
Abstract
Chronic pharyngitis is characterized as a common inflammation of the pharyngeal mucosa, and anti-inflammatory medications are the common treatment to relieve it. Polysacharides of Citrus grandis L. Osbeck (PCG) and luteolin have been reported to have anti-inflammatory activities. In this study, the protective effects of PCG and luteolin on chronic pharyngitis are evaluated and the underlying mechanisms are explored. PCG and luteolin are administrated to animal models with granuloma, ear edema and chronic pharyngitis and the effects of PCG and luteolin on disease severity are evaluated. We also evaluate the effects of PCG and luteolin on inflammatory cytokine production in macrophages stimulated with lipopolysaccharides (LPS)/interferon-gamma (IFN-γ) and detect the effects of PCG and luteolin on macrophage polarization. Finally, we evaluate the effects of PCG and luteolin on activations of LPS-induced downstream signaling pathways. PCG and luteolin alleviate the disease severity of granuloma, ear edema and chronic pharyngitis. PCG and luteolin suppress the productions of pro-inflammatory cytokines interlukin-6 (IL-6), interlukin-12 (IL-12) and tumor necrosis factor alpha (TNF-α) in macrophages. Luteolin promotes macrophage M2 polarization by enhancing expressions of arginase (Arg1) and mannose receptor C type 1 (Mrc1). PCG and luteolin suppress nuclear factor kappa-light-chain-enhancer of activated B cells (NF-κB) activation and interferon regulatory factor 1 (IRF1), interferon regulatory factor 5 (IRF5) expression. PCG together with luteolin relieves chronic pharyngitis by anti-inflammatory via suppressing NF-κB pathway and the polarization of M1 macrophage.
Introduction
Chronic pharyngitis (CP) is a common inflammation of the pharyngeal mucosa which caused significant problems in daily life, starting from throat discomfort in the morning to alteration of voice. 1 CP is usually caused by bacterial or viral infection, or by chemical or physical irritation.
There are various therapeutic strategies to treat CP. Among them, anti-inflammatory medications are commonly used. The anti-inflammatory and antioxidant activities of naturally occurring polysaccharides have been reported,2,3 suggesting the potential of polysaccharides to treat inflammatory disease. In China, Citrus grandis L. Osbeck is with polysaccharide component and possesses anti-inflammatory activities, and is widely used in traditional Chinese medicine to treat cough and promote digestion. 4 Luteolin is an important flavone, which is found in several plant products, including broccoli, pepper, thyme, and celery. The antioxidant and anti-inflammatory properties of luteolin have been reported, which depend on inhibiting activation of NF-κB and MAPK signaling pathways. 5 The aims of current study are to examine the potential effects of polysaccharides from Citrus grandis L. Osbeck (PCG) and luteolin on CP. We found that both PCG and luteolin reduced disease symptoms in rabbits with CP. PCG and luteolin also relieved disease severity in mice with ear edema and rats with granuloma.
Materials and methods
Extraction of polysaccharides from Citrus grandis L. Osbeck
The polysaccharides were extracted from Citrus grandis L. Osbeck as described previously. 1 Column chromatographic extraction with gradient elution followed by automatic separation was used to extract polysaccharides. Briefly, 5 g Citrus grandis L. Osbeck was put into a column by the wet column preparation method, with a minimum volume of solvent. The polysaccharides reached dynamic equilibrium in solution in 2 h. The same solvent was used to rinse the column, and the elutes were subsequently collected in fractions. Finally, the extraction mixture was adjusted to 80% ethanol and the polysaccharides was harvested by centrifuge at 5000 g for 20 min and then washed in 80% ethanol for twice.
Animals
The use of animals was conformed to the Guiding Principles in the Care and Use of Animals approved by the Yantai Yuhuangding Hospital. Adult male Wistar rats weighing 180–220 g, male ICR mice weighing 18–22 g and New Zealand white rabbits weighting 2.2–2.8 kg were used. Animals were housed 5 per cage and were provided with distilled water and food ad libitum, and kept under a 12 h light/dark cycle at constant temperature (22.5°C) and humidity (55%).
