
Abstract
This study aimed to demonstrate the histopathology and immunoexpression of exercise-derived myokines in dentate gyrus (DG), medial prefrontal cortex (mPFC) and cerebellum of depressed Wistar rats during depression and after practising voluntary running. Depression was developed by forced swimming for 2 weeks. Voluntary running was performed by voluntary running for 3 weeks. Brain sections were processed and immunostained to detect brain-derived neurotrophic factor (BDNF), macrophage migration inhibitory factor (MIF), vascular endothelial growth factor (VEGF) and interleukin-6 (IL-6). ImageJ software was used to measure the optical density (OD). BDNF was expressed in neurons in DG, mPFC and granular and Purkinje cells in cerebellum. MIF was expressed in neurons of sub-granular zone in DG, mPFC and Purkinje cells. VEGF was expressed in many neurons in DG, mPFC and Purkinje cells. IL-6 was expressed in some neurons in DG, in neuropil of mPFC and in Purkinje cells. In depression, the OD of studied myokines significantly decreased in all examined areas. After voluntary running, the OD of myokines significantly increased in all areas. This study defines the immunohistochemical expression of myokines in brain areas in depression and after voluntary running and reveals the involvement of the mPFC and cerebellum in the pathophysiology of depression.
Introduction
The beneficial effects of physical exercise on the indicators of depression have been previously shown.1,2 Variable levels of cytokines and growth factors are released in response to exercise and are termed myokines. Conflicting outcomes have been reported by studies investigating the correlation between these exercise-derived factors and depressive behaviours in rodents.3–6
In patients with major depression, brain-derived neurotrophic factor (BDNF) is positively related to the volume and function of the hippocampus, 7 and BDNF deficiency is associated with significant neurological defects in the frontal cortex. 8 Macrophage migration inhibitory factor (MIF) is thought to play a role in the neuro-immune interaction, which may be linked to depression. 9 The levels of vascular endothelial growth factor (VEGF) have been recently shown to be related to neurogenesis in depression cases. 10 In addition, interleukin-6 (IL-6) has recently been found to play a significant role in mediating the psychological and neurobiological manifestations of depression. 11
The dentate gyrus (DG) in the hippocampal formation has been implicated in cognitive and mood disorders due to the reduction in adult hippocampal neurogenesis. 12 The medial prefrontal cortex (mPFC) has connections with hippocampal structures associated with long-term memory and central autonomic structures associated with the expression of emotions.13,14 The cerebellum has strong connections with brain regions responsible for emotions and is, thus, believed to play an important role in the pathophysiology of depression.15,16 Recent studies investigating the effect of physical exercise on the morphological changes in the cerebellum 17 and whether physical exercise promotes angiogenesis and the functional recovery of cells after brain injuries have yielded inconsistent results.18,19
While the levels of myokines could be linked to depression and influence the plasticity and survival of neurons, 20 no studies have used immunohistochemistry to investigate the expression of these myokines in brain areas vulnerable to depression. This study aims to investigate the following: (a) the expression of BDNF, MIF, VEGF and IL-6 in the DG, mPFC and cerebellum using a rat model of depression and (b) the effect of voluntary running on the expression of these myokines in these areas of the brain.
Materials and methods
This experimental case–control study was conducted at King Fahd Centre for Medical Research, King Abdul-Aziz University, Jeddah, Saudi Arabia, and the research laboratory of the College of Medicine, Taibah University, Almadinah, Saudi Arabia.
All animal handling and experimental procedures were conducted according to the EC Directive 86/609/EEC for animal experiments (http://ec.europa.eu/environment/chemicals/lab_animals/legislation_en.htm) and the guidelines of the National Committee of Bio-Ethics in Saudi Arabia (https://doi.org/10.1111/dewb.12114). Ethical appro-val was obtained from the ethics committee of King Abdul-Aziz University, Jeddah, Saudi Arabia.
Experimental animals
A total of 60 male Wistar rats aged 8–10 weeks weighing 200–250 g were obtained from the Experimental Animal Centre of the King Fahd Centre for Medical Research, Jeddah, Saudi Arabia. The rats were housed for 7 days to allow acclimatisation to the laboratory conditions and were observed for any abnormal behaviour in their ordinary movement and feeding behaviour. Only those rats that showed apparently normal movements and normal feeding behaviour were included in the study.
Before the experiment, all rats were exposed to a 6-min forced swimming test (FST), and only those animals that showed extreme behaviour during the test were selected for the experiment. 21 The selected animals (45 rats) were singly housed in well-ventilated cages with ad libitum access to food and water. The environment in the skill laboratory was controlled at a 12 h/12 h light/dark cycle (the lights were turned on at 7:00 a.m.), temperature of 22°C–24°C and humidity of 50%–60%. 22
The 45 selected rats were randomly divided into the following three groups each comprising 15 rats:
Control group. These rats were maintained on a standard chow diet for 5 weeks, including the weekends (35 days), screened using the 6-min FST to automatically record their locomotor activity and immediately sacrificed.
Depression group. These rats were subjected to the forced swimming (FS) protocol for 2 weeks, including the weekends (14 days), housed for an additional 3 weeks without exercise, screened using the 6-min FST to record their locomotor activity and immediately sacrificed.
Depression/exercise group. These rats were subjected to the FS protocol for 2 weeks, including the weekends (14 days), allowed to perform voluntary wheel running for 3 weeks, screened using the 6-min FST to record their locomotor activity and immediately sacrificed.
