
Abstract
Background
The prevalence of multiple sclerosis is associated with the major histocompatibility complex class II DR15 haplotype HLA-DRB1*15:01∼HLA-DRB5*01:01.
Objective
To assess whether multiple sclerosis progression is associated with the main susceptibility haplotype HLA-DRB1*15:01∼HLA-DRB5*01:01.
Methods
Patients (n = 1230) and healthy controls (n = 2110) were genotyped for HLA-DRB1 and HLA-DRB5. The baseline Expanded Disability Status Scale (EDSS) score was determined and patients were followed for at least 3 years.
Results
After follow-up of the consecutive cohort 349 patients were classified as having clinical isolated syndrome and 881 patients as having multiple sclerosis. The susceptibility allele HLA-DRB1*15:01 was more frequent in clinical isolated syndrome (odds ratio 1.56) and multiple sclerosis (odds ratio 3.17) compared to controls. HLA- DRB1*15:01 was the only enriched HLA-DRB1 allele in multiple sclerosis patients. Comparison of clinical characteristics between HLA-DRB1*15:01∼HLA-DRB5*01:01 negative and positive patients with multiple sclerosis showed that baseline EDSS score, disease duration and frequency of the category secondary progressive multiple sclerosis with relapse were increased in the HLA-DRB1*15:01∼HLA-DRB5*01:01 positive group.
Conclusion
The study confirmed HLA-DRB1*15:01 and HLA-DRB5*01:01 as the main susceptibility alleles and showed weak indirect evidence for a role in progression of the disease.
Introduction
Multiple sclerosis (MS) is a debilitating autoimmune disease of the central nervous system predominantly in women and is caused by the combination of a complex genetic background and environmental trigger factors.1,2 The greatest individual genetic risk is conferred by the major histocompatibility complex (MHC) or human leukocyte antigen (HLA)-DR15 haplotype, which contains the two DRB alleles HLA-DRB1*15:01 and HLA-DRB5*01:01 in virtually 100% linkage disequilibrium.3,4 Based on data from many studies, the HLA-DR15 haplotype is now considered to confer, in Caucasians, up to 60% of the MS disease risk, while the known non-MHC risk markers account for approximately 20%.5–7 The two DR antigens generated by the HLA-DR15 haplotype are the heterodimer of HLA-DRA1*01:01/HLA-DRB1*15:01, also referred to as DR2b and the heterodimer of HLA-DRA1*01:01/HLA-DRB5*01:01, also referred to as DR2a. 8 This short HLA haplotype is part of the complete HLA-DR15 haplotype (HLA-DRA1*01:01∼HLA-DRB5*01:01∼HLA-DRB1*15:01∼HLA-DQA1*01:02∼HLA-DQB1*06:02). Furthermore, it has been shown that the impact of important environmental risk factors for MS (Epstein–Barr virus infection, cigarette smoking and low vitamin D3 serum level) is increased in HLA-DR15 haplotype carriers in a more than additive fashion.2,9
Various approaches have been pursued to assess the impact of the HLA-DR15 haplotype on disease progression, as reviewed by van der Walt et al. 10 In eight of 18 studies, no effect of the HLA-DR15 haplotype was seen, in six of 18 younger ages at diagnosis (onset) were reported, and in four of 18 marginally worse EDSS outcome was observed. The few studies that report an influence showed a protective effect of the HLA-DRB1*04 and the HLA-DRB1*01/HLA-DRB1*15 allele combination for the primary-progressive course, 11 an earlier conversion of HLA-DR15 carriers to secondary progressive multiple sclerosis (SPMS), 12 and a worse disease outcome. 13 HLA-DR15 and HLA-DR4 were associated with relapsing–remitting multiple sclerosis (RRMS) and SPMS, while primary progressive multiple sclerosis (PPMS) was additionally related to HLA-DR4/HLA-DQw8, HLA-DR7/HLA-DQw9 and HLA-DRw8/HLA-DQw4 haplotypes. 14 Numerous pharmacogenomic studies have explicitly examined the effect of HLA-DR allele status as well as other genes on treatment response of first-line therapies in MS. To date all have concluded that neither HLA-DR15 nor any other HLA allele showed an association with response to either interferon-beta or glatiramer acetate treatment (reviewed in Mahurkar et al.) 15
In this study we examined a large cohort of consecutive patients referred to a northern German centre for MS to address whether the HLA-DRB1*15:01∼HLA-DRB5*01:01 haplotype is associated with clinical progression.
Methods
Study subjects
A total of 1230 consecutive individuals was recruited between 1994 and 2010 at the MS outpatient unit and day care clinic of the University Medical Center Hamburg-Eppendorf, typed for HLA, and clinically evaluated at each visit; 2110 ethnically matched platelet and stem cell donors recruited between 1995 and 2010 at the Department of Transfusion Medicine served as controls. The local ethics committee (Ethik-Kommission der Ärztekammer Hamburg) approved the study (no. 2758) and written informed consent was obtained from all subjects.