Cotton pellet-induced granuloma tissue formation test
We established the animal model of cotton pellet-induced granuloma tissue formation according to previous report. 6 First, rats were anesthetized using chloral hydrate (350 mg/kg). Two sterilized cotton pellets (20 mg) were implanted subcutaneously on each side of the nape through a small ventral incision in the nape of the animals. After the implantation of the cotton pellets, each rat treatment group (n = 5 per group) received topical treatment of vehicle (PBS), Aspirin (50 mg/kg) or PCG (2 mg/kg) and/or luteolin (20 mg/kg) per day for 1 week. Then, the rats were anesthetized with chloral hydrate and the implanted cotton pellets were removed with the surrounding fibrovascular tissue and dried at 60°C for 12 h. The dry weight was then measured, and the net granuloma weight was calculated by subtracting the original pellet weight from the dry pellet weight.
Xylene-induced mouse ear edema
The male ICR mice were randomly divided into five groups (n = 8). Mice were treated with vehicle, aspirin (50 mg/kg) or PCG (2 mg/kg) and/or luteolin (20 mg/kg). One hour after the treatment, ear edema was induced by applying 30 μL xylene on the inner surface of the right ear, while the left ear was used as control. Thirty minutes later, the mice were sacrificed with both ears removed and weighed. Edema was defined as the difference in weight between the two ears.
CP rabbit model
Rabbits were sprayed with 2.5% ammonia water into the pharynx mucosal twice per day (600 μL total) for 15 consecutive days. On day 8, 0.5 mL oil of turpentine was injected into the pharynx mucosal of the rabbits. The rabbits were randomly divided into the control, CP, PCG, luteolin and PCG + luteolin groups (n = 15 per group). Each treatment group received the respective treatment of vehicle, Aspirin (50 mg/kg) or PCG (2 mg/kg) and/or luteolin (20 mg/kg) per day for 14 consecutive days. After 24 h following the last administration, animals were anesthetized and pharyngeal tissue was removed and fixed in 4% formalin. The rabbits were anesthetized with urethane (1 g/kg). Tissues were sliced for hematoxylin and eosin staining to observe pathological differences between the groups under a light microscope (Olympus, Tokyo, Japan).
Macrophage treatment
Macrophages from peripheral blood of the male BALB/c mice were isolated by FACS sorting using anti-CD14 and F4/80 antibodies (Miltenyi Biotec Inc.CA, USA). 2×105 macrophages were plated in 24 well plate and treated with lipopolysaccharide (LPS) (500 ng/ml) and IFN-γ (100 ng/mL, Peprotech, USA) or IL-4 (20 ng/ml, R&D system, Minneapolis, MN, USA), together with PCG 20 µg/ml or luteolin (20 µM) or PCG/luteolin combination. Cells were collected at different time points after PCG, luteolin treatment for further analysis.
RT-PCR
Total RNA from macrophages was isolated using an RNeasy Mini kit (Qiagen, CA, USA) according to the manufacturer’s instructions. Reverse transcription was performed using a reverse transcription kit (Applied Biosystem, Waltham, MA, USA). Real time quantitative PCR reactions were set up in triplicate with SYBR® Green Master Mix (Biorad, CA, USA) and run on a LightCycler 480 (Roche, Penzberg, Upper Bavaria, Germany). The following primers were used in the current study: IL-6 (Fwd: 5’-CCTCTGGTCTTCTGGAGTACC-3’, Rev: 5’-AAGTGCATCATCGTTGTTCATACA-3’), TNF-α (Fwd: 5’-ATGAGCACAGAAAGCATGA-3,’ Rev: 5’-CTCCACTTGGTGGTTTGCTA-3’), IL-12 p35(Fwd: 5’-AGGACTTGAAGATG TACCAG-3,’ Rev: 5’-CTATCTGTGTGAGGAGGG-3’), IL-12 p40 (Fwd: 5’-GGAAGCAC GGCAGCAGAATAA-3,’ Rev: 5’-CTTGAGGGAGAAGTAGGAATG-3’), Arg1 (Fwd: 5’-CCTGAAGGAACTGAAAGGAAAG-3,’ Rev: 5’-TTGGCAGATATGCAGGGAGT-3’), Mrc1 (Fwd: 5’-GCAAATGGAGCCGTCTGTGC-3,’ Rev: 5’-CTCGTGGATCTCCGTGACAC-3’), Ym1 (Fwd: 5’-GGGCATACCTTTATCCTGAG-3,’ Rev: 5’-CCACTGAAGTCATCCATGTC-3’), Fizz (Fwd: 5’-AGAGGTGGAGAACCCAGCTTTGAT-3,’ Rev: 5’-TTTCAAGAAGCAG GGTAAATGGGCA-3’), β-actin (Fwd: 5’-AGTGTGACGTTGACAT CCGT-3,’ Rev: 5’-TGCTAGGAGCCAGAGCAGTA-3’).