Development of the animal model of depression
This model was developed using a protocol described by Eldomiaty et al. 23 The rats were individually subjected to FS inside a vertical glass cylinder (height: 60 cm; diameter: 22 cm) containing water (height: 45 cm) maintained at 23°C–25°C with no possibility of escape. After 15 min of swimming, the rats were removed and allowed to dry for 15 min in a heated enclosure (32°C) before being returned to their home cages. This procedure was repeated once daily for 2 weeks, including the weekends (14 times). This procedure was a modification of other protocols that also used FS to develop animal models of depression.24–26
Voluntary wheel running
After developing depression (2 weeks of FS), the rats in the depression/exercise group were moved to cages equipped with rat running wheels prepared for voluntary running and activity wheel counters (Lafayette Instrument Company, Inc., North Lafayette, IN, USA) (Figure 1). The rats were allowed to run voluntarily for 3 weeks (21 days). The wheels were inspected every other day to ensure that they were working properly. 27

Voluntary rat running wheel (Lafayette Instrument Company, Inc., North Lafayette, IN, USA). The figure shows a rat while accessing the wheel voluntarily.
Assessment of locomotor parameters in rats from different groups during the FST
To confirm the development of the depressive state after the FS and/or its relief after voluntary running, we performed the 6-min FST with automatic tracing using computer software (Ethovision XT version 8.0; Noldus Information Technology, Wageningen, The Netherlands) to determine the distance moved by the rat in centimetres and the immobility duration (time spent passively floating) in seconds.24,28–30 All rats were subjected to this test immediately before sacrifice between 8:00 and 10:00 a.m. 31
Light microscopy structural study of different brain regions
At the specified time of sacrifice, the brain was freshly dissected, and half of the brain was coronally sectioned and fixed in buffered formalin for the histological examination of the DG, mPFC and cerebellum. 32 The brain tissue sections were stained with haematoxylin and eosin (H&E) and examined under an Olympus BX 36 Bright field Automated microscope. The images were digitised in a 2448 × 1920-pixel matrix using a DP27 colour digital video camera to determine the structural changes in different areas.
Immunohistochemical staining and evaluation of immunoexpression
Serial paraffin sections were stained using primary polyclonal anti-rat antibodies against selected myokines (Uscn Life Science Business Co., Ltd, Wuhan; polyclonal Rat BDNF PAA011Ra01, Rat MIF PAA698Ra01, Rat VEGF PAA143Ra71, Rat IL-6 PAA079Ra71). The staining procedure was performed using an immunoperoxidase technique with a Bench-Mark instrument (recommended by Ventana Company, Oro Valley, AZ, USA). The paraffin sections were deparaffinised, rehydrated, treated with 3% H2O2 for 5 min and washed with phosphate-buffered saline (PBS) for 15 min. The sections were blocked with 1.5% normal goat serum in PBS and then incubated for 45 min at room temperature with the primary antibody. The sections were subsequently incubated with a second-stage biotinylated antibody at room temperature. After rinsing with PBS, the reaction products were visualised by immersing the sections in diaminobenzidine. Finally, the sections were counterstained with haematoxylin, dehydrated and cover-slipped using Protex mounting media (DAB-Stock Stain box; Boster Biotechnology, Pleasanton, CA, USA). 33
A light microscopy examination of the immunoexpression of different myokines was performed in different brain areas, and the immunohistochemical staining was evaluated by measuring the optical densities (ODs) of the stained sections. 34 Briefly, five representative non-overlapping fields were captured as photomicrographs at a magnification of ×200 (microscopic field area of 786.4 u2). The captured images were transferred to a computer for an image analysis. The OD evaluation was performed using the ImageJ (NIH) software, version 2.0.0-beta4 (National Institutes of Health, Bethesda, MD, USA). Each image was first deconvoluted using DAB vector into three different coloured images (i.e. green, brown, and blue). The calibration of the brown DAB image was performed by measuring the mean integrated intensities (mean grey value) of five non-overlapping areas of stained tissue. The intensity numbers were converted into the OD using the following equation: (OD = log (max intensity/mean intensity)), where the max intensity = 250 and the mean intensity = mean grey value. The results were exported to a Microsoft Excel spreadsheet.
Statistical analysis
The statistical analysis was performed using the IBM SPSS (version 21) statistical package. All data are expressed as the means ± standard errors of the means (SEM). Comparisons between groups were performed using one-way analysis of variance (ANOVA), followed by Fisher’s multiple comparison tests as necessary. The statistical significance of the mean comparisons was set at P < 0.05. The correlation between different myokines in all groups was done using Pearson correlation test and the P and r values were recorded. The statistical analyses performed are described in the table’s captions.
Results
FST before sacrifice
The results of the automatic scoring of the locomotor parameters during the FST in the rats from the different groups are shown in Figure 2.

Graph illustrating the immobility duration and the distance moved for control, depression and depression/exercise groups (15 rats each) during the 6-min forced swimming test (means ± SEM).
The mean distance moved by the rats in the depression group (310.7 ± 33.7 cm) was significantly lower than that in the control rats (693.6 ± 37.5 cm) (P < 0.001). In addition, the mean distance moved by the rats in the depression/exercise group (606.1 ± 33.5 cm) was significantly higher than that in the rats in the depression group (P < 0.001).
The mean immobility duration in the depression group (288 ± 8.07 s) was significantly higher than that in the control rats (160.05 ± 12.7 s) (P < 0.001). The mean immobility duration in the rats in the depression/exercise group (192.14 ± 17.91 s) was significantly lower than that in the rats in the depression group (P < 0.001).
Histopathological study
DG
According to the examination of the H&E stained sections, the DG in the control rats consisted of the following components: upper limb, lower limb and stem (Figure 3(a)). Each component comprised the following layers: (a) the molecular stratum, which comprised neuronal processes, glial cells and a few neurons; (b) the stratum granulosum, which comprised densely packed granular cells with rounded vesicular nuclei and several immature granule cells with oval nuclei that appeared in the sub-granular zone (SGZ) and between the mature neurons; and (c) stratum pleomorph, which comprised glial cells and several neurons (Figure 3(b) and (c)).

