
Abstract
Lake sediments constitute natural archives of past environmental changes. Historically, research has focused mainly on generating regional climate records, but records of human impacts caused by land use and exploitation of freshwater resources are now attracting scientific and management interests. Long-term environmental records are useful to establish ecosystem reference conditions, enabling comparisons with current environments and potentially allowing future trajectories to be more tightly constrained. Here we review the timing and onset of human disturbance in and around inland water ecosystems as revealed through sedimentary archives from around the world. Palaeolimnology provides access to a wealth of information reflecting early human activities and their corresponding aquatic ecological shifts. First human impacts on aquatic systems and their watersheds are highly variable in time and space. Landscape disturbance often constitutes the first anthropogenic signal in palaeolimnological records. While the effects of humans at the landscape level are relatively easily demonstrated, the earliest signals of human-induced changes in the structure and functioning of aquatic ecosystems need very careful investigation using multiple proxies. Additional studies will improve our understanding of linkages between human settlements, their exploitation of land and water resources, and the downstream effects on continental waters.
Introduction
Lake systems are fundamentally important to the environment, biosphere, and human populations, but are under increasing stress from anthropogenic impacts, and vulnerable to Earth’s changing climate. Understanding how freshwater ecosystems change through space and time is crucial to ensuring global-scale resource sustainability at a time when humans increasingly drive environmental change. The timing and onset of human activities and how these have modified aquatic ecosystems in the past are becoming of increasing scientific and management concern (Dearing et al., 2006; Gell et al., 2007b; Saulnier-Talbot, 2016), especially as global aquatic ecosystems shift towards alternative Anthropocene states (Kopf et al., 2015). Palaeolimnological analysis, the study of the physical, chemical and biological information preserved in lake and wetland sediments, provides a valuable approach to reconstruct natural and anthropogenic changes for periods pre-dating instrumental observation. Long-term records of environmental change are of particular interest to establish the range of natural variability (i.e. ‘background’ or ‘reference’ conditions of an ecosystem in the absence of extensive human impact), from which the timing and magnitude of anthropogenic disturbance may be quantified. The identification of this discernible change allows the setting of management targets (Gell et al., 2013), and aids scientific understanding of the complex interactions between people, climate and the environment. Such data are crucial for modelling future scenarios relating to water quality and availability. Furthermore, knowing the timing of first impact provides data on the response and/or recovery of an aquatic system once the ‘pressure’ (e.g. point source pollution) has been removed, and insight into the legacy and cumulative effects of early human impacts on contemporary (and future) aquatic systems.
People have affected natural environmental systems for millennia at local scales by disturbing vegetation cover (Edwards and Whittington, 2001), but sustained land-use changes began with the rise of Neolithic farming: the so-called ‘Neolithic revolution’. According to Ruddiman (2005), the global effects of early farming on the atmosphere are detectable some 8000 years ago when humans began to influence atmospheric CO2 levels, although the extent of human impact on the climate at this time remains controversial (Lewis and Maslin, 2015). Nonetheless, direct evidence of early human impacts on landscapes at a local scale is well known through an extensive global array of archaeological investigations at local and regional scales (e.g. Whittle and Bickle, 2014).
Today, the direct (e.g. through species introductions, water abstraction) and indirect (e.g. through altering lake catchments) impact of people on lake systems is unquestionable (Hering et al., 2015; Leavitt et al., 2009). The effects of early human activity on aquatic ecosystems, typically driven by soil erosion, habitat modification or pollution, have been demonstrated in many case studies. These studies show that human pressures on aquatic systems can take many forms and are highly variable in time and space, but the timing and magnitude of first detectable impacts at regional and continental scales remain ill-defined. In order to contextualise the pace, direction and magnitude of recent aquatic transitions, and to consider how reversible they may be, this paper provides a global synthesis of the earliest drivers and responses of these vital ecosystems.
While lake sediment records can provide local and regional evidence about environmental changes, detecting human influence throughout history can be challenging because of the subtlety, complexity and variable nature of many early human impacts on the environment (see Mills et al., 2017). The nature of disturbance, preservation of the impact signal and, not least, the provision of a reliable chronology all affect our ability to detect changes. Moreover, palaeolimnological proxies are frequently used to indicate early human terrestrial disturbances in a region (e.g. pollen assemblages, charcoal and sediment influx to infer deforestation in a catchment) but these are not necessarily accompanied by evidence of a concurrent aquatic transition. By ‘aquatic transition’, we refer to an episode during which an aquatic ecosystem experiences a change in the way it is physico-chemically or biologically structured or is affected in its functions, such as in passing from one trophic state to another. Available research seemingly suggests that the first dectectable human impacts on the environment rarely lead to aquatic transitions, although future analysis focusing more on aquatic variables may challenge this view.
Herein, we review evidence of early (pre-1850
Lake sensitivity to human impact
Early anthropogenic impact on aquatic systems is highly variable and can depend on geographical location and duration of inhabitation. Figure 1 synthesizes the timing of arrival of anatomically modern humans in various regions, migration routes, and global sea-level fluctuations (since lower sea-level during ice ages facilitated passage between land masses). A more detailed description is provided in the Supplementary Material (available online). However, the first occurrence of humans in the archaeological/palaeolimnological record does not necessarily correspond to a shift, or transition, in aquatic ecosystems (Armesto et al., 2010; Bush et al., 2016; Dodson, 1979). In fact, the first ‘detectable’ human impact within a lake catchment may not force a response in the aquatic system, possibly because of the negligible degree of impact caused by smaller population sizes, their culture, and less ‘sophisticated’ technologies.