Enzyme-linked immunosorbent assay
The levels of IL-6, TNF-α, IL-12 were measured by enzyme-linked immunosorbent assay (ELISA). ELISA kits were obtained from R&D systems (Minneapolis USA). Macrophage culture supernatants were harvested for ELISA analysis according to the manufacturer’s protocol.
Western blot
A total of 20 μg of proteins from whole cell lysate were loaded onto a 12% sodium dodecyl sulfate (SDS) polyacrylamide gel under denaturing conditions and were transferred to a polyvinylidene difluoride (PVDF) membrane (Bio-Rad, USA). Membranes were blocked with 10% non-fat dry milk dissolved in PBS with 0.1% Tween-20 overnight with rotation at 4°C. The following day, membranes were probed with p-IKKα/β, IKKβ, β-actin, IRF1, IRF5 and HSP60 antibodies for 2 hour at room temperature. The membranes were washed and probed with appropriate horseradish peroxidase conjugated secondary antibody. Chemiluminescent substrate (Thermo Fisher Scientific, Waltham, MA) was used to detect the bands. All antibodies were purchased from Abcam (Cambridge, MA, USA).
Statistical analysis
All data were presented as mean ± standard deviation (SD). Data were analyzed by one or two-way analysis of variance (ANOVA) analysis. Statistical difference was considered as significant when p value is less than 0.05.
Results
Protective effects of PCG and luteolin on granuloma and edema
We tested the anti-inflammatory effects of PCG and luteolin in our rat granuloma model and mice edema model. As shown in Figure 1(a), rats treated with PCG or luteolin had significantly lower granuloma weight when compared to vehicle treated rats. In addition, PCG/luteolin combination treatment enhanced the protective effects, as good as the positive control aspirin treated group, and the difference between PCG/luteolin treated rats and vehicle treated rats was more significant. Similarly, in mice edema model (Figure 1(b)), both PCG and luteolin protected mice from ear edema and the PCG/luteolin combination showed better effect than PCG or luteolin single treatment. Thus, our results suggested that both PCG and luteolin displayed anti-inflammatory effects in rat granuloma and mice edema.
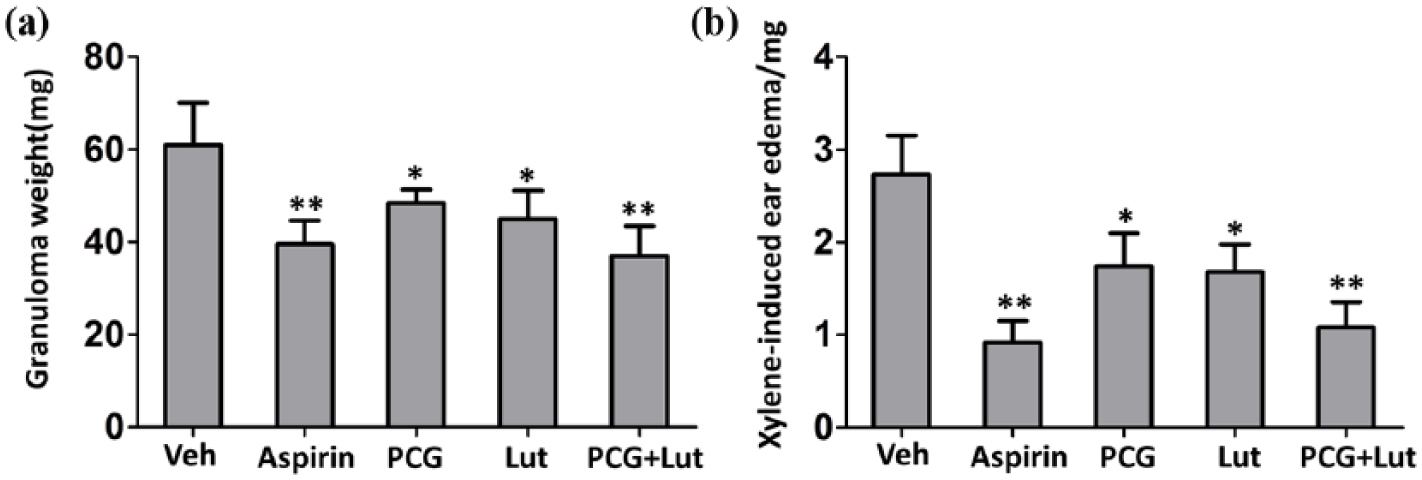
Anti-inflammatory effects of PCG and luteolin on rats granuloma and mice ear edema. (a) Granuloma weight and (b) Xylene-induced ear edema were analyzed. Results are expressed as the mean ± SD (n = 5). *P < 0.05 and **P < 0.01 compared to vehicle group.