A photomicrograph of sections in dentate gyrus of adult rat. (a) Specimen of control rat shows the dentate gyrus is formed of upper limb (UL), lower limb (LL) and stem (S). (b, c) The upper and lower limbs of control rat consist of three strata. Stratum molecular (SM) contains neuronal processes (white arrows), glial cells (arrow heads) and few neurons (dotted arrow). Stratum granulosum (SG) contains granule cells with rounded vesicular nuclei (thin black arrows). Immature small granular cells with oval nuclei are seen in the sub-granular zone and in between the mature neurons (thick arrows). Stratum pleomorph (SP) contains glial cells (arrow heads) and some neurons (dotted arrows). (b′, c′). The upper and lower limbs of depressed rat show many cells with dark-stained pyknotic nuclei (thin black arrows). Small immature neurons (thick arrows) and apparent vacuolations (white arrows) are present in the sub-granular zone of the lower limb. (b″, c″) The upper and lower limbs of depression/exercise rat show numerous granule cells with vesicular nuclei (thin black arrows). Immature small granular cells (thick arrows) and little vacuolations (white arrows) are seen in the sub-granular zone. Many dilated blood vessels appeared in the pleomorph zone (v) (H&E: a, ×200; b–c″, ×400).
In the depression group, the vacuolations in the SGZs appeared in both the upper and lower limbs of the DG, many degenerated neurons exhibited darkly stained pyknotic nuclei, particularly in the upper limb and stem regions, and a few small-sized immature granular cells were observed (Figure 3(b′) and (c′)).
In the depression/exercise group, both limbs of the DG showed an apparently normal stratum granulosum with few or no vacuolations in the SGZs. The granular cells had normal rounded vesicular nuclei with many immature granular cells in between and in the SGZ. Few degenerated small cells were observed in the SGZ of several specimens, and numerous dilated blood vessels appeared in the pleomorph superficial zone (Figure 3(b″) and (c″)).
mPFC
The examination of the H&E stained sections of the mPFC from rats in the control group revealed apparently healthy groups of neurons arranged in the following six organised cortical layers: the molecular layer (LI), the outer granular layer (LII), the pyramidal cell layer (LIII), the inner granular layer (LIV), the ganglionic layer (LV) and the multiform layer (LVI). The granule cells had large rounded vesicular nuclei. The pyramidal cells possessed vesicular nuclei, a slightly basophilic granular cytoplasm and long peripheral processes. The glial cells appeared small with dark nuclei. The neuropil contained neuronal and glial cellular processes and blood vessels with a narrow perivascular space (Figure 4(a)–(c)).

Photomicrographs of sections in the mPFC of adult rat. (a–c) Specimens of control rat illustrate the organised layers of the mPFC; the molecular layer (LI), the outer granular layer (LII), the pyramidal cell layer (LIII), the inner granular layer (LIV), the ganglionic layer (LV) and the multiform layer (LVI). The granule cells have large rounded vesicular nuclei and prominent nucleoli (thin arrows). The pyramidal cells possess vesicular nuclei and long peripheral processes (thick arrows). The glial cells are small in size with dark nuclei (arrow heads). The neuropil contains neuronal and glial cellular processes (dotted arrows) and blood vessels (V). (d–f) Specimens of depressed rat show widening of the layer I (LI) and condensation of layer II (LII) and layer III (LIII). Many neurons appear irregular, shrunken, darkly stained and contain shrunken pyknotic nuclei and wide pericellular space (arrows). (g–i) Specimens of depression/exercise rat show nearly organised layers of the mPFC. Most of the granular (arrows) and pyramidal (thick arrows) cells appear normal. Few neurons appear shrunken and surrounded by haloes (arrow heads) (H&E: a, d, g, ×100; b, c, e–i, ×400).
In the depression group, the frontal cortex showed disorganised cortical layers, a widened LI, evident condensation and the merging of LII and LIII. Many neurons appeared irregular, shrunken and darkly stained and possessed shrunken pyknotic nuclei and wide pericellular spaces (haloes). Several neurons contained fragmented nuclei (Figure 4(d)–(f)).
The frontal cortical morphology in the depression/exercise group was restored towards a normal appearance as demonstrated by the apparently increased normal neurons and decreased perineuronal haloes. Numerous pyramidal cells and granular cells appeared normal, but a few neurons remained shrunken and surrounded by haloes (Figure 4(g)–(i)).
Cerebellum
The examination of the H&E stained sections of the cerebellum from rats in the control group revealed normal cerebellar folia within the outer cortex and inner medulla. The cortex, which constitutes the grey matter, comprises the following three layers: the outer molecular layer, middle Purkinje layer and inner granular cell layer (Figure 5(a)). The outer molecular layer contained basket cells, neuroglia, the large apical dendrites of Purkinje cells and several blood vessels. The Purkinje cell layer showed large flask-shaped cells typically arranged in a single row at the junction between the molecular layer and the granular layer, large rounded vesicular nuclei and prominent nucleoli. The granular cell layer comprised groups of small densely packed granular cells with darkly stained nuclei surrounded by very little cytoplasm (Figure 5(b)).

Photomicrographs of sections in cerebellum of adult rat. (a, b) Specimens of control rat illustrate cerebellar folia formed of an outer cortex and inner medulla. The cortex is layered into outer molecular cell layer (ML), middle Purkinje cell layer (PL) and inner granular cell layer (GL). The molecular layer contains basket cells (thin arrows), neuroglia (arrow head) and large apical dendrites of Purkinje cells (white arrows). The Purkinje layer contains large flask-shaped Purkinje cells (thick arrows) arranged in a single row with large rounded vesicular nuclei and prominent nucleoli. The granular cell layer contains groups of small densely packed granule cells (*) with darkly stained nuclei and little cytoplasm. (c) Specimen of depressed rat shows the molecular layer (ML) with apparently decreased neurons and nearly missed dendrites of Purkinje cells. The Purkinje cells appear shrunken with acidophilic cytoplasm and small deeply stained nuclei and are surrounded by pericellular spaces (arrows). The cells of the granular layer appear clumped with some pyknotic nuclei (curved arrow). (d) Specimen of depression/exercise rat shows the molecular layer (ML) with re-establishment of dendrites of Purkinje cells and presence of numerous blood vessels (V). Purkinje cells are seen with large rounded vesicular nuclei, prominent nucleoli and well apparent dendrites (arrows). Some cells are seen with small deeply stained nuclei (dotted arrow). The granular cell layer appears to retain the normal grouping appearance of cells (H&E: a, ×100; b–d, ×400).
