a graded ethanol series, sections were rinsed in distilled water and subjected to antigen retrieval with 20 μg/mL DNase-free proteinase K (Cat# ST533, Beyotime Biotechnology). Endogenous peroxidase activity was blocked using methanol containing 3% hydrogen peroxide. After three washes with PBS, sections were incubated with rabbit anti-NeuN (1:200, Cat# ab177487, Abcam) followed by Alexa Fluor-conjugated goat anti-rabbit IgG (1:200, Cat# ab150080, Abcam). TUNEL detection solution was then added and incubated in the dark at 37°C for 1 h. Cell nuclei were counterstained with DAPI (Beyotime Biotechnology) and sealed with antifade mounting medium. Apoptotic neurons (TUNEL+/NeuN+) were visualized and photographed using a fluorescence microscope.
Western Blotting (WB)
Total protein was extracted from spinal cord tissue using RIPA lysis buffer containing protease inhibitors (Beyotime Biotechnology). Homogenates were incubated on ice for 30 min and centrifuged at 12 000×g for 15 min at 4°C. Supernatants were collected, and protein concentration was determined using a bicinchoninic acid assay kit (Beyotime Biotechnology). Equal amounts (30 μg) of protein were separated via SDS–polyacrylamide gel electrophoresis and transferred onto polyvinylidene difluoride membranes. After blocking in non-fat milk for 1-2 h, membranes were incubated overnight at 4°C with the following primary antibodies targeting Bax (1:1000, Cat# A12009, ABclonal, Wuhan, China), Bcl-2 (1:1000, Cat# A19693, ABclonal), Phosphorylated IRE1α (p-IRE1α, 1:1000, Cat# PA5-105424, Cell Signaling Technology, Danvers, MA, USA), XBP1-spliced form (XBP1s, 1:1000, Cat# 24868-1-AP, Proteintech Group, Rosemont, IL, USA), and β-actin (1:1000, Cat# AC026, ABclonal). After washing, membranes were incubated for 1 h at room temperature with horseradish peroxidase-conjugated goat anti-rabbit IgG (H + L) secondary antibody (1:5000, Cat# AS014, ABclonal). Protein bands were visualized using enhanced chemiluminescence reagents and imaged using a gel documentation system. Band intensities were quantified using ImageJ software (NIH, Bethesda, MD, USA), and the expression levels of target proteins were normalized to β-actin.
Statistical Analysis
Data are expressed as mean ± standard deviation (SD). GraphPad Prism 9 software (GraphPad Software, San Diego, CA, USA) was used for statistical analysis and visualization. One-way analysis of variance (ANOVA) was applied to compare differences among multiple groups. A P-value <0.05 was considered statistically significant.
Results
Needle-Knife Therapy Restores motor Function and Alleviates Pain in CSR rats
Clinically, needle-knife therapy has been shown to relieve pain, improve cervical range of motion and function, and regulate inflammatory markers in patients with CSR, demonstrating notable therapeutic efficacy.