Protective effects of PCG and luteolin on CP
To evaluate the possible protective effects of PCG and luteolin on CP, we established the rabbit CP model and then administrated PCG and/or luteolin to the rabbits. Then we compared the morphology differences. As summarized in Table 1, all CP rabbits displayed obvious inflammatory cell infiltration, vasodilatation and congestion, epithelial proliferation and glandular secretion in pharyngeal tissues while only few control rabbits displayed such symptoms. As predicted, PCG or luteolin treatment significantly decreased the numbers of rabbits with CP symptoms, indicating both PCG and luteolin protected rabbits from CP. In addition, PCG/luteolin combination treatment had better protective effects than PCG or luteolin single treatment, as the combination treatment resulted in the lowest numbers of rabbits with CP symptoms. Thus, our data suggested that both PCG and luteolin protected rabbit from CP.
Comparison of morphology of pharynx in different groups (n = 15).
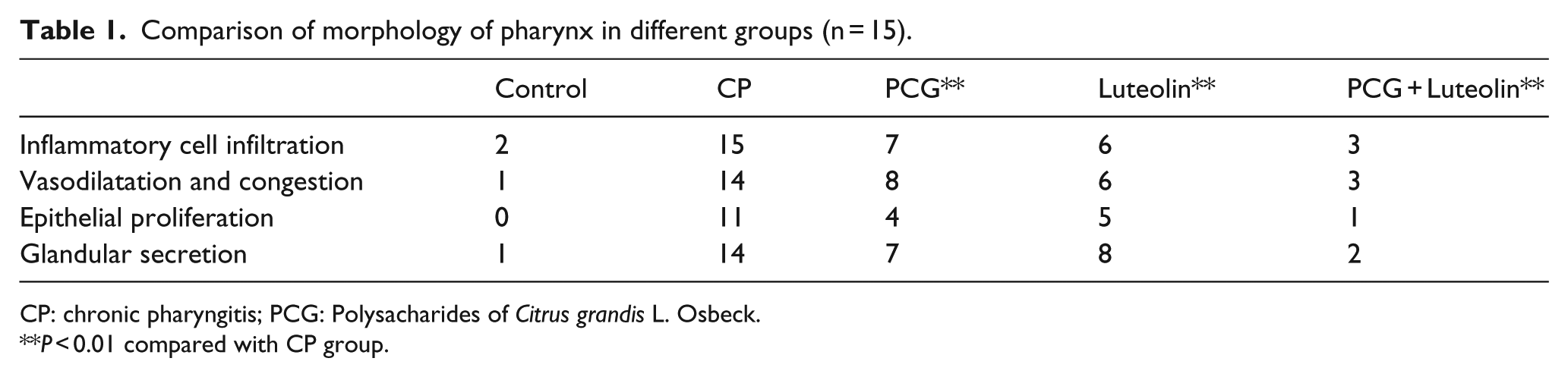
CP: chronic pharyngitis; PCG: Polysacharides of Citrus grandis L. Osbeck.