The molecular level of the cerebellum of the rats in the depression group showed an apparently decreased number of neuronal cells and Purkinje cell dendrites. The Purkinje cell layer showed a marked degeneration, many shrunken Purkinje cells with an acidophilic cytoplasm and small deeply stained nuclei surrounded by marked pericellular spaces. The cells in the granular layer appeared clumped, and several cells exhibited pyknotic nuclei (Figure 5(c)).
After the voluntary exercise, the molecular layer in the depressed/exercise rats exhibited a marked restoration of the dendrites of the Purkinje cells and numerous blood vessels. The Purkinje cell layer had preserved cells with large rounded vesicular nuclei, prominent nucleoli and easily visualised dendrites, but several degenerated cells with small, deeply stained nuclei were also observed. The granular cell layer retained its normal grouping appearance of cells (Figure 5(d)).
Immunohistochemical study of the expression of myokines
BDNF immunoexpression
DG
In the DG of the control rats, strong nuclear and cytoplasmic BDNF immunoreactions were observed in numerous mature and immature granular cells in all regions. Many immature neurons in the SGZ and between the mature neurons appeared immuno-negative. In the depression group, both limbs of the DG exhibited negative reactions in the mature neurons, but several immunoreactions appeared in the immature neurons in the SGZ. After the voluntary running, a moderate immunoreaction was observed in nearly all mature and immature granular cells in both limbs (Figure 6(a)–(c)).

Photomicrographs representing the BDNF immunoexpression in sections of (a–c) the dentate gyrus, (d–f) the medial prefrontal cortex and (g–i) the cerebellum in the studied groups showing its neuronal expression with a marked decrease of the immunoexpression in the depression group, then its obvious increase in the depression/exercise group in all the studied brain areas (anti-BDNF immunostain ×400).
mPFC
In the mPFC of the control rats, moderate nuclear and cytoplasmic BDNF immunoreactions were observed in numerous granular and pyramidal cells in nearly all layers of the cortex. In the depression group, the reaction was markedly decreased, and only a few neurons exhibited faint immuno-positivity. After the voluntary running, a marked restoration of the immunoreactivity was observed, and many neurons exhibited mild reactions (Figure 6(d)–(f)).
Cerebellum
The BDNF immunoexpression in the cerebellum of the control rats showed strong nuclear and cytoplasmic reactions in numerous Purkinje and granular cells in the cerebellar cortex. After inducing depression, a marked decrease in immunostaining was observed, and only a few neurons expressed faint reactions. After the voluntary exercise, an obvious increase in the immunoexpression was observed in the granular and Purkinje cells and the axon-like fibres of the molecular layer (Figure 6(g)–(i)).
According to the OD measurements of the anti-BDNF stained sections of different brain areas, the BDNF expression in the depression group was significantly lower than that in the control group, and the BDNF expression in the depression/exercise group was significantly higher than that in the depression group (Figure 7(a), Table 1).

Graphs illustrating the optical density of myokines expression in brain regions of different groups (15 rats each): (a) anti-BDNF-stained sections, (b) anti-MIF stained sections, (c) anti-VEGF stained sections And (d) anti-IL-6-stained sections. X-axis shows the optical density and Y-axis shows the studied brain areas.
The means and the significance of the optical densities of different groups in different studied brain areas.

SE: standard error; DG: dentate gyrus; BDNF: brain-derived neurotrophic factor; mPFC: medial prefrontal cortex; MIF: migration inhibitory factor; VEGF: vascular endothelial growth factor; IL-6: interleukin-6; n: sample number.
Significance is considered at P > 0.05. P1 is the significance between control and depression groups. P2 is the significance between depression and depression/exercise groups.
MIF immunoexpression
DG
In the DG of the control rats, mild cytoplasmic MIF immunoreactivity was observed in the neurons of both limbs, particularly in the SGZ and immature granular cells. In the depression group, the neurons in both limbs of the DG exhibited negative reactions. Then, after the voluntary running, a mild immunoreaction was detected in the cells in the SGZ, particularly in the upper limb (Figure 8(a)–(c)).