17
To further investigate its effects and potential molecular mechanisms, we established a CSR rat model using the spinal canal insertion method and applied needle-knife therapy. No significant differences in PWMT, PWTL, or gait scores were observed between the Normal and Sham groups (P > 0.05). Compared to the Sham group, CSR rats exhibited significantly reduced PWMT and PWTL, along with increased gait disturbance scores. Following needle-knife therapy, CSR rats showed significantly elevated pain thresholds (PWMT and PWTL) and reduced gait scores (Figure 1A–C). These findings indicate that needle-knife therapy markedly restores motor function and alleviates pain in CSR rats. Improvement of motor function and pain relief by needle-knife therapy in CSR rats. (A) Mechanical pain threshold (PWMT), (B) thermal pain threshold (PWTL), and (C) gait score in each group. ****P < 0.0001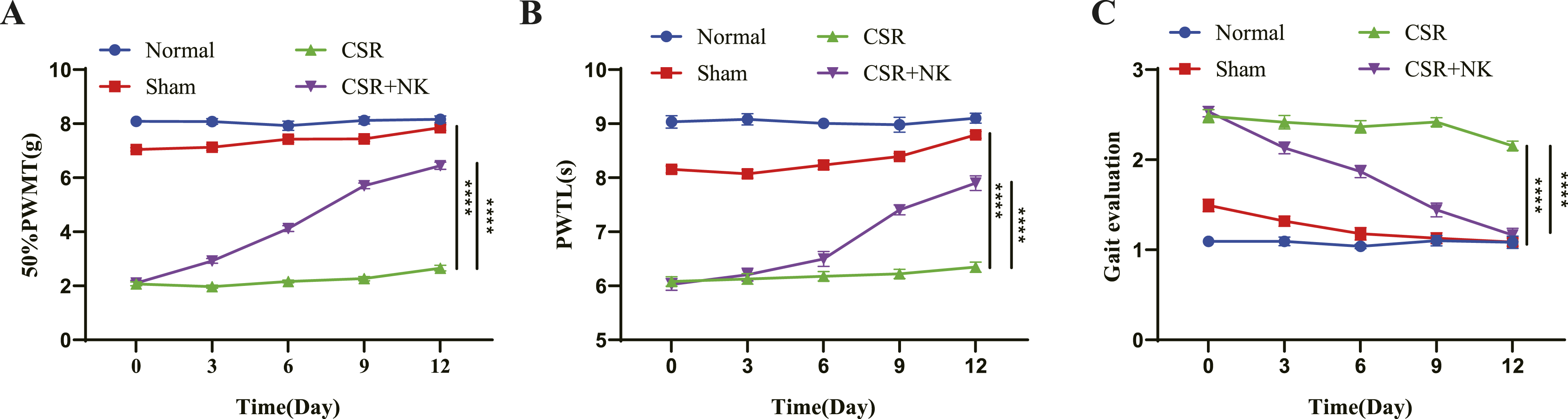
Needle-Knife Therapy Alleviates Spinal Cord Pathology and Suppresses Inflammation in CSR Rats
HE staining and ELISA were used to assess spinal cord histopathology and inflammatory cytokine levels. HE staining revealed that the spinal cord structures in the Normal and Sham groups were intact, with no significant pathological damage or inflammatory cell infiltration. In contrast, spinal cords from the CSR group exhibited loose architecture, edema, and mild inflammatory infiltration, indicating significant local tissue damage and inflammation due to nerve root compression. Needle-knife therapy alleviated these structural damages and reduced inflammatory cell infiltration (Figure 2A). ELISA showed that levels of IL-6 and TNF-α in the Sham group were slightly elevated but not significantly different. However, both IL-6 and TNF-α levels were markedly increased in the CSR group and significantly reduced after needle-knife therapy (Figure 2B and C). Immunofluorescence staining revealed increased numbers of Iba1-positive microglia and elevated TNF-α expression in the CSR group, with evident co-localization, indicating a proinflammatory microglial phenotype. Needle-knife therapy markedly reduced the number of Iba1+/TNF-α+ double-positive cells, suggesting that it inhibits microglial proinflammatory polarization (Figure 2D). Collectively, these results demonstrate that needle-knife therapy exerts anti-inflammatory and neuroprotective effects by alleviating spinal cord pathology in CSR. Attenuation of spinal cord pathological damage and inflammatory cytokine expression by needle-knife therapy in CSR rats. (A) HE staining showing pathological changes in spinal cord tissue across groups (scale bar = 100 μm). (B, C) ELISA results of IL-6 and TNF-α levels in spinal cord tissue. (D) Immunofluorescence double staining of Iba1 and TNF-α showing microglial activation (red: Iba1; green: TNF-α; blue: DAPI; scale bar = 200 μm). ****P < 0.0001