P < 0.01 compared with CP group.
PCG and luteolin inhibited inflammatory cytokines production in macrophages
Macrophages secreted several inflammatory cytokines, which played significant role in inflammation. We next tested the effects of PCG and luteolin on inflammation cytokines production in LPS and IFN-γ activated macrophages. We found that LPS and IFN-γ treatment induced IL-6 (Figure 2(a) and (e)), TNF-α (Figure 2(b) and (f)) and IL-12 (Figure 2(c), (d) and (g)) production in macrophages at both mRNA level (Figure 2(a)–(d)) and protein level (Figure 2(e)–(g)). Both PCG and luteolin significantly suppressed IL-6, TNF-α and IL-12 production in macrophage. The inhibitory effects were more significant in PCG/luteolin combination treatment.
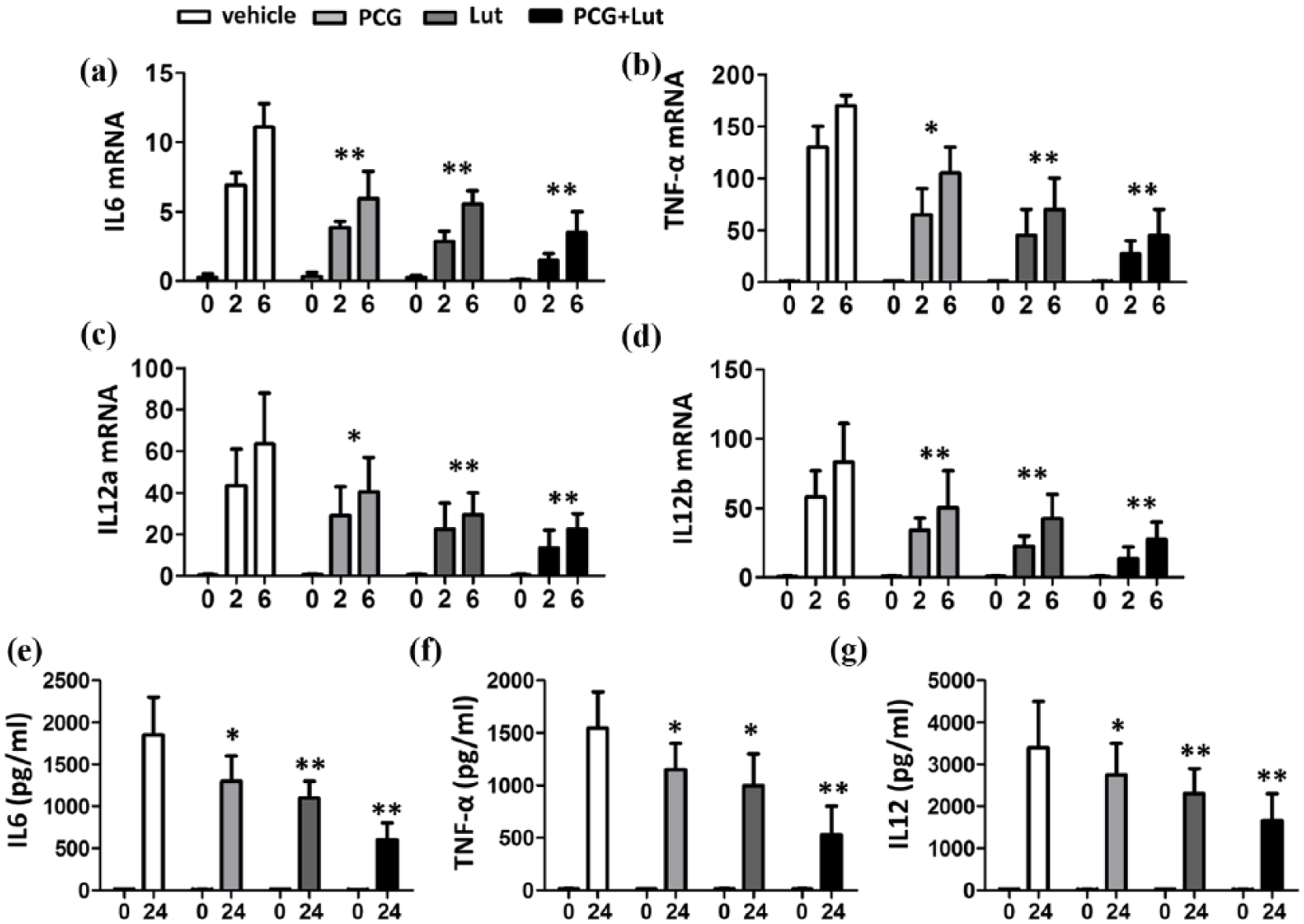
Effects of PCG and luteolin on IL-6, TNF-α and IL-12 expressions in macrophages. (a–d) qPCR analysis of the indicated genes using peripheral blood macrophages treated with PCG (20 µg/ml) and/or luteolin (20 µM) at 0, 2 and 6 h with IFN-γ and LPS stimulation. Data are presented as fold relative to the β-actin mRNA level. (e–g) ELISA of the indicated cytokines in the supernatants of peripheral blood macrophages treated with PCG and/or luteolin at 0 and 24 h with IFN-γ and LPS stimulation. Data are presented as mean ± SEM values and representative of at least three independent experiments. Statistical analyses represent variations in experimental replicates. *P < 0.05; **P < 0.01 compared with vehicle group.