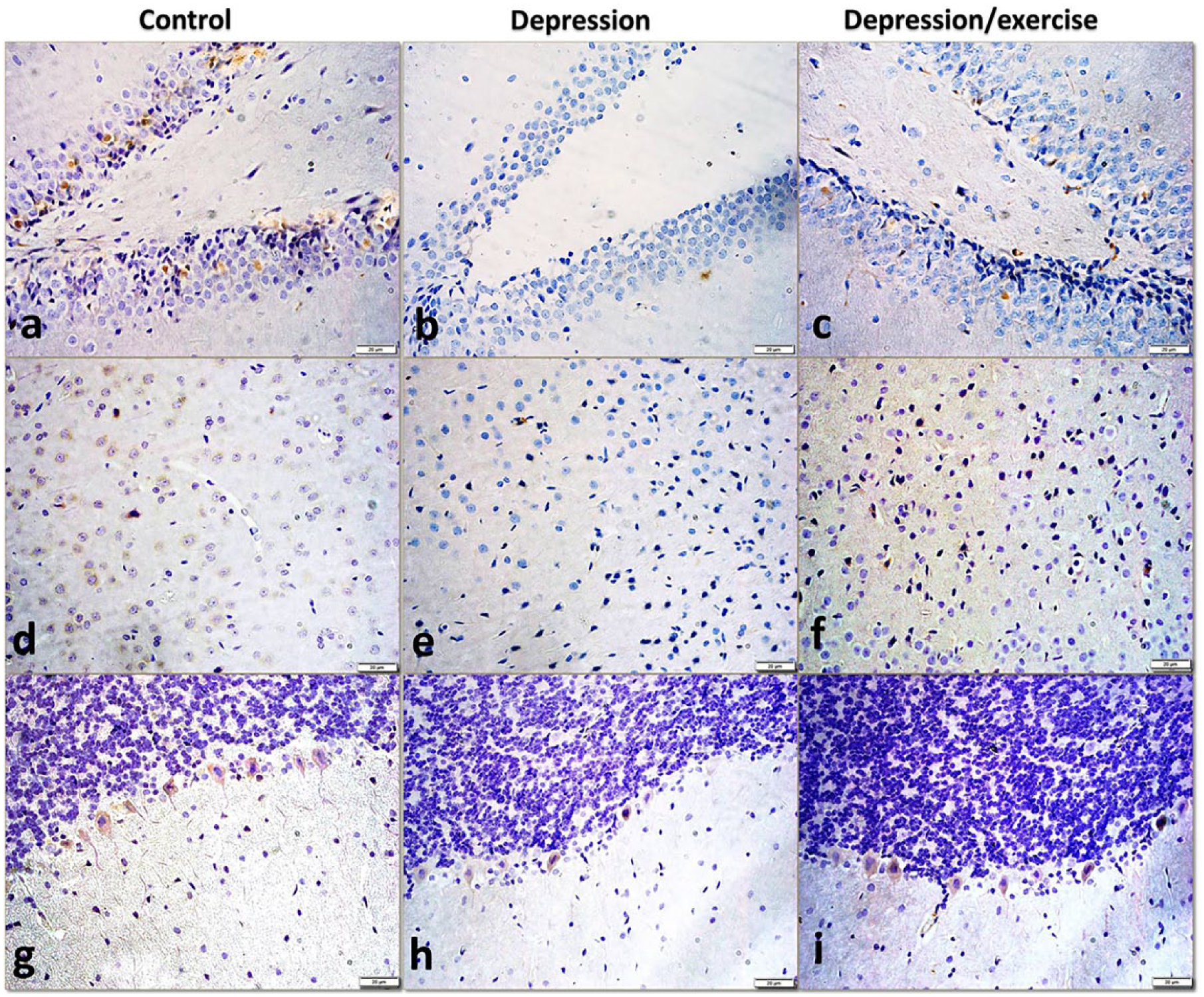
Photomicrographs representing the MIF immunoexpression in sections of (a–c) the dentate gyrus, (d–f) the medial prefrontal cortex and (g–i) the cerebellum in all the studied groups showing its neuronal expression with a markedly decreased or negative expression in depression group, and then its restoration in the depression/exercise group. Note the expression in the neuropil of the medial frontal cortex and in the Purkinje cells processes of the cerebellum in the depression exercise group (anti-MIF immunostain, ×400).
mPFC
In the mPFC of the control rats, a mild cytoplasmic immunoreaction was observed in many neurons. In the depression group, a negative immunoreaction was observed in nearly all cells. Then, in the depression/exercise group, an immunoreaction was detected in several neurons and the neuropil (Figure 8(d)–(f)).
Cerebellum
In the anti-MIF immunostained sections of the cerebellum of the control rats, a moderate immuno-positivity was observed in the cytoplasm and processes of the Purkinje cells. After inducing depression, only a weak reaction appeared in several Purkinje cells. Then, after the voluntary running, a moderate expression was observed in the Purkinje cells and their processes (Figure 8(g)–(i)).
According to the OD measurements in photographs of anti-MIF stained sections of the studied brain areas, the MIF expression in the depression group was significantly lower than that in the control group, and the MIF expression in the depression/exercise group was significantly higher than that in the depression group (Figure 7(b), Table 1).
VEGF immunoexpression
DG
The examination of the anti-VEGF immunostained sections of the DG of the control rats revealed strong immunoreactions in the cytoplasm of numerous granular cells in both limbs and the neuropil of both the molecular and polymorph layers. In the depression group, the expression was markedly decreased, and negative neurons were observed in both limbs of the DG. The stem region in several sections showed strong reactions. In the depression/exercise group, the immuno-positivity was obvious in the granular cells in both limbs and the molecular layer (Figure 9(a)–(c)).
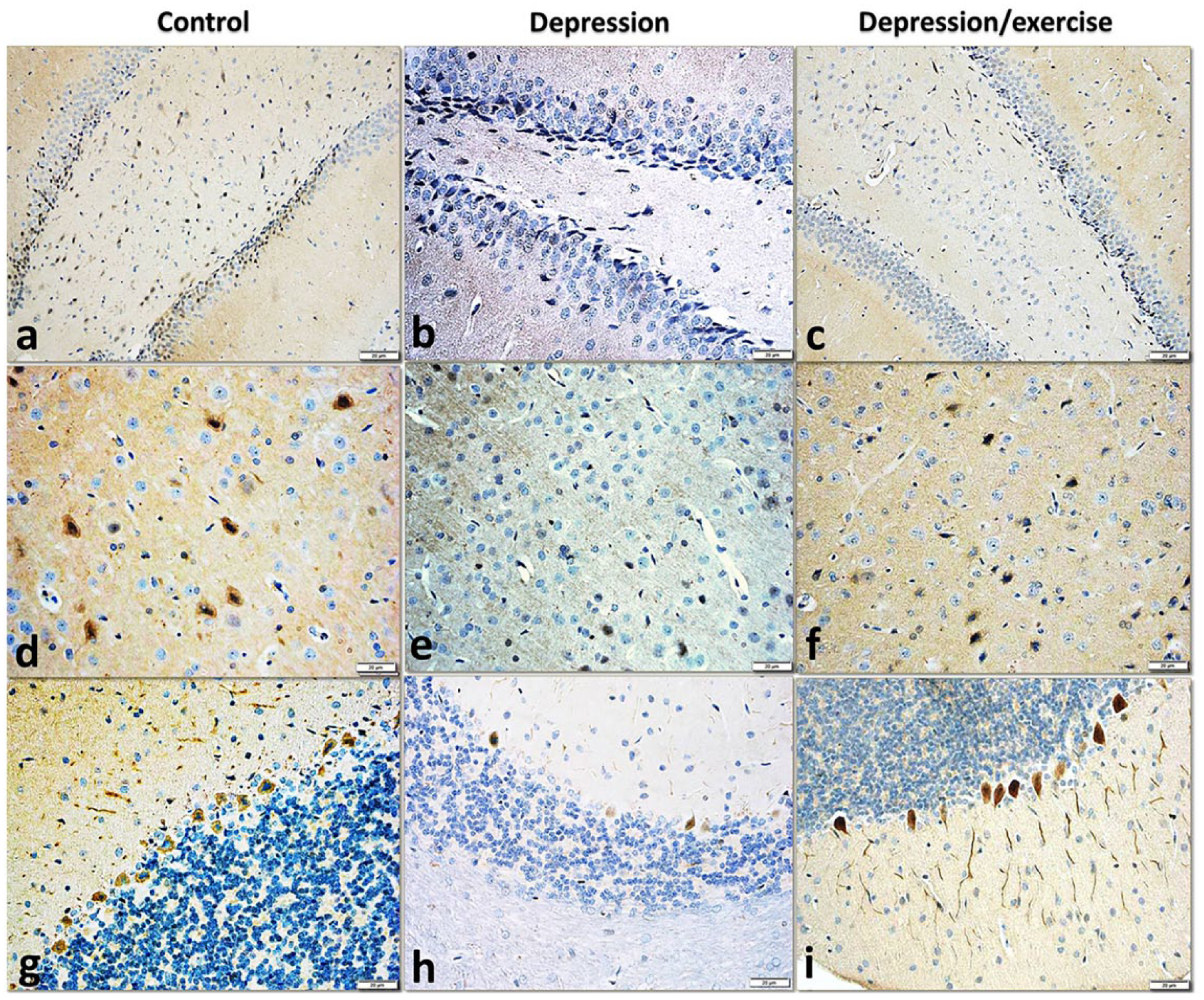
Photomicrographs representing the VEGF immunoexpression in sections of (a–c) the dentate gyrus, (d–f) the medial prefrontal l cortex and (g–i) the cerebellum in all the studied groups showing its neuronal expression with a marked decrease of the immunoexpression in the depression group, then its obvious increase in the depression/exercise group in all the studied brain areas. Note the increased immunoexpression in the neuropil of the medial frontal cortex and the granular layer of the cerebellum of the depression/exercise group (anti-VEGF immunostain, ×400).
mPFC
In the mPFC of the control rats, strong VEGF immunoreactions were observed in several neurons and the neuropil. In the depression group, a weak reaction was observed in many neurons, while in the depression/exercise sections, the restoration of the immuno-positivity was obvious in several neurons and the neuropil (Figure 9(d)–(f)).
Cerebellum
In the cerebellum of the control rats, a strong VEGF immunoexpression was observed in the Purkinje cells and their dendrites and the neuropil in the molecular layer. After inducing depression, only a few Purkinje cells were faintly stained. In the depression/exercise group, the Purkinje cells and their dendrites expressed strong immunoreactions. In addition, the neuropil in the granular cell layer showed strong reactions (Figure 9(g)–(i)).