Needle-Knife Therapy Attenuates neuronal Apoptosis and Protects Neurons in CSR Rats
To investigate the effects of needle-knife therapy on neuronal preservation, Nissl staining was performed. Spinal cords from the Normal and Sham groups showed abundant neurons with evenly distributed Nissl bodies and intact morphology. In the CSR group, neuronal count was significantly reduced, morphology was shrunken, and Nissl body staining was sparse. Rats in the CSR + NK group displayed improved neuronal morphology and increased Nissl staining intensity, indicating neuroprotective effects (Figure 3A). To assess neuronal apoptosis, TUNEL/NeuN double immunofluorescence staining was conducted. Few TUNEL-positive cells were observed in the Normal and Sham groups, and NeuN-positive neurons exhibited normal morphology. In the CSR group, TUNEL-positive apoptotic cells were significantly increased in the anterior horn of the spinal cord, co-localizing with NeuN, indicating elevated neuronal apoptosis. Needle-knife therapy significantly reduced TUNEL+/NeuN+ double-positive cell numbers (Figure 3B and C). qRT-PCR and WB analysis further confirmed that pro-apoptotic protein Bax was significantly upregulated and anti-apoptotic Bcl-2 was downregulated in CSR rats. Needle-knife therapy reversed these changes, decreasing Bax and increasing Bcl-2 expression (Figure 3D–F). These data suggest that needle-knife therapy protects neurons in CSR by inhibiting apoptosis. Reduction of neuronal damage and inhibition of neuronal apoptosis by needle-knife therapy in CSR rats. (A) Nissl staining assessing neuronal number and morphology in spinal cord tissue (scale bar = 200 μm). (B/C) TUNEL and NeuN dual immunofluorescence staining showing neuronal apoptosis (red: TUNEL-positive cells; green: NeuN-positive cells; blue: DAPI), scale bar = 200 μm. (D-F) qRT-PCR (D/E) and WB (F) results of Bax and Bcl-2 expression at mRNA and protein levels. ****P < 0.0001
Needle-Knife Therapy Inhibits Hyperactivation of IRE1α-XBP1 Signaling in CSR Rats
Previous studies have shown elevated levels of XBP1s and p-IRE1α in the diseased tissues of Alzheimer’s disease patients.18,19 The IRE1α-XBP1 signaling, a key branch of endoplasmic reticulum stress, has been implicated in neuronal apoptosis.12,20 To explore its relevance in CSR, we evaluated levels of p-IRE1α and XBP1s in spinal cord tissues. WB results showed significantly elevated levels of both p-IRE1α and XBP1s in the CSR group, whereas needle-knife therapy significantly reduced the expression of both proteins (Figure 4). These findings indicate aberrant activation of the IRE1α-XBP1 axis in CSR, which is suppressed by needle-knife therapy. Downregulation of key proteins in the IRE1α-XBP1 signaling in the spinal cord of CSR rats by needle-knife therapy. WB analysis of p-IRE1α and XBP1s protein levels across groups. ****P < 0.0001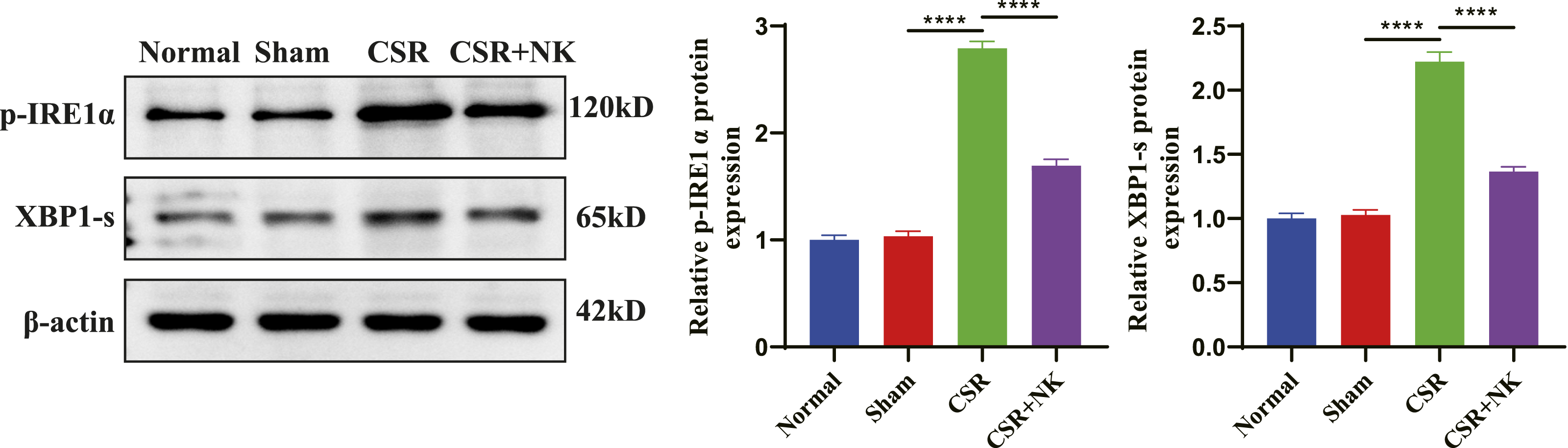
