
Abstract
Background:
Shroom family member 3 (SHROOM3) encodes an actin-associated protein that regulates epithelial morphology during development. Several genome-wide association studies (GWAS) have identified genetic variances primarily in the 5’ region of SHROOM3, associated with chronic kidney disease (CKD) and poor transplant outcomes. These genetic variants are associated with alterations in Shroom3 expression.
Objective:
Characterize the phenotypic abnormalities associated with reduced Shroom3 expression in postnatal day 3-, 1-month and 3-month-old mice.
Methods:
The Shroom3 protein expression pattern was determined by immunofluorescence. We generated Shroom3 heterozygous null mice (Shroom3Gt/+) and performed comparative analyses with wild type littermates based on somatic and kidney growth, gross renal anatomy, renal histology, renal function at postnatal day 3, 1 month, and 3 months.
Results:
The Shroom3 protein expression localized to the apical regions of medullary and cortical tubular epithelium in postnatal wild type kidneys. Co-immunofluorescence studies confirmed protein expression localized to the apical side of the tubular epithelium in proximal convoluted tubules, distal convoluted tubules, and collecting ducts. While Shroom3 heterozygous null mice exhibited reduced Shroom3 protein expression, no differences in somatic and kidney growth were observed when compared to wild type mice. Although, rare cases of unilateral hypoplasia of the right kidney were observed at postnatal 1 month in Shroom3 heterozygotes. Yet renal histological analysis did not reveal any overt abnormalities in overall kidney structure or in glomerular and tubular organization in Shroom3 heterozygous null mice when compared to wild type mice. Analysis of the apical-basolateral orientation of the tubule epithelium demonstrated alterations in the proximal convoluted tubules and modest disorganization in the distal convoluted tubules at 3 months in Shroom3 heterozygotes. Additionally, these modest abnormalities were not accompanied by tubular injury or physiological defects in renal and cardiovascular function.
Conclusion:
Taken together, our results describe a mild kidney disease phenotype in adult Shroom3 heterozygous null mice, suggesting that Shroom3 expression and function may be required for proper structure and maintenance of the various tubular epithelial parenchyma of the kidney.
Introduction
Genome-wide association studies (GWAS) have identified significant associations between genetic variants in the SHROOM3 gene, and the risk for chronic kidney disease (CKD).1 -3 There are 4 Shroom family member proteins implicated in various morphological processes, but Shroom3 is considered unique as it is the only member capable of initiating apical constriction. 4 Traditionally, Shroom3 modulates the direct recruitment and subcellular organization of F-actin and Rho-kinase to induce epithelial cell shape changes through apical constriction.5 -7 This role in actomyosin regulation is important for epithelial morphogenesis of essential organs ranging from the gut, lens, neural tube, heart, and in our focus, the kidneys.8 -10 Moreover, Shroom3 is implicated in various developmentally important pathways including apicobasal cell elongation, planar cell polarity signaling, and rosette formation among others.8,11,12
Despite the strong associations of Shroom3 with kidney disease, its role in renal epithelial maintenance, repair, and development is not well established. To outline potential roles for Shroom3 in the kidneys, functional genetic variation studies have begun investigating the impact of altered Shroom3 expression on kidney development, function, and disease. These studies have found significant associations between genetic variants in SHROOM3 and CKD, baseline estimated glomerular filtration rate (eGFR), urinary albumin-to-creatinine ratio (uACR), and blood urea nitrogen.3,13 -15 Studies investigating mouse models with varying Shroom3 expression levels support these findings and attempt to uncover the cause of these significant associations to further understand the role of Shroom3 in the kidneys. For example, during kidney development, it was established that Shroom3 plays a role in regulating glomerular development and is required for normal podocyte architecture and function. This has been established in studies that characterized the embryonic phenotype of Shroom3 homozygous null mice showing glomerular abnormalities and disruptions in podocyte cytoarchitecture during embryogenesis and a reduction in glomerular number at embryonic (E) day 18.5. 16 However, the investigation behind the phenotype of Shroom3 homozygous null mice was not extended to the postnatal adult kidney as these mice were embryonically lethal due to neural tube morphogenesis failure resulting in exencephaly. 17 Therefore, studies analyze Shroom3 heterozygous null mice postnatally to investigate the effects of reduced Shroom3 expression in adult mice kidneys. The postnatal Shroom3 heterozygous null mice phenotype at 1-year-old exhibits focal segmental glomerulosclerosis with thickened Bowman’s capsules, albuminuria, and podocyte foot process effacement.16,18 Several other studies analyzing Shroom3 genomic variants have also demonstrated podocyte cytoskeletal abnormalities leading to albuminuria and CKD.18 -20 Furthermore, in a model of acute kidney injury resulting from ischemia reperfusion, 3-month old Shroom3 heterozygous null mice exhibited poor tissue recovery as a result of alterations in epithelial repair and redifferentiation after kidney injury. 21 Studies have also demonstrated poor renal allograft function and interstitial fibrosis from donor kidneys that exhibit Shroom3 overexpression resulting from a single nucleotide polymorphism risk allele at intronic rs17319721. 22
In this study, we investigated Shroom3 heterozygous null mice at postnatal day 3, postnatal day 30, and postnatal day 90 to determine the functional and phenotypic consequences of reduced Shroom3 expression. We show that reduced Shroom3 expression causes altered apical-basolateral epithelial polarity in select adult nephron segments and sometimes results in hypoplastic unilateral kidneys. Altogether, these studies demonstrate that Shroom3 heterozygous null mice exhibit a mild kidney disease phenotype at 3 months that does not result in overt kidney damage or decreased kidney function.