HLA genotyping
DNA was extracted from ethylenediamine tetraacetic acid blood as described previously. 16 Samples were typed for HLA class II (DRB1 and DRB5) using the reverse sequence specific oligonucleotide dotblot method. Exon 2 for both genes was amplified and interrogated.17–19 Helmberg-Score software was used for allelic assignment in cooperation with the most recent nomenclature report and library.20,21 Remaining ambiguities were resolved to two-field allele resolution (DRB1) by sequencing of amplimers obtained after sequence-specific polymerase chain reaction with appropriate primers (Olerup/CareDx, Stockholm, Sweden).
Clinical evaluation
Clinical tests comprised the symbol digit modality test, 9-hole-peg-test, timed 25-footwalk and timed tandem walk, which were performed at each clinical visit. Performance was rated by the EDSS. 22 In Kaplan–Meier analysis, the time interval to progress from an EDSS score below 4 to an EDSS score below 9 served as the clinical endpoint.
Statistical analysis
Allele carrier frequencies were calculated by gene counting. 23 The control cohort was in Hardy–Weinberg equilibrium (see Supplementary Table 1). Odds ratios (ORs) were calculated to measure MS risk. Significance was evaluated by two-sided t-test for continuous variables and Fisher’s exact test or Pearson test for categorical variables. Kaplan–Meier curves were tested by log-rank and hazard ratios by likelihood ratio. Values of P≤0.05 were considered statistically significant. P values were corrected when appropriate.
Results
Follow-up of the consecutive cohort of 1230 patients revealed that 349 patients had clinically isolated syndrome (CIS) and 881 patients had MS (Table 1). The phenotypic frequency of the MS susceptibility allele HLA-DRB1*15:01 increased from 26.7% in controls to 38.0% in CIS and 53.8% in MS. A similar increase was seen for HLA-DRB5*01:01 from 26.5% in controls to 53.0% in MS (see Supplementary Table 2). The HLA-DRB1*15 allelic group was the only one to be singled out as enriched in MS over controls (Figure 1). Significantly underrepresented HLA-DRB1 allelic groups were HLA-DRB1*07, 08, 11, 13 and 14. After Bonferroni correction only the HLA-DRB1*01 group remained significantly reduced, which reflects the smaller confidence interval of the OR and the higher prevalence of this group in German Caucasians as compared with, for example, the HLA-DRB1*09 group. To address further the status of these ‘alternative alleles’ we stratified the MS risk analysis for the DRB1*15:01 carrier status (see Supplementary Table 3). This analysis revealed no significant protective effect of any DRB1 allelic group in the non-DRB1*15:01 carriers except for the DRB1*07 allelic group. However, the effect was weak (OR 0.71) and not significant when corrected for multiple comparisons. Interestingly, DRB1*07 was significantly increased in the DRB1*15:01 carriers (OR 2.60) and this remained significant after Bonferroni correction. Thus the highest MS risk was observed in DRB1*15:01, 07 heterozygotes (OR 5.05).
HLA-DRB1*15:01 allele and phenotypic frequencies in multiple sclerosis, CIS and German controls.

CIS: clinically isolated syndrome.

Forrest plot of odds ratios (ORs) for multiple sclerosis (MS) risk according to HLA-DRB1 allelic group. HLA-DRB1*15 risk group in red, neutral allelic groups (*03, *04, *09, *10, *12, *16) in yellow, underrepresented and potentially protective allelic groups (*07, *08, *11, *13, *14) in green. OR and 95% confidence intervals (CIs); Bonferroni corrected P (pcor).
Next we compared the clinical characteristics between HLA-DRB1*15:01 negative and positive patients with MS to look for confounding variables (Table 2). There was no difference in age, sex and frequency of MS categories between the two groups. Significant but small differences were observed for higher baseline EDSS scores and longer disease duration in the HLA-DRB1*15:01∼HLA-DRB5*01:01 positive group. To analyse further the trend seen for SPMS we compared the frequency of MS categories and subcategories (Table 3). Here again an increased frequency of relapsing SPMS was observed for the HLA- HLA-DRB1*15:01∼HLA-DRB5*01:01 positive group.
Comparison of clinical characteristics between HLA-DRB1*15:01 negative and positive patients with multiple sclerosis.

Two-sided t-test for continuous variable, Fisher’s exact test for categorical variable, uncorrected P.
EDSS: Expanded Disability Status Scale; MS: multiple sclerosis.Note. Bold value represents P ≤ 0.05.
Multiple sclerosis category and DRB1*15:01 status (N=881).