Needle-Knife Therapy Alleviates Motor Dysfunction and Pain in CSR Rats by Suppressing the IRE1α-XBP1 Signaling
To determine whether needle-knife therapy exerts therapeutic effects via the IRE1α-XBP1 signaling, we activated the pathway in CSR rats using the specific IRE1α activator IXA4 (10 mg/kg, intraperitoneally) and evaluated motor function. WB analysis confirmed that IXA4 treatment increased p-IRE1α and XBP1s levels, partially counteracting the inhibitory effects of needle-knife therapy on this pathway (Figure 5A). Functionally, IXA4 treatment further reduced PWMT and PWTL and increased gait disturbance scores in CSR rats. Furthermore, for rats in the CSR + NK + IXA4 group, the beneficial effects of needle-knife therapy were significantly diminished (Figure 5B–D). These findings suggest that activation of the IRE1α-XBP1 pathway partially reverses the functional benefits of needle-knife therapy, supporting the role of this signaling in mediating the therapeutic effects of needle-knife therapy in CSR. Alleviation of motor dysfunction and pain through inhibition of the IRE1α-XBP1 signaling pathway by needle-knife therapy in CSR rats. (A) WB verification of the effect of IXA4 on p-IRE1α and XBP1-s levels in spinal cord tissue. (B–D) Mechanical pain threshold (PWMT), (C) thermal pain threshold (PWTL), and (D) gait score following IXA4 administration in CSR rats with or without needle-knife therapy. ****P < 0.0001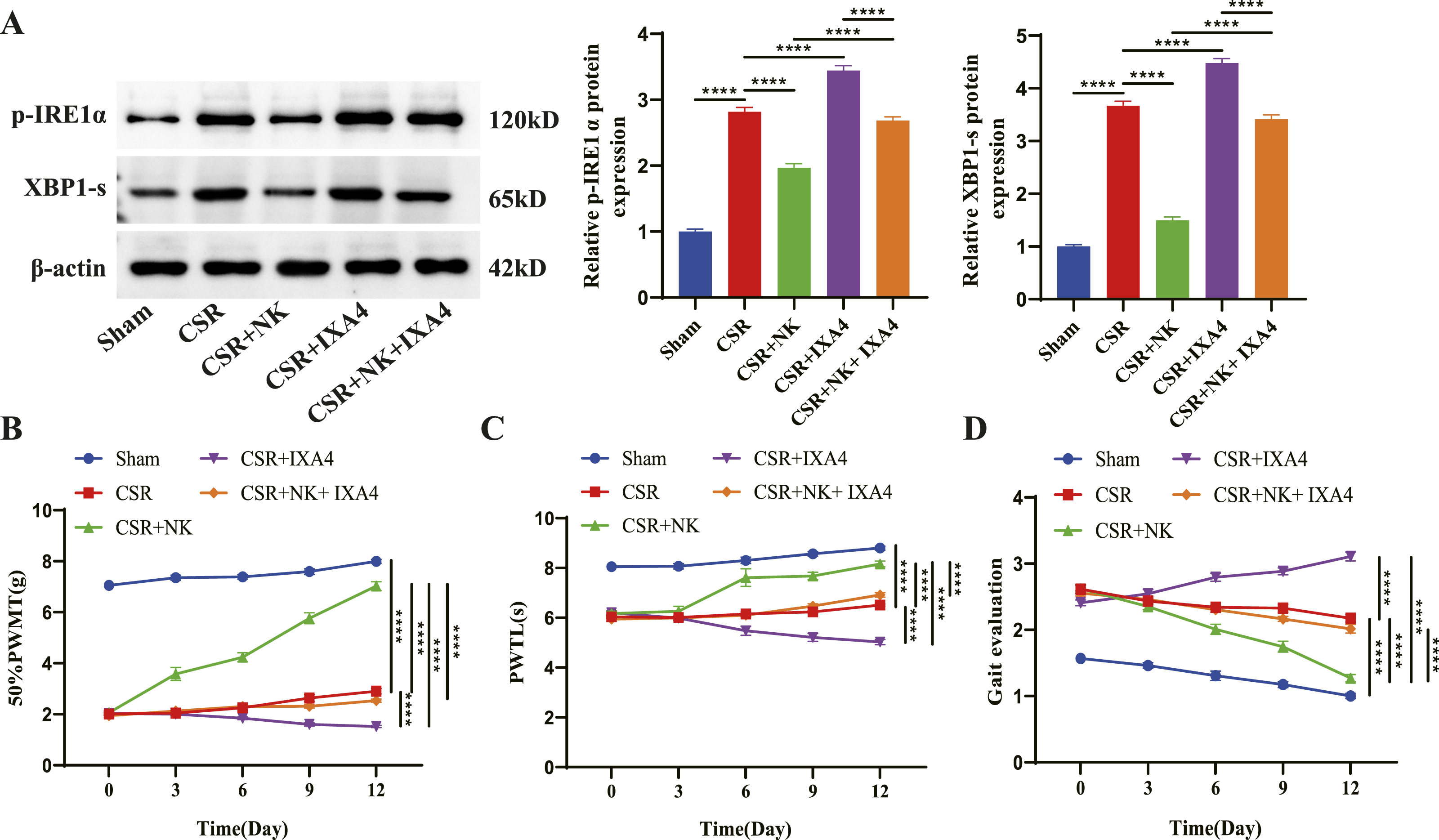
Needle-Knife Therapy Alleviates Pathological Damage, Inflammation, and neuronal Apoptosis in CSR rats via Suppression of the IRE1α-XBP1 Signaling
We further explored whether the protective effects of needle-knife therapy on spinal pathology, inflammation, and neuronal apoptosis are mediated by inhibition of the IRE1α-XBP1 pathway. HE staining, ELISA, and immunofluorescence showed that rats in the CSR + IXA4 group had more severe spinal cord damage, including disorganized architecture, aggravated edema, and increased inflammatory cell infiltration. In the CSR + NK + IXA4 group, spinal cord morphology was more compromised and inflammatory cell infiltration was higher than in the CSR + NK group (Figure 6A). ELISA revealed significantly higher levels of IL-6 and TNF-α in the CSR + IXA4 group compared to the CSR group, and similarly, elevated levels of IL-6 and TNF-α were found in the CSR + NK + IXA4 group compared to that in the CSR + NK group (Figure 6B and C), indicating that pathway activation intensifies inflammation and disrupts the anti-inflammatory effects of needle-knife therapy. Increased Iba1-positive microglia and TNF-α expression were observed in both CSR + IXA4 and CSR + NK + IXA4 groups, with strong cytoplasmic co-localization and elevated Iba1+/TNF-α+ double-positive cell counts (Figure 6D), further supporting the role of IRE1α-XBP1 signaling in microglial activation. Inhibition of pathological damage, inflammation by needle-knife therapy via suppression of the IRE1α-XBP1 signaling. (A) HE staining evaluating pathological changes in spinal cord tissue after IXA4 treatment (scale bar = 200 μm). (B, C) ELISA analysis of IL-6 and TNF-α levels following IXA4 intervention. (D) Immunofluorescence detection of microglial activation following IXA4 treatment. *P < 0.05, **P < 0.01, ***P < 0.001, ****P < 0.0001
Neuronal assessments also demonstrated that IXA4 significantly reduced neuronal count, altered neuronal morphology (Figure 7A and B), increased TUNEL+/NeuN+ apoptotic neuron numbers (Figure 7C and C), upregulated Bax, and downregulated Bcl-2 expression in rats in the CSR + NK group (Figure 7E and F). These findings suggest that IRE1α-XBP1 activation compromises the neuroprotective, anti-apoptotic effects of needle-knife therapy in CSR. Inhibition of neuronal apoptosis by needle-knife therapy via suppression of the IRE1α-XBP1 signaling. (A/B) Nissl staining of spinal cord neurons after needle-knife therapy with or without IXA4, scale bar = 200 μm. (C/D) TUNEL/NeuN dual immunofluorescence staining assessing neuronal apoptosis after IXA4 treatment, scale bar = 200 μm (red: TUNEL-positive cells; green: NeuN-positive cells; blue: DAPI). (E/F) qRT-PCR and WB detection of Bax and Bcl-2 expression in spinal cord tissue. *P < 0.05, **P < 0.01, ****P < 0.0001
Discussion
Previous clinical and experimental studies have demonstrated that acupuncture can alleviate pain, improve nerve function, and reduce inflammation in CSR by modulating neural signaling and promoting local blood circulation. 21 Despite its widespread application and favorable safety profile, conventional acupuncture often lacks precision in targeting deep soft tissue adhesions and nerve root entrapment, which limits its therapeutic efficacy in cases involving more severe structural compression. 22 Needle-knife therapy was developed to overcome these limitations by integrating the minimally invasive advantages of acupuncture with the mechanical release capabilities of modern surgical techniques. 23 In the present study, we established a rat model of CSR and found that needle-knife therapy confers neuroprotective effects in CSR by targeting both inflammation and apoptosis.