Luteolin promoted Arg1 and Mrc1 expression in macrophages
LPS and IFN-γ polarized macrophages toward the M1 phenotype and induced production of inflammatory cytokines including TNF, IL-12, IL-1β. 7 As both PCG and luteolin suppressed LPS/IFN-γ induced inflammatory cytokines production in macrophages, we hypothesized that PCG and luteolin prevented macrophages M1 polarization while promoted M2 polarization. To test this hypothesis, we treated IL-4 polarized M2 macrophages with PCG and luteolin and detected the expression of M2 markers including Arginase (Arg1), Ym1, mannose receptor C type 1 (Mrc1) and Found in inflammatory zone (Fizz1). IL-4 induced expressions of Arg1 (Figure 3(a)), Ym1 (Figure 3(b)), Mrc1 (Figure 3(c)) and Fizz1 (Figure 3(d)). The application of PCG did not affect the IL-4 induced expression of all four genes. In contrast, luteolin alone or together with PCG significantly enhanced IL-4 induced expressions of all Arg1 and Mrc1, suggesting luteolin promoted macrophage polarization toward M2 phenotype.
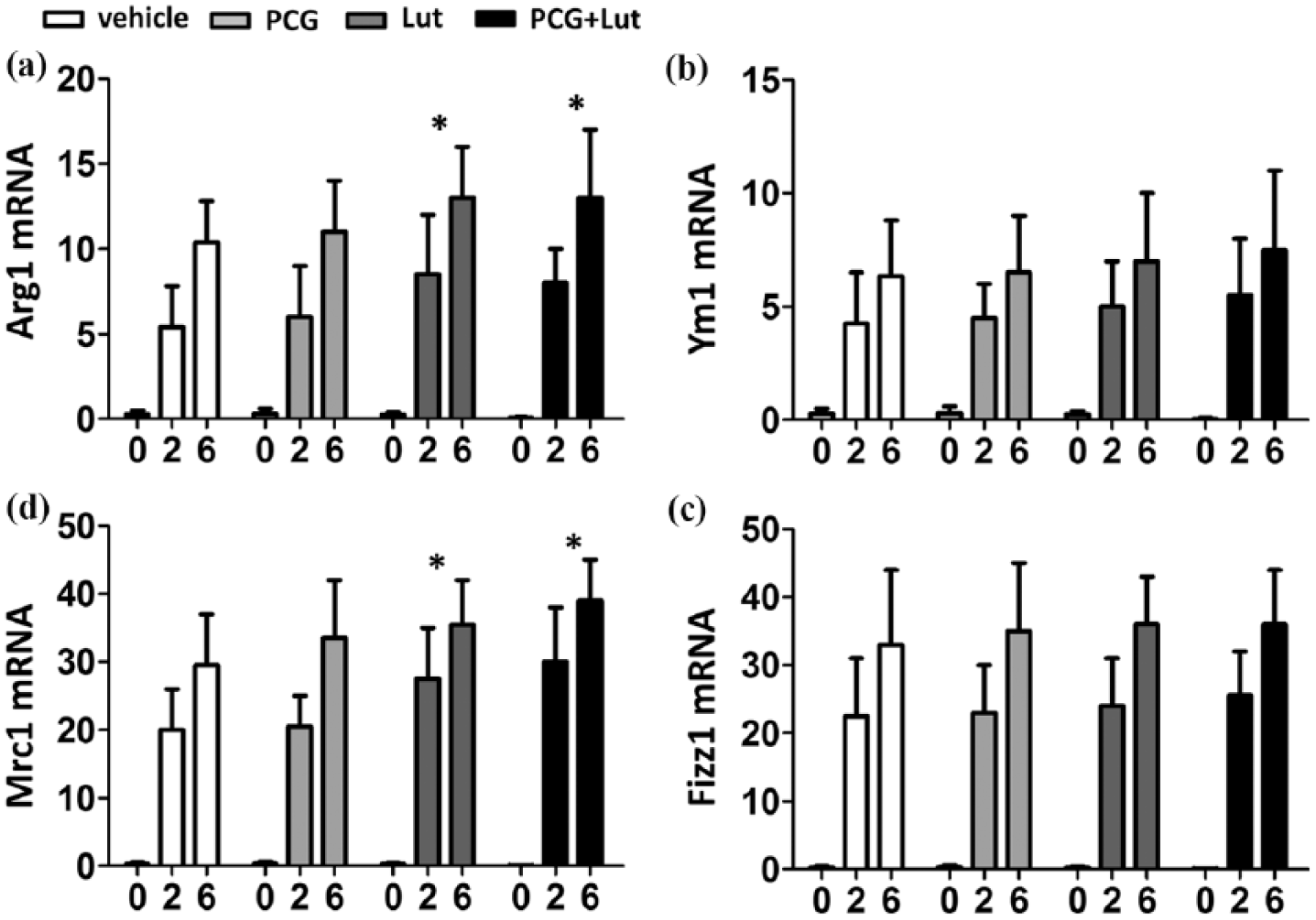