According to the OD measurements of photographs of anti-VEGF stained sections, the VEGF expression in the depression group was significantly lower than that in the control group, and the VEGF expression in the depression/exercise group was significantly higher than that in the depression group in all studied areas (Figure 7(c), Table 1).
IL-6 immunoexpression
DG
The examination of anti-IL-6 immunostained sections of the DG of the control rats revealed a mild cytoplasmic immunoexpression in several granular cells, particularly in the SGZ and the neuropil in the molecular and polymorph layers. No IL-6 immunoexpression was observed in the DG of the rats in the depression group, but in the depression/exercise group, a mild cytoplasmic reaction was observed in the granular cells, particularly in the lower limb, and the reaction extended to the polymorph and molecular layers (Figure 10(a)–(c)).

Photomicrographs representing the IL-6 immunoexpression in sections of (a–c) the dentate gyrus, (d–f) the medial frontal cortex and (g–i) the cerebellum in all groups showing a marked decrease of the immunoexpression in the depression group, then its obvious increase in the depression/exercise group in all the studied brain areas. The expression appears in the neurons and neuropil of the dentate and cerebellum, but only in the neuropil, glial cells and endothelium of blood vessels of the medial frontal cortex (anti-IL-6 immunostain, ×400).
mPFC
A mild IL-6 expression was observed in the mPFC of the control rats, primarily in the neuropil, glial cells and epithelium of blood vessels. The nerve cells in the control sections appeared negative. In the depression group, only a few endothelial cells expressed IL-6 immunoreactions. In the depression/exercise group, the IL-6 immunoreaction was restored in the neuropil and glial cells. In addition, a strong reaction was exhibited by the endothelial cells of blood vessels (Figure 10(d)–(f)).
Cerebellum
In the cerebellum of the control rats, a moderate cytoplasmic IL-6 immunoexpression was observed in the Purkinje cells and their processes. Immuno-positivity was also observed between groups of cells in the granular and molecular zones. In the depression group, a weak reaction was expressed by a few Purkinje cells. In the depression/exercise group, a strong immunoreaction was detected in the cytoplasm of the Purkinje cells and their processes, and an immunoreaction also appeared in the neuropil in the granular zone (Figure 10(g)–(i)).
According to the OD measurements of photographs of anti-IL-6-stained sections, the IL-6 expression in the depression group was significantly lower than that in the control group, and the IL-6 expression in the depression/exercise group was significantly higher than that in the depression group in all studied brain areas (Figure 7(d), Table 1).
The correlation between values of optic density of different expressed myokines revealed significant correlation between the immune-expression of different myokines in all examined areas of the brain except for the non significant correlation found between MIF and BDNF in the lower limb of DG (P = 0.069) (Table 2).
Correlation between the optical densities of BDNF, VEGF, MIF and IL-6 in different areas of the brain in the studied groups.