In this study, behavioral assessments including PWMT, PWTL, and gait score demonstrated that needle-knife therapy significantly alleviated neuropathic pain and improved motor function in rats with CSR. These improvements are consistent with clinical observations where needle-knife treatment has been reported to relieve pain and enhance mobility in patients with cervical spondylosis-related conditions. 24 The therapeutic effects are likely attributed to the mechanical release of soft tissue adhesions and restoration of the nerve root canal space. In CSR, fibrous tissue hyperplasia and myofascial adhesions surrounding the cervical vertebrae may contribute to nerve root entrapment and chronic pain. 25 Needle-knife therapy, which involves minimally invasive subcutaneous release of contractured fascia or tendinous attachments, likely relieves localized mechanical compression and restores microcirculation around the nerve root, thereby mitigating nociceptive signaling. 26 Our histological analysis revealed substantial structural disruption and inflammatory infiltration in the spinal cord of CSR rats, which were significantly attenuated following needle-knife intervention. Meanwhile, elevated levels of proinflammatory cytokines IL-6 and TNF-α were found in CSR rats, both of which were markedly reduced after treatment. IL-6 and TNF-α are key mediators of chronic neuroinflammation and neuropathic pain. They promote central sensitization, glial activation, and neuronal hyperexcitability, and have been implicated in both the development and persistence of radicular pain. 27 Needle-knife therapy appears to modulate this inflammatory cascade, potentially through decompression-induced restoration of neuronal homeostasis and downregulation of cytokine signaling. This is consistent with studies showing that relieving mechanical stress on neural tissue can significantly suppress neuroinflammation and glial activation. 28 Additionally, microglia are primary immune effector cells in the central nervous system and play a pivotal role in the development and maintenance of neuropathic pain by releasing cytokines, chemokines, and reactive oxygen species. 29 Activated microglia in the dorsal and ventral horns can sensitize spinal neurons, exacerbate apoptosis, and contribute to neurodegenerative changes. 30 Following needle-knife therapy, we observed a reduction in Iba1+/TNF-α+ cell populations, indicating suppression of microglial proinflammatory polarization. This suggests that the treatment may disrupt the pain-sustaining loop between mechanical injury and microglial activation. Furthermore, in our study, CSR rats exhibited significant neuronal loss and increased apoptosis in the spinal cord, which were substantially alleviated by needle-knife therapy. The balance between pro-apoptotic Bax and anti-apoptotic Bcl-2 proteins is central to neuronal survival in injury conditions. 31 Our data showed that needle-knife therapy downregulated Bax and upregulated Bcl-2 at both the protein and mRNA levels, suggesting effective inhibition of the apoptotic pathway.