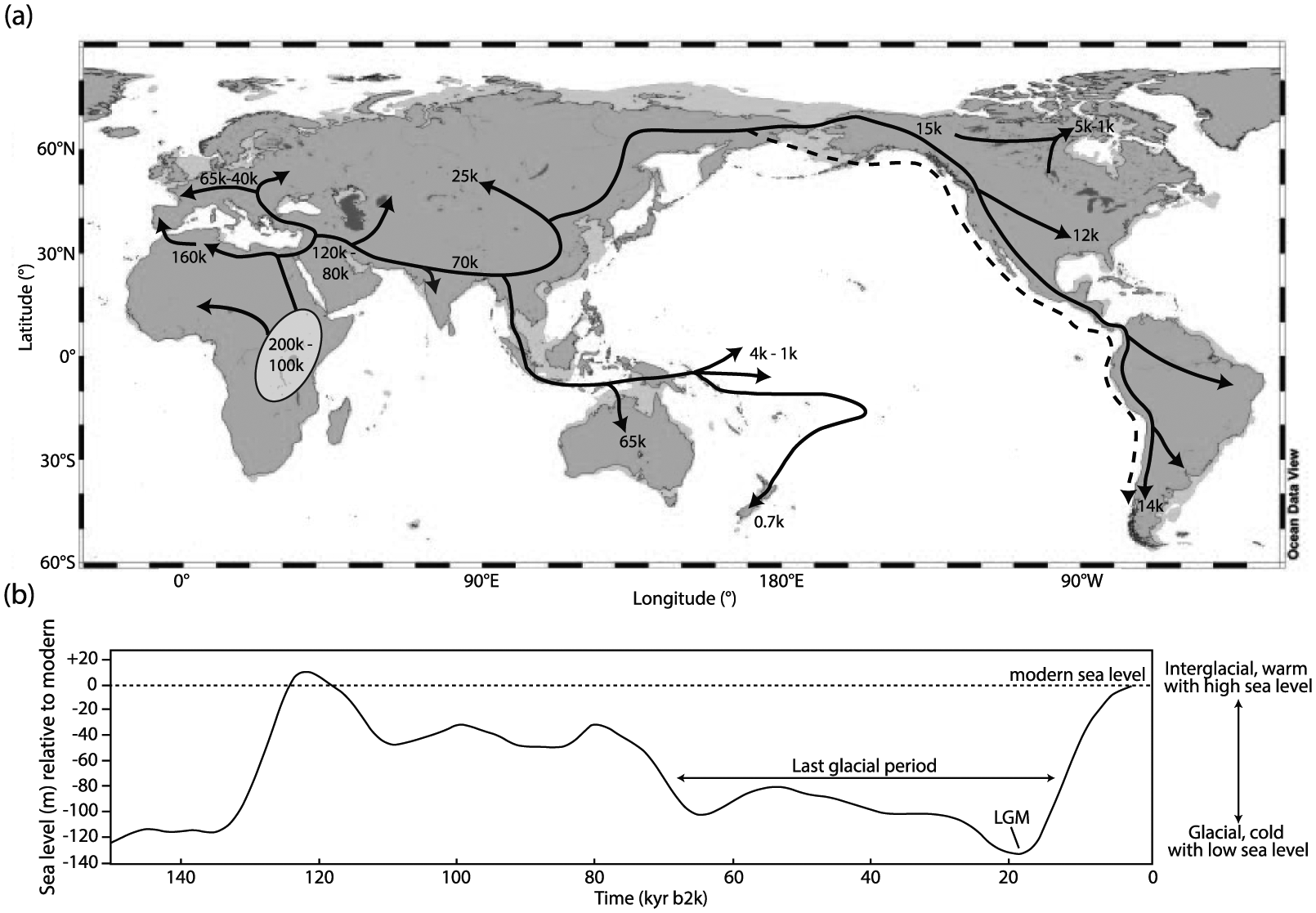
(a) Map showing temporal and directional markers (arrows) of early human migrations across continents (k = 1000 years before year 2000), based on the literature cited in the text. Dashed line indicates a possible coastal migration route. Oval indicates origin of anatomically modern humans (AMH) in eastern Africa. Pale grey indicates emerging land contours at lower sea levels (−100 m), facilitating access to land masses. (b) Sea level change over the last 150 kyr (based on Waelbroeck et al., 2002).
With a lower magnitude of stress, some lake ecosystems may have the capacity to adapt rather than shift into an alternative state. This may also be true where technology has evolved gradually through time, for example in Europe (Bradshaw et al., 2005b) and China (Shen et al., 2006). In these regions, lake systems adapted to slow changes until c. 6000 b2k, when catchment modification and the intensification of agriculture increased to such an extent that a distinct transition is observed in the function of aquatic ecosystems. In other parts of the world, where advanced technology and intensive agricultural practices were introduced more recently (e.g. within the last two centuries in Australia and New Zealand), the effects on ecosystems are far more acute, though this could also reflect the sensitivity of the systems (Gell and Reid, 2014, 2016; Gell et al., 2005b; Kattel et al., 2014). Lakes in remote regions have only been subjected to anthropogenic nutrient loading via atmospheric deposition as a result of transboundary pollution during the last few decades (Bergstrom and Jansson, 2006; Bergstrom et al., 2005; Brahney et al., 2015; Wolfe et al., 2001; Yang et al., 2010b).
The sensitivity of lakes to any given driver not only depends on the intensity of the perturbation, but also on how the lake system processes the environmental perturbation; some lakes may exhibit an immediate response to a particular disturbance, whereas others may appear unresponsive to the same change or pressure (see Scheffer et al., 2012). For example, large and deep lakes show higher buffering capacity, expressed by a delayed response, against nutrient input from the catchment (Dearing and Jones, 2003), whereas smaller lakes may show very little lag between the change in land use and the lake response (e.g. Heinsalu and Veski, 2007).
Detectable human impact relative to climate