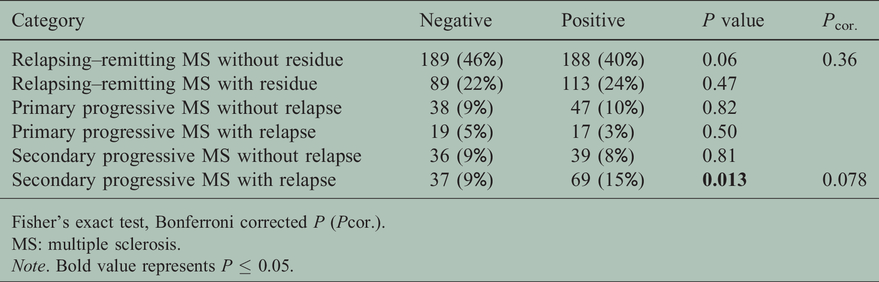
Fisher’s exact test, Bonferroni corrected P (Pcor.).
MS: multiple sclerosis.Note. Bold value represents P ≤ 0.05.
In 669 patients with MS and complete neurological examination, the distribution of low (0–4.5) and high (5–8.5) baseline EDSS scores correlated with the MS category (Table 4) and the presence of HLA-DRB1*15:01∼HLA-DRB5*01:01 (Table 5). MS patients with high-level baseline EDSS scores were more frequently positive for the susceptibility HLA-DRB1*15:01∼HLA-DRB5*01:01 haplotype. It is of note that SPMS with relapse stands out with the highest baseline EDSS (Table 4), which in turn will reduce the probability of further increasing in follow-up. To correct for this we decided to use the time for change from the lower EDSS score group to the higher EDSS score group as the clinical endpoint in Kaplan–Meier analysis.
Multiple sclerosis category and baseline EDSS groups (total N=669).

Pearson test.
MS: multiple sclerosis.
Baseline Expanded Disability Status Scale (EDSS) groups and HLA-DRB1*15:01 status.

Fisher’s exact test.
Kaplan–Meier analysis for disease progression from baseline EDSS score less than 4 (0–3.5) to EDSS score less than 9 (4–8.5) confirmed most of these findings (Figure 2). While disease progression occurred in 25% of the cases after 8.9 years in RRMS, this happened in PPMS and SPMS after 3.4 and 2.3 years, respectively (Figure 2(a)). The univariate hazard ratio for progression between SPMS versus RRMS was 3.6 (95% CI 2.4–5.5). Men had a higher risk of progression than women (Figure 2(b)), with a univariate hazard ratio of 1.5 (95% CI 1.1–2.1). The presence of HLA-DRB1*15:01∼HLA-DRB5*01:01 had no effect on progression, either in the combined data (Figure 2(c)) or in the RRMS subset (Figure 2(d)), as well as the PPMS and SPMS subsets (data not shown).

Kaplan–Meier plots of Expanded Disability Status Scale (EDSS) progression stratified for (a) multiple sclerosis (MS) category, (b) sex, (c) HLA-DRB1*15:01 status in the total cohort (N = 434) and (d) in the subset of patients with relapsing–remitting multiple sclerosis (RRMS). PPMS, primary progressive MS; SPMS, secondary progressive MS.
Discussion
The analysis of this single-centre northern German MS patient cohort confirmed that HLA-DRB1*15:01 and HLA-DRB5*01:01 are the most prominent MS risk alleles. The finding of an enrichment of HLA-DRB1*15:01 and HLA-DRB5*01:01 in CIS and MS, the higher baseline EDSS scores and longer disease duration in the HLA-DRB1*15:01∼HLA-DRB5*01:01 positive group suggests that the HLA-DRB1*15:01∼DRB5*01:01 haplotype weakly influences the clinical progression of MS. However, this could not be confirmed in the Kaplan–Meier analysis for progression.
Despite its long and well-known association with MS, the mechanism by which HLA-DR15 confers disease susceptibility is not yet understood. Recent data indicate increased T-cell proliferation in MS patients carrying the HLA-DR15 allele 24 and an increased risk of the presence of oligoclonal bands in the cerebrospinal fluid, 25 nevertheless further studies are needed to shed light on the underlying mechanism and its potential modulation by environmental factors. Environmental agents such as vitamin D, viral infections, diet and noxious agents 2 have been reported to contribute to MS risk in general, and in the case of cigarette smoking 9 and obesity 26 to enhance MS risk synergistically in combination with HLA-DR15. Cigarette smoking has recently also been associated with a worse disease outcome in EDSS parameters. 27 The impact of vitamin D on the clinical course of MS has yet to be examined in clinical studies. Following the hypothesis of a disease-relevant expression of an alternative vitamin D response element (VDRE) in the promoter region of HLA-DR15 allele carriers, it has been suggested that a vitamin D deficiency in early life or in utero allows autoreactive T cells of HLA-DR15 allele carriers to escape central thymic deletion. Nolan and colleagues showed the presence of the constant VDRE in all HLA-DRB1 alleles yet they discussed the potentially protective effect on MS risk in HLA-DRB1*04, *07, *09 (DR53 group) allele carriers attributable to the non-responsive VDRE motif that they had detected in these alleles. 28 In contrast to these studies we did not observe a protective effect of the DR53 group associated alleles in our cohort, as only HLA-DRB1*07 showed a trend for underrepresentation in our MS patient cohort. HLA-DRB1*04 showed an equal distribution in controls and MS patients and only a few cases in our cohort were HLA-DRB1*09 carriers leading to a wide CI. In conclusion, there was no single dominant protective HLA-DRB1 group.