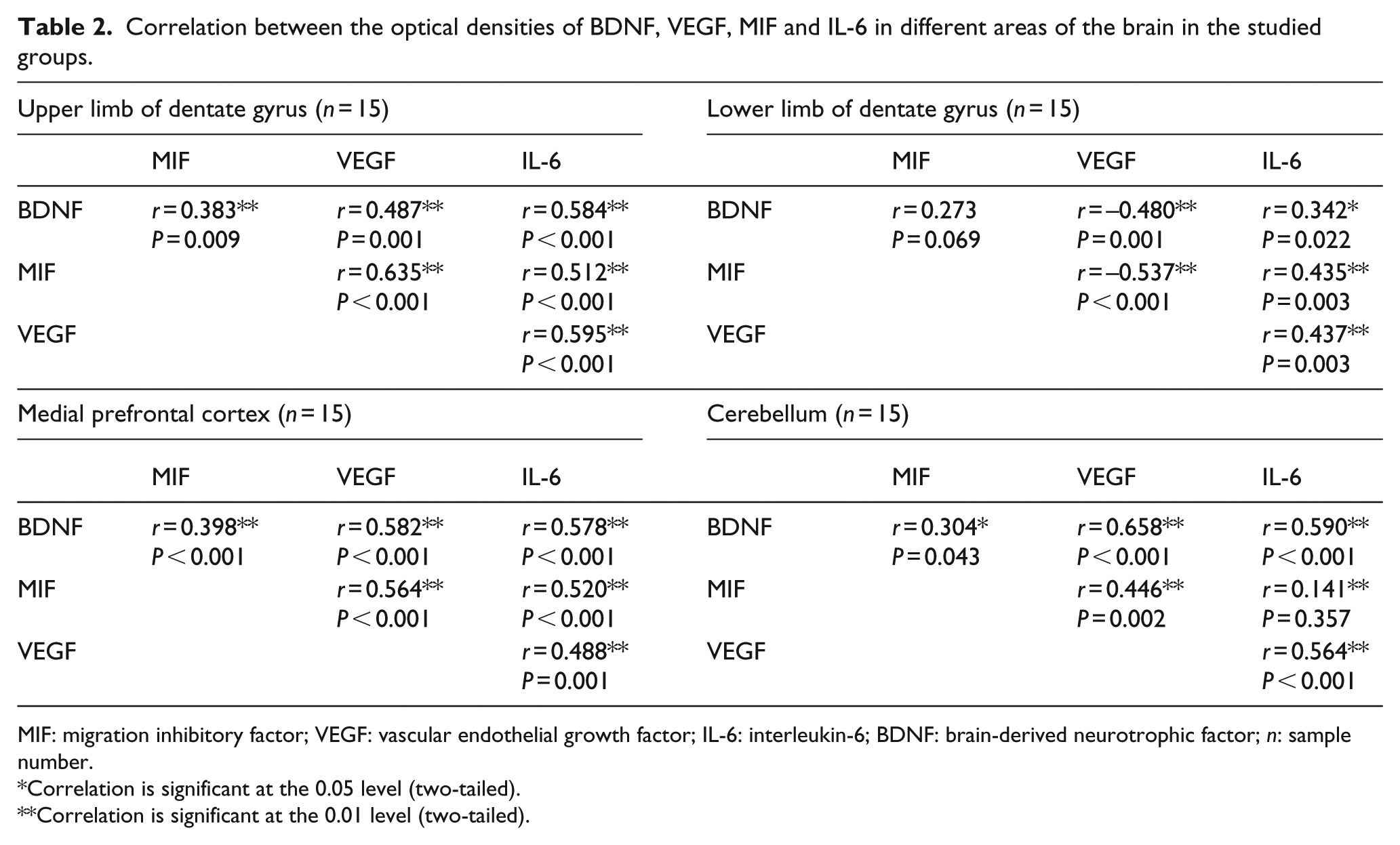
MIF: migration inhibitory factor; VEGF: vascular endothelial growth factor; IL-6: interleukin-6; BDNF: brain-derived neurotrophic factor; n: sample number.
Correlation is significant at the 0.05 level (two-tailed).
Correlation is significant at the 0.01 level (two-tailed).
Discussion
In this study, the induction of depression caused a marked degeneration in the DG, and voluntary running markedly repaired this degeneration. These changes could be due to the changed expression of neurotrophic and vascular neurogenic myokines in brain cells, which is consistent with our results.35–38
The histological study of the mPFC of depressed rats showed a disorganisation of the cortical layers and degeneration of many neurons. This finding is consistent with a recent study in which a reduced neuronal size and glial cell density were observed in the prefrontal cortex of subjects with major depression. 39 Also, some stereological studies have reported a decrease in cortical thickness in depression,40,41 suggesting neuronal loss along with a reduction in the neuropil. 42 Consistent with our hypothesis and findings, Rajkowska 43 suggested that neurotrophic factors play a role in the cellular changes that occur in the mPFC in depression.
This study revealed that exercise likely has a protective effect on the neurons and neuropil in the mPFC, which was evident by the restoration of the normal morphology. Although many studies have reported the exercise-induced proliferation and enhanced survival of hippocampal neurons in rodents,44–46 no reports thus far have shown the protective role of exercise on the mPFC.
In this study, the cerebellar cortex of the depressed rats showed a marked degeneration in the Purkinje cells with decreased dendritic extensions. This finding could denote the possible contribution of the cerebellum to the pathophysiology of depression and may be consistent with the reported cerebellar atrophy and reduced cerebellar size in patients with depression.47,48
After exercise, the restored dendrites of the Purkinje cells and the increased blood vessels in the molecular layer of the cerebellar cortex can denote the beneficial effect of running which is consistent with the previously reported enhancement of the synaptogenesis of Purkinje cells and improved motor performance after chronic treadmill exercise. 49 Nevertheless, this beneficial effect of exercise might not be mediated via neurogenesis because, in contrast to the hippocampus, many studies have failed to detect neurogenesis in Purkinje cells after exercise.50,51
Recently, physical exercise has proved to upregulate certain cytokines and immune cells while downregulating others. 2 Recently, many researchers have investigated the effects of different types of exercise on the expression of these myokines in brain areas. 2 In this study, we investigated the effect of voluntary running on the localization and the intensity of myokines in different brain areas.
The OD values of BDNF immune-expression in the DG, mPFC and cerebellum were significantly lower in the depression group compared to control one, and its values in the depression/exercise group increased significantly than in the depression group.
The downregulation of BDNF expression in brain areas of depressed rats highlights its role in the pathophysiology of depression 52 and is in consistent with some previous studies that reported its decrease in the hippocampus of depressed rats.53,54 On the other hand, its upregulation in these areas after voluntary running reflects the role of BDNF in alleviating depression symptoms, which is similar to the effect of antidepressant drugs in animals and human.55–59
The mPFC has recently been considered associated with depression and BDNF expression in response to different types of antipsychotic and antidepressant treatments.60–62 Therefore, the increased expression of BDNF in the mPFC detected in this study could be a contributing factor mediating the relieving effect of voluntary running on depression. This hypothesis is further supported by the findings that electroconvulsive therapy (ECT) alleviates symptoms of depression and increases the BDNF level in both the hippocampus and frontal cortex in a rodent model of depression.63–66 However, conflicting evidence exists regarding whether the increased expression of BDNF in the frontal cortex after chronic treatment of depression is a direct effect on the frontal cortex or an indirect effect caused by the remodelling of hippocampal neuronal terminations in the mPFC.67,68
The functional relationship between the cerebellum and brain areas associated with psychiatric diseases is well recognised.69,70 However, limited studies have provided evidence regarding the direct role of the cerebellum in depression.71,72 In our study, depression caused a marked decrease, whereas voluntary running resulted in a noticeable increase in the immunoexpression in the three cerebellar cortical layers. This changed expression of BDNF in the cerebellar cortex provides evidence of the role of the cerebellum in mood disorders.
The increased BDNF expression in the Purkinje cells and their dendrites after voluntary running is consistent with the improvement in the motor performance of rats after treadmill running38,49 and can reflect the promoting effect of voluntary exercise on synaptogenesis and neuronal networks in the cerebellum. 51
Recently, MIF has been considered a promising cytokine that plays a significant role in the pathophysiology of depression and has potential implications for depression management. 73 In this study, the OD values of MIF immune-expression, in all brain regions, decreased significantly in the depression group than in control one and significantly increased in the depression/exercise group than in depression one.
The decreased immunoexpression of MIF during depression is consistent with the increased depression-like behaviour in MIF knockout mice and the significant reduction in the number of proliferative cells in the DG after the pharmacological inhibition of MIF.74,75 The increased MIF expression after the voluntary running programme is consistent with the findings reported by Bloom and Al-Abed, who highlighted the central role of MIF in the exercise-mediated alleviation of depressive behaviour. 73
VEGF is considered the master regulator of angiogenesis. 76 Recently, the neuroprotective role of VEGF in psychiatric diseases has attracted attention.2,77 In this study, the OD values of VEGF immune-expression were significantly lower in depression group than that in control in all studied brain areas, and after voluntary running, the OD values increased significantly compared with the depression group in all areas.
The changed expression of VEGF during depression and after exercise might reflect the important role of this cytokine in signalling pathways engaged in behaviour and neurogenesis.77,78 The increased expression after the voluntary running is consistent with several recent studies79–81 and may reflect the neurotrophic action of this cytokine during exercise.2,82 This finding could be consistent with a new hypothesis that physical exercise triggers angiogenic genes83–85 and supports the suggestion of using pro-angiogenesis factors, including VEGF, in the treatment of neurodegenerative diseases. 86
IL-6 is commonly known as a proinflammatory cytokine, but several authors consider IL-6 an anti-inflammatory immunosuppressive factor. 87 Recently, IL-6 has been considered to play a role in depression by changing the connectivity of the hippocampus to the anterior cingulate cortex. 88 In this study, depression decreased the expression significantly in all studied brain areas, whereas voluntary running significantly increased the expression, but different patterns were observed in different brain areas.
The low expression of IL-6 in the studied brain areas and its localisation in glial and endothelial cells are consistent with a recent study reporting that IL-6 is primarily expressed in astrocytes and minimally expressed in neurons. 89 The changing expression of this myokine in the studied brain areas during depression and after the improvement of the depressive behaviour reflects its involvement in the pathophysiology of depression and the modulation of neurogenesis. 89
The significant correlation between the immune-expression of different myokines in all examined areas of the brain (DG, mPFC and cerebellum) can denote the harmony in reaction of these areas during induction and/or relief of depression and so, this can confirm the hypothesis that the mPFC and cerebellum are involved in the pathophysiology of depression.
In conclusion, this study explored the histopathological changes in the DG, mPFC and cerebellum both in depression and after voluntary running. To the best of our knowledge, this is the first study to immunohistochemically investigate the cellular expression of myokines (i.e. BDNF, MIF, VEGF and IL-6) and whether they are upregulated in particular areas of the brain after practising voluntary running. This study clearly reveals the involvement of the mPFC and cerebellum in the pathophysiology of depression.
Footnotes
Acknowledgements
The authors are so grateful to all the research technicians at King Fahd Centre for Medical Research, King Abdul-Aziz University, and at the Medical Research Laboratory of Taibah University for their help and support throughout the project.
Author’s contribution
All authors shared setting the design of the work, analysis and interpretation of the data. M.A.E. carried out the follow-up of all the practical work, photographing and analysis of photographs. M.A.E., Y.M.E, S.A.A., M.K.D and S.M.A. shared the writing and revising the draft critically. A.M.E. shared analysing the data and revising the draft. All authors approved the final manuscript.
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research authorship and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by grant number 34/127 from King Abdul-Aziz City for Science and Technology, Kingdom of Saudi Arabia.