The expression levels of Arg1 and Mrc1 in macrophage were affected by luteolin. (a–d) qPCR analysis of the indicated genes using peripheral blood macrophages treated with PCG and/or luteolin at 0, 2 and 6 h with IL-4 stimulation. Data are presented as fold relative to the β-actin mRNA level. Data are presented as mean ± SEM values and representative of at least three independent experiments. Statistical analyses represent variations in experimental replicates. *P < 0.05; **P < 0.01 compared with vehicle group.
PCG and luteolin suppressed LPS-induced NF-κB activation and IRF-1 IRF-5 expression
It had been reported that NF-κB activation, IRF-1 and IRF-5 were involved in the commitment of proinflammatory M1 polarization.7,8 As PCG and luteolin had anti-inflammatory activities and suppressed M1 polarization, we tested the effect of PCG and luteolin on NF-κB activation and IRF-1, IRF-5 expression. As shown in Figure 4, LPS induced phosphorylation of IKK and expressions of IRF1 and IRF5. In the presence of luteolin or PCG, the phosphorylation of IKK was obviously decreased. Although PCG treatment did not affect IRF1 and IRF5 expression, Luteolin decreased expressions of both IRF1 and IRF5in LPS-treated macrophages, indicating the suppression of LPS-induced downstream signaling pathways. In addition, the PCG and luteolin combination had better suppression effects. Thus, our data suggested that PCG and luteolin suppressed NF-κB and IRF activation.