The influence of pairs of classic HLA alleles in strong linkage disequilibrium, HLA-DRB1*15:01∼HLA-DQB1*06:02 and HLA-DRB1*01:01∼HLA-DQA1*01:01, on age at onset has recently been shown in a large cohort. 29 In addition, evidence for a strong protective effect of HLA-DQA1*01:01 in the presence of HLA-DRB1*15:01 and an allelic interaction between HLA-DQB1*03:01 with HLA-DQB1*03:02 was found. Re-examining our cohort of MS patients, we found HLA-DQA1*01:01 in combination with HLA-DRB1*15:01 in 39 of 881 patients and HLA-DQB1*03:01 with HLA-DQB1*03:02 in 18 of 881 patients. Further statistical analysis showed no significant correlation with age of onset or clinical outcome parameter in these patient subgroups. With respect to magnetic resonance imaging lesion load and histopathological findings, more extensive CNS inflammation has been described in HLA-DR15 haplotype carriers based on small MS cohorts 30 and recently also in PPMS. 31 However, these data have not been confirmed in larger cohorts.32,33
In the era of disease-modifying therapy (DMT) for MS, it has become even more difficult to assess disease progression (i.e. Kaplan–Meier analysis using an increase in EDSS as a clinical endpoint). This is because, along with possible effects of the DMT, the baseline EDSS is highly associated with the MS category. Therefore analysis must be stratified accordingly, which reduces the power of the present study by reducing the total N from originally 1230 subjects to 434 subjects, who showed EDSS score-progression (Figure 2(a)–(c)) to 272 subjects with RRMS (Figure 2(d)). Under these circumstances, we have no direct evidence for an impact of HLA-DRB1*15:01∼HLA-DRB5*01:01 on disease progression. We predict that, if the indirect evidence we see were to become manifest in a larger prospective study controlled for DMT, the effect would be weak, approximately in the range of the effect seen for the gender. In conclusion, apart from MS category and patient sex we could not identify HLA-DRB1*15:01∼HLA-DRB5*01:01 as a predictor for clinical progression.
Although this is the largest single-centre MS patient cohort to be evaluated for the HLA-DRB1*15:01∼HLA-DRB5*01:01 haplotype as a biomarker to predict clinical progression,10,33 the study is limited by its retrospective design and lack of data on HLA class I and non-MHC modifying markers. In particular, the impact of the protective HLA-A*02:01 allele 34 and the risk-increasing HLA-A*03:01 allele, as well as the presence of other recently identified MS-relevant genes are not included. 6 It remains unclear to what extent these genes, alone or in combination, contribute to disease progression. Finally, due to the retrospective design and lack of a mandatory schedule for clinical follow-up, we cannot exclude a bias towards worse disease development as patients with more severe disease activity are more likely to visit the centre more frequently.
Conclusions
For a northern European group, we here confirm the HLA-DRB1*15:01∼HLA-DRB5*01:01 haplotype as the single most influential disease susceptibility marker in MS and show evidence for its weak impact on clinical disease progression.
Supplemental Material
MSO894615 Supplemental Material - Supplemental material for Is multiple sclerosis progression associated with the HLA-DR15 haplotype?
Supplemental material, MSO894615 Supplemental Material for Is multiple sclerosis progression associated with the HLA-DR15 haplotype? by Klarissa Hanja Stürner Inessa Siembab Gerhard Schön Jan-Patrick Stellmann Nika Heidari Boris Fehse Christoph Heesen Thomas H Eiermann Roland Martin Binder Thomas MC in Multiple Sclerosis Journal—Experimental, Translational and Clinical
Footnotes
Acknowledgments
The author(s) thank W Fehrle for help with revision of the manuscript.
Conflict of Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: RM was supported by an ERC advanced grant (ERC-2013-ADG 340733).
ORCID iDs
Klarissa Hanja Stürner https://orcid.org/0000-0003-2199-3315 Christoph Heesen https://orcid.org/0000-0001-8131-9467 Thomas H Eiermann ![]()
Supplemental material
Supplemental material for this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.