Mechanically, IRE1α-XBP1 signaling is a major branch of the ER stress response, which is activated in response to the accumulation of misfolded or unfolded proteins. Upon activation, IRE1α undergoes autophosphorylation and splices XBP1 mRNA, producing the active transcription factor XBP1s, which regulates the expression of genes involved in protein folding, ER-associated degradation, and apoptosis. 32 While initially protective, prolonged activation of the IRE1α-XBP1 pathway is implicated in promoting apoptosis, inflammation, and cellular dysfunction in various neurodegenerative and inflammatory central nervous system disorders. 33 In our study, CSR rats exhibited marked activation of this signaling axis, as evidenced by upregulated p-IRE1α and XBP1s levels in the cervical spinal cord. Needle-knife therapy significantly downregulated the expression of both p-IRE1α and XBP1s, suggesting that attenuation of ER stress and its downstream proapoptotic consequences contributes to the therapeutic effects. Moreover, to directly assess the causative role of the IRE1α-XBP1 signaling in CSR and the mechanism underlying the therapeutic effects of needle-knife therapy, we employed IXA4, a small-molecule activator of IRE1α known to enhance its endoribonuclease activity and promote XBP1s. IXA4 has been validated as a tool compound to selectively stimulate the IRE1α-XBP1 pathway. 34 In our study, administration of IXA4 to CSR rats receiving needle-knife therapy effectively reversed the therapeutic improvements conferred by the intervention. Given the established role of ER stress in neuronal apoptosis and neuroinflammation,35,36 suppression of the IRE1α-XBP1 pathway may serve as a critical molecular mechanism by which needle-knife therapy alleviates spinal cord pathology and improves functional outcomes in CSR. Furthermore, needle-knife therapy is traditionally considered to exert therapeutic effects by releasing soft tissue adhesions and improving local microcirculation. 8 Based on our findings, it is reasonable to hypothesize that these mechanical and hemodynamic improvements may contribute to the observed attenuation of ER stress. Specifically, enhanced local blood flow may alleviate ischemia and hypoxia in compressed nerve tissues, thereby reducing the accumulation of misfolded proteins and the activation of ER stress sensors such as IRE1α. Given that hypoxia and metabolic disturbance are well-recognized triggers of ER stress and unfolded protein response signaling, 37 improved tissue perfusion may indirectly suppress the IRE1α-XBP1 pathway. Additionally, better clearance of inflammatory mediators and metabolic byproducts could further mitigate cellular stress and neuronal apoptosis. Although this proposed mechanism requires direct experimental validation, it provides a plausible biological link between the mechanical action of needle-knife therapy and its molecular effects observed in this study.
While the present study provides compelling evidence supporting the therapeutic efficacy and mechanistic underpinnings of needle-knife therapy in CSR, limitation remains. For instance, the study was conducted in a rodent model of CSR, which, although widely used for simulating nerve root compression and its associated neuroinflammatory and neuropathic responses, may not fully recapitulate the chronic and multifactorial nature of human cervical spondylotic radiculopathy. In clinical settings, CSR typically develops through progressive degenerative changes, including intervertebral disc degeneration, osteophyte formation, ligament hypertrophy, and sustained nerve root compression. These processes are often accompanied by long-term inflammation, ischemia, and structural remodeling. In contrast, the present model primarily reflects an acute compressive insult with relatively rapid onset. Therefore, while this model is valuable for elucidating mechanistic pathways, such as the involvement of IRE1α-XBP1 signaling in neuroinflammation and apoptosis, caution is warranted when extrapolating these findings directly to human disease. Future studies employing chronic compression models or large-animal systems may better capture the progressive pathology of CSR and enhance translational relevance.
Conclusion
This study demonstrates that needle-knife therapy effectively alleviates mechanical and thermal hyperalgesia and improves motor dysfunction in rats with CSR. The neuroprotective effects are achieved, at least in part, by downregulating the IRE1α-XBP1 signaling, thereby inhibiting proinflammatory microglial activation and neuronal apoptosis. Administration of the IRE1α agonist IXA4 partially reversed the therapeutic benefits of needle-knife therapy, further confirming the pivotal role of this pathway in mediating its effects. To our knowledge, this is the first study to elucidate the molecular mechanism of needle-knife therapy from the perspective of ER stress signaling, providing both theoretical rationale and experimental evidence for its clinical application in CSR.
Footnotes
Ethical Considerations
All animal procedures were approved by the Ethics Committee of Hunan University of Chinese Medicine (Ethics Number: HNUCM21-2512-17) and were conducted in accordance with institutional guidelines.
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This study was supported by 2025 Hunan University of Chinese Medicine University-Hospital Joint Fund (2025XYLH146).
Declaration of Conflicting Interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.