Methods
Mouse Models
Shroom3 heterozygous mutant mice Shroom3Gt(ROSA)53Sor/J (or Shroom3Gt/+) were gifted from Dr. Thomas Drysdale at the University of Western Ontario and originally generated by Hildebrand and Soriano. 17 These mice contain a gene trap cassette, SAβgalCrepA, that contains an adenovirus splice acceptor (SA), a bifunctional gene encoding a fusion between β-galactosidase (β-gal), Cre recombinase, and an MC1 polyadenylation (pA) sequence. 23 This cassette is inserted between exons 3 and 4 of the Shroom3 gene, resulting in the elimination of the short isoform and truncation of the long isoform of Shroom3. 17 Due to the embryonic lethality of homozygous Shroom3 mutants, the line is maintained using Shroom3 gene trap mice that only carry one copy of the gene trap allele (Shroom3Gt/+).
Tissue Processing and Histology
Whole kidneys from postnatal day 3, 1 month, and 3-month-old Shroom3Gt/+ and CD1 wild type mice were resected and imaged using a Leica EZ4 dissecting microscope and imaged using LAS software. All kidneys were fixed in 4% paraformaldehyde at 4°C for 48 hours and paraffin embedded. Kidneys were sectioned to 5 µm on a Leica microtome, mounted on Superfrost plus microscope slides (VWR, Mississauga, Ontario), and incubated overnight at 37oC. Tissue sections were either stained with hematoxylin and eosin (H&E) (Sigma Aldrich, Oakville, Ontario) or utilized for immunofluorescence. All histology images were acquired using the Olympus BX80 light microscope and Cell-Sens image acquisition software. For immunofluorescence and immunohistochemistry, kidney sections were deparaffinized and re-hydrated using xylene and graded ethanol washes (100%, 95%, 70%, and 50%) then washed in phosphate buffered saline (PBS). Antigen retrieval was performed for 5 minutes in a pressure cooker with 11.4 mmol/L sodium citrate buffer solution (pH 6.0). For immunofluorescence, samples were blocked with 7.5% normal goat serum and 4.5% bovine serum albumin at room temperature for 1 hour, followed by incubation with primary antibodies
Blood Pressure Measurements (Tail-Cuff)
Blood pressures of conscious mice were measured using a CODA 6-Channel High Throughput Non-Invasive Blood Pressure system (CODA-HT6, Kent Scientific, Torrington, Connecticut) according to the manufacturer’s instructions. The apparatus applied the tail-cuff method to determine blood pressure. Mice were placed into holders that restrained movement; the head was secured with a nose cone while the tail extended out of the rear end of the holder. The holders were kept on a warming platform to maintain the mice at a minimum temperature of 32°C for the duration of the experiment. Two cuffs were placed around the tail: an occlusion cuff which impedes blood flow by inflation, and a Volume Pressure Recording (VPR) cuff which measures the swelling of the tail that occurs as a result of returning blood flow upon deflation of the occlusion cuff. The VPR sensor measures and outputs 6 different physiological readings: systolic and diastolic blood pressure, mean arterial pressure, heart rate, tail blood flow, and tail blood volume. To condition the mice and minimize the level of stress caused by the tail-cuff apparatus, mice were trained daily for 2 to 3 weeks prior to taking the basal blood pressure measurement. Blood pressures were monitored over 2 sets of 15 cycles each, with 5 additional acclimatization cycles at the start of the first set; measurements were taken at the same time each day.
Body Weight and Kidney Weight Measurements
Mice were weighed at postnatal day 3, 1 month, and 3 months, then euthanized by carbon dioxide. The kidneys were extracted, trimmed of excess tissue, and washed in ice-cold PBS. Kidneys were blotted dry and weighed on the same scale used to measure body weight. Kidney weights were only obtained at postnatal 1 month and 3 months as the postnatal day 3 kidneys were too small to obtain accurate weight measurements.
Urinalysis
Urine was collected on various days, and collection always occurred at 9:00 AM on each day. To induce urination, the mice were put on a cold glass plate where they were scruffed and pressure was directly applied to the bladder area. The urine was transferred to an Eppendorf tube and then stored at -80°C until analysis. Urine was analyzed for urinary protein and creatinine levels using ChemStrip 10 dip sticks (Roche, Pickering, Ontario) (Western University Hospital, London, Ontario). Urine was also analyzed for the presence of glucose using BioStrip reagent strips (Innovatek, Vancouver, British Columbia).
Statistical Analysis
Two-tailed t-tests were performed using GraphPad Prism (v8.1.2, San Diego, California) to compare levels in wild type and Shroom3Gt/+ mice, P-values of <.05 were considered statistically significant and data were reported as mean ± SEM.
Results
Results 1: Normal Shroom3 Protein Expression in the Tubular Epithelium
Our previous studies characterized the expression and function of Shroom3 in both embryonic and adult mouse kidneys. These studies revealed that Shroom3 is expressed in the glomeruli and is responsible for maintaining normal podocyte cytoarchitecture and function by modulating the cellular actomyosin network.
16
In this study, we built on these previous findings by expanding the expression pattern characterization of Shroom3 to the kidney tubular epithelium. Immunofluorescence in adult 3-month-old mice revealed apical Shroom3 expression in the cortical and medullary tubules, with higher levels of expression seen in the cortical tubules (

Shroom3 is expressed in the kidney tubular epithelium.
Results 2: Shroom3Gt/+ Mice Do Not Show Gross and Histological Abnormalities
We performed a series of experiments to establish an overall characterization of the Shroom3Gt/+mice. In comparing the Shroom3 expression pattern between wild type and Shroom3Gt/+mice, it was confirmed that the heterozygous null mice exhibited decreased Shroom3 expression levels by immunofluorescence. However, while there is reduced protein expression of Shroom3 in the Shroom3Gt/+mice, the expression pattern remains apically localized in both the wild type and Shroom3Gt/+mice (
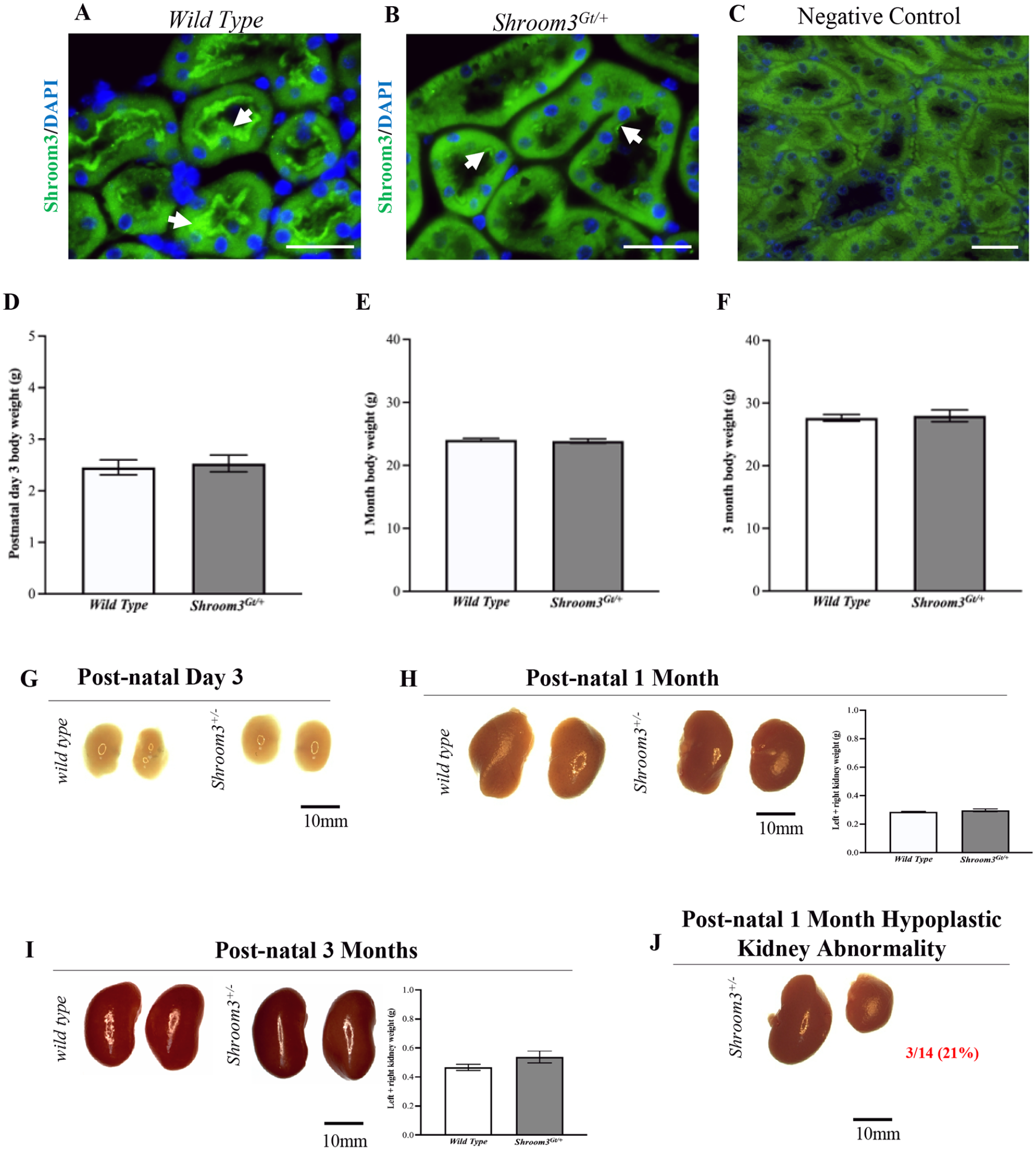
Heterozygous Shroom3 mutant kidney expression pattern. (

Shroom3Gt/+mice do not exhibit overt histological abnormalities. (
Results 3: Shroom3Gt/+ Mice Demonstrate Altered Proximal Tubule Apical-Basolateral Orientation
Our previous studies defined the effects of Shroom3 null mutant mice on embryonic kidney development. In comparison to wild type mice, the Shroom3 null mutant embryos demonstrated abnormal disrupted glomerular formation during earlier stages of kidney development (E13.5). The glomerular abnormalities observed at E13.5 were not obvious at E18.5, electron microscopy revealed abnormal podocyte morphology at later stages of kidney development (E18.5).
16
Additionally, these studies observed a dose-dependent reduction in glomerular number in both Shroom3 homozygous, and to a lesser extent, heterozygous null mutant embryos in comparison to wild type at E18.5. This demonstrates that abnormalities during early kidney development can lead to disrupted glomerular formation and reduced glomerular number in later stages of development.
16
In our present study, we extend these findings by investigating whether kidney developmental abnormalities in Shroom3Gt/+ embryos translate to abnormalities in the different nephron segments in postnatal and adult mice. Given that we did not observe any overt histological abnormalities in the glomeruli and renal tubular epithelia (
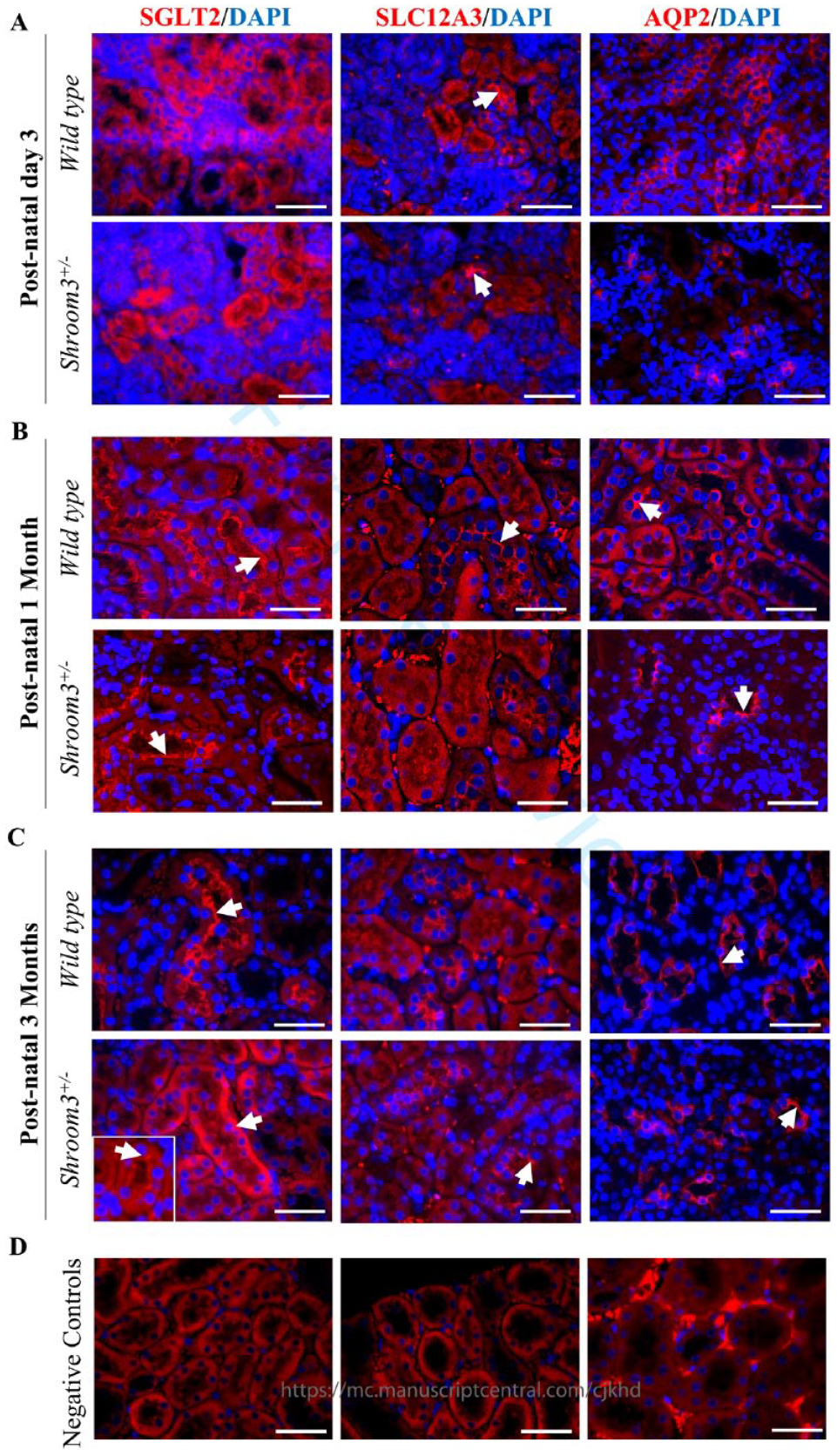
Shroom3Gt/+mice exhibit modest alterations in apical-basolateral orientation in 3-month-old mice. (
Results 4: Shroom3Gt/+ Mice Do Not Show Evidence of Proximal Tubular and Glomerular Injury
Previous studies have suggested that a loss of normal apical-basolateral orientation in renal epithelium can be associated with tubular injury.
24
Given the disrupted apical integrity and altered epithelial cell polarity seen in the renal tubular epithelium (
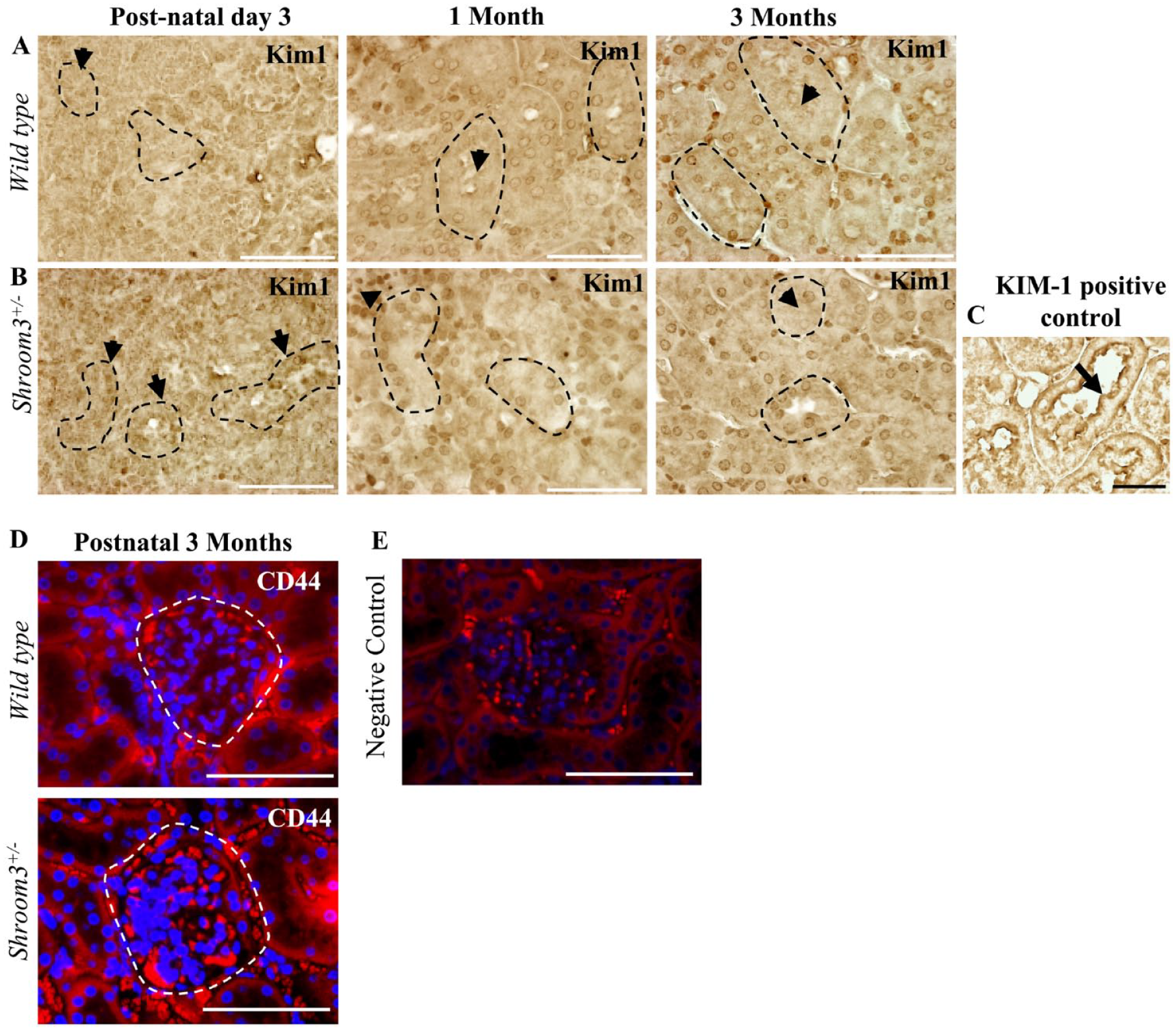
Shroom3Gt/+mice do not exhibit proximal tubular and glomerular injury. (
Results 5: Shroom3Gt/+ Mice Do Not Show Renal and Cardiovascular Physiological Abnormalities
The most prominent molecular-level tubular abnormalities occurred in the proximal convoluted tubules of 3-month-old Shroom3Gt/+mice. This is made evident by the observed altered localization of the proximal convoluted tubule marker, SGLT2, in the Shroom3Gt/+kidneys (
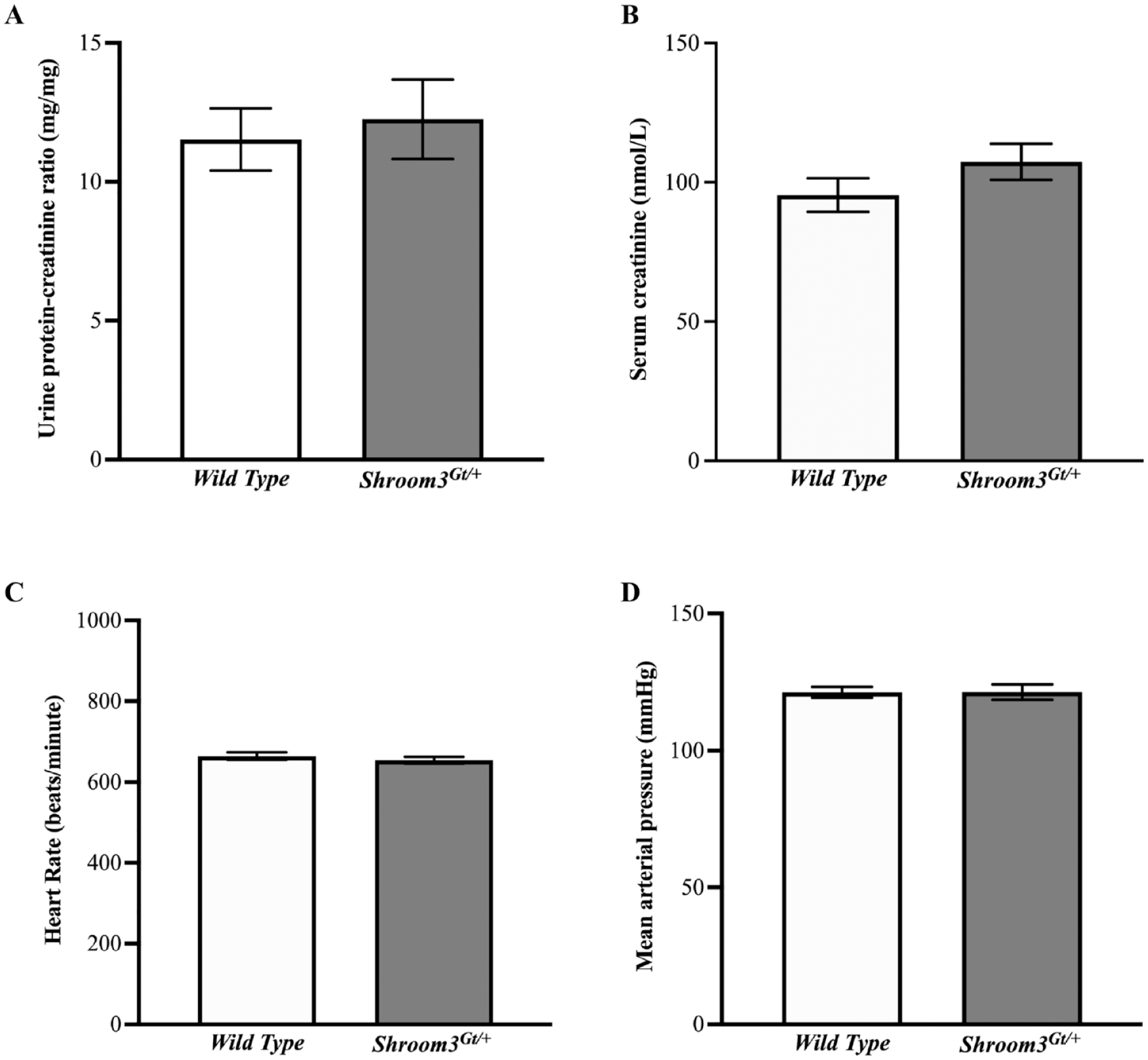
Shroom3Gt/+mice do not show renal or cardiovascular physiological abnormalities at postnatal 3 months. (
Discussion
The data presented in this study provides evidence that Shroom3Gt/+ mice exhibit a mild kidney phenotype at 3 months, primarily at the molecular level. Consistent with the Shroom3 expression pattern, we detected altered apical-basolateral orientation of proximal tubular cells at postnatal 3 months as demonstrated by the reduced expression and lack of apical SGLT2 in the Shroom3Gt/+ kidneys. This mild kidney abnormality was not associated with changes in somatic growth, gross anatomy and histology, tubular morphology and integrity, or other cellular and molecular mechanisms in the Shroom3Gt/+mutants. However, we did observe hypoplastic unilateral kidneys in postnatal day 3- and 1-month old mice. Taken together, our results show that Shroom3 is important in maintaining apical integrity in the renal tubular epithelium; however, its heterozygous deletion is not a major contributing factor to abnormal kidney structure and function.
Shroom3 Protein Expression in Adult Mice
Our previous studies characterized the expression pattern of Shroom3 in the developing kidney. Endogenous gene expression was examined through LacZ activity which reports the presence of Shroom3 expression. It was found that Shroom3 is strongly detected in the condensing mesenchyme located in the outermost part of the kidney cortex and in specific cells of the developing nephron structures including the developing and mature podocyte cell layer. Shroom3 was also detected in the ureteric bud stalks and tips of the developing collecting ducts. 16 Our results expanded on the expression pattern and demonstrated Shroom3 expression in the proximal and distal convoluted tubules as well as the medullary collecting ducts. In all cases, expression was localized primarily to the apical regions of the cell. This Shroom3 expression pattern can translate to important functional roles in maintaining proper cell shape, providing tissue rigidity, and regulating the subcellular localization of various important molecules such as F-actin, RhoA, Rho-Kinases, and non-muscle-Myosin II.6 -8,17 Additionally, Shroom3 may be responsible for apical cell-cell adhesion as it associates with the actin cytoskeleton in both adherens and tight junctions.7,11,31,32 Collectively, these studies demonstrate that the Shroom3 expression pattern is also detected in the postnatal period suggesting it has an important role in maintaining the tubular epithelium. This suggests a likely role for Shroom3 in cell communication, organization and regulation, and future studies could focus on uncovering the molecular significance of the Shroom3 expression pattern in the mature tubular epithelium cells of adult mice kidneys.
Reduced Shroom3 Expression May Impair Branching Morphogenesis and Nephrogenesis
During kidney development, the ureteric bud undergoes branching morphogenesis and signals the nephron progenitor cells to form the various segments of the nephron. Bifurcations of the ureteric bud repeatedly occur throughout embryonic development to produce thousands of collecting tubules and millions of nephrons in order to establish the renal architecture. 33 This highlights how interactions between the ureteric epithelium and nephron progenitor cells are required for kidney development. As previously described, Shroom3 demonstrated high expression levels in the early specialized mesenchyme progenitor cells with lower expression in the ureteric bud epithelium. 16 Therefore, Shroom3 may play a role in kidney development where reduced expression may, in some instances, impair nephrogenesis and branching morphogenesis leading to an anatomically smaller kidney. This may explain the rare observation of the hypoplastic unilateral kidney seen to some degree in postnatal day 3 and more evidently in postnatal 1 month old Shroom3Gt/+ mice. Yet this observation was only seen in 3/14 of the 1-month-old Shroom3Gt/+ mice and these smaller kidneys did not exhibit any marked histological abnormalities. However, based on the expression pattern and function of Shroom3, we hypothesize that Shroom3 regulates the ability of the nephron progenitor cell population to aggregate and therefore its reduced expression may disrupt reciprocal signaling events in certain instances causing disturbances in normal kidney morphology. Further investigation is required to provide a better understanding of these processes in affected hypoplastic kidneys.
Reduced Shroom3 Expression Alters Epithelial Polarity in Select Adult Nephron Segments
Shroom3 has an established role in generating the appropriate shape and function of epithelial tissues during embryonic development.8,16,34 It typically accomplishes cell shape changes and tissue morphogenesis through modulating the actin cytoskeleton at the apical surface of developing epithelial cells.7,9,12 Previous studies demonstrated this in the kidneys using embryonic Shroom3 null mice where loss of apically distributed actin resulted in glomerular abnormalities including podocyte effacement and reduced podocyte numbers. 16 Studies confirmed that reductions in Shroom3 expression postnatally causes defects in kidney function, such as albuminuria and podocyte foot process effacement.16,18 Our present study furthered the investigation on potential regulatory roles by looking at the relationship between Shroom3 and cell polarity in adult nephron segments. The SGLT2 immunofluorescence results demonstrated molecular abnormalities in the Shroom3Gt/+ kidneys that were primarily restricted to the proximal tubules; the nephron segment with the most prominent Shroom3 expression. Additionally, SLC12A3 and AQP2 did not demonstrate misexpression in the distal convoluted tubules nor the collecting ducts, respectively. Since recent evidence suggests that actin cytoskeleton dynamics may be involved in modulating ion channel localization, the disrupted apical integrity observed in Shroom3Gt/+ mice proximal tubules may be due to a lack of Shroom3 mediated actin organization and localization.35 -37 However, more studies are needed to confirm if Shroom3 plays a role in actin-dependent ion channel regulation in the nephron. Altogether, our findings in combination with others demonstrate that decreased Shroom3 levels in adult mice can lead to altered apical localization of key ion channels in the proximal tubule, possibly through a lack of Shroom3 mediated actin dynamics regulating proper ion channel localization.
Reduced Shroom3 Expression May Impact Tubular Repair and Drive Secondary Glomerular Injury
In our present study, we found no evidence of overt histological damage at any of the studied ages. However, we found mild disturbances in the apical integrity of proximal convoluted tubules demonstrated by the marked apical absence and reduced expression pattern of the tubule marker SGLT2 at 3 months. Kidney function appeared normal as there were no detectable differences in urinary protein/creatinine ratios and serum creatine levels in 3-month-old Shroom3Gt/+ mutants. Our study did not detect any overt glomerular pathologies in postnatal Shroom3Gt/+ mice at 3 months, despite previous evidence of glomerular abnormalities during embryonic kidney development and glomerulopathy at 1 year. 16 It is possible that the severe nephron alterations during development result in nephron death while the surviving nephrons exhibit changes at the molecular level. Therefore, the molecular changes found at 3 months may contribute to apical-basolateral disorientation which then prevents appropriate recovery from kidney injury. This concept was demonstrated by our recent publication that subjected Shroom3Gt/+mutants to ischemia reperfusion kidney injury. This study demonstrated that after acute kidney injury, Shroom3Gt/+mice were able to recover kidney function, yet exhibited worse kidney histopathology, specifically in the tubular epithelium. 21 These findings could suggest a model where proximal convoluted tubular damage could drive secondary damage in renal corpuscles or even amplify existing damage. A similar study investigated sublethal diphtheria toxin-induced renal epithelial injury in proximal tubule cells and found that after an acute non-recurring injury, the tubules demonstrated a complete recovery. However, after repeated injury, the tubules were unable to repair properly resulting in glomerulosclerosis, demonstrating that glomerular damage occurred as a secondary effect of repeated acute proximal tubular injury. All in all, this study showed that long term repeated injury surpasses the self-repair capacity in the proximal tubules leading to tubular damage and inflammation that may then drive postnatal glomerular damage. 38 Comparably, our present findings demonstrated that Shroom3 deficiency initially results in mild damage in the proximal tubules and does not immediately affect the glomeruli or other segments of the nephron. However, over the period of 1 year, the cellular damage in the proximal tubules progressively worsened, causing secondary damage to the renal corpuscles. 16 Our findings suggest that the observed mild molecular abnormalities seen in Shroom3 heterozygous null mice at 3 months may not immediately contribute to abnormal kidney structure and function, however abnormalities may manifest with age or in the event of external insults. Taken together, the abnormal molecular phenotype seen with reduced Shroom3 expression at 3 months may contribute to increased sensitivity to kidney disease and worsened tubular repair and glomerular disease after a kidney insult or with aging.
Conclusion
In conclusion, the heterozygous loss of Shroom3 in mice does not result in overt kidney damage or decreased kidney function. Rather, the only mild abnormality detected in the Shroom3Gt/+mutants was the altered apical orientation of proximal tubular cells which did not translate to tubular damage or decreased kidney function. These findings set the important groundwork establishing that Shroom3 genetic mutations alone do not cause kidney disease. Yet, there is a possibility that individuals with Shroom3 genetic anomalies may experience worsened kidney disease when in the presence of an external insult, such as acute kidney injury, an area that requires further exploration in future studies.
Footnotes
Acknowledgements
We thank Dr. Robert Gros for his analysis on blood pressure.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This study was supported by grants from the Kidney Foundation of Canada and NSERC and startup funds from McMaster University (to D.B.).
Ethical Considerations
This study was approved by the Animal Research Ethics Board at McMaster University (Animal Utilization Protocol #22-02-04) on April 2, 2022. All mice experiments were performed in accordance with Canadian Council for Animal Care and McMaster University institutional guidelines.