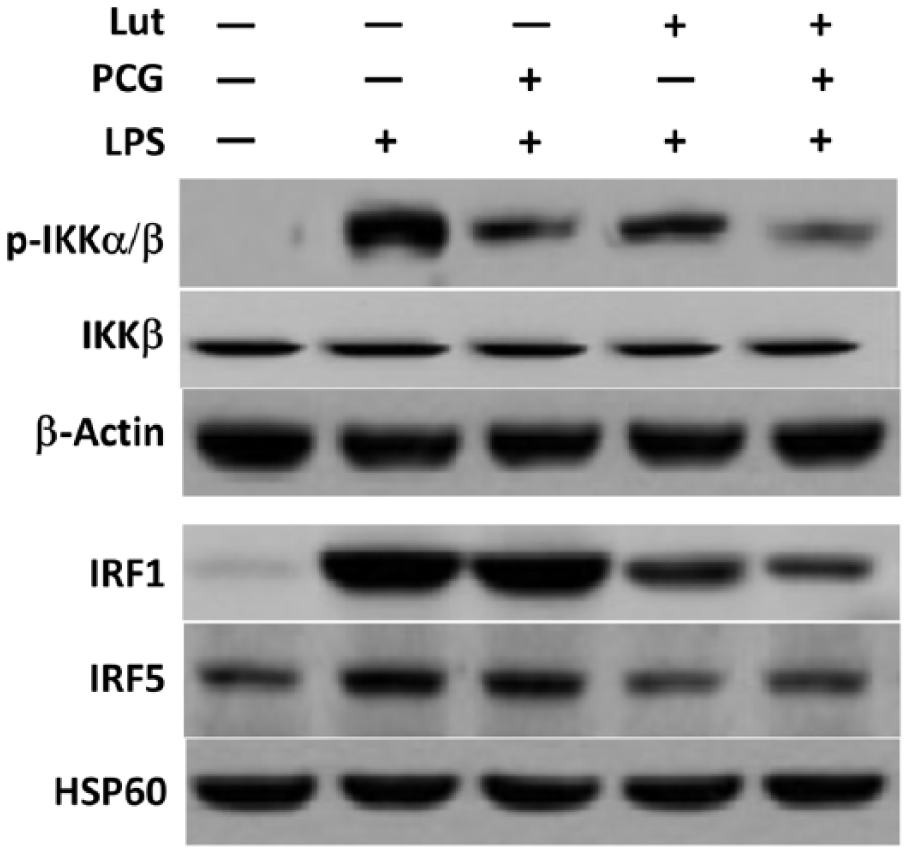
PCG and luteolin suppressed NF-κB activation and IRF1, IRF5 expression in macrophages. Peripheral blood macrophages were stimulated with LPS with or without PCG, luteolin. Cells were incubated for 6 h before harvest.
Discussion
Macrophages are highly versatile large white blood cells that play a central role in all stages of inflammatory response. 9 Once activated, macrophages secret a wide array of inflammatory cytokines and chemokines. Macrophages which are classically induced by T helper type 1 (Th1) cytokines, such as interferon (IFN)-γ, and bacterial LPS are known as M1 macrophages. They express high levels of CD86, as well as a profile of pro-inflammatory cytokines, such as IL-12, IL-6 and TNF-α. 10 In contrast, M2 macrophage activation is induced by fungal cells, immune complexes, macrophage colony stimulating factor (MCSF), IL-4, IL-13, IL-10 and tumor growth factor (TGF)-beta. This activation leads to the secretion of high amounts of IL-10 and low levels of IL-12. M2 macrophages exhibit anti-inflammatory functions. 11 Our results indicate the possible suppressive effects of PCG and luteolin on macrophage M1 polarization and that luteolin promoted macrophages polarization toward M2 phenotype. The NF-κB and IRF/STAT pathways, activated by IFNs and toll-like receptor (TLR) signaling, polarize macrophages to the M1 activation state via STAT1. In contrast, IL-4 and IL-13 skew macrophages toward the M2 activation state via STAT 6. 12 NF-κB is an important transcriptional factor regulating a wide variety of genes involved in inflammation. In canonical NF-κB pathway, stimuli lead to activation of the IKK complex by phosphorylation. Our data showed that PCG and luteolin suppressed phosphorylation of IKK, indicating the suppression of NF-κB activation. M1 macrophages have been shown to upregulate IRF1 and IRF5, which are critical for M1 polarization and the induction of IL-12, IL-23 and TNF. PCG and luteolin suppressed expression of both IRF1 and IRF5 in LPS treated macrophages.
Footnotes
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
This work was supported by grants from Yantai Municipal Scientific and Technological Development Project (2015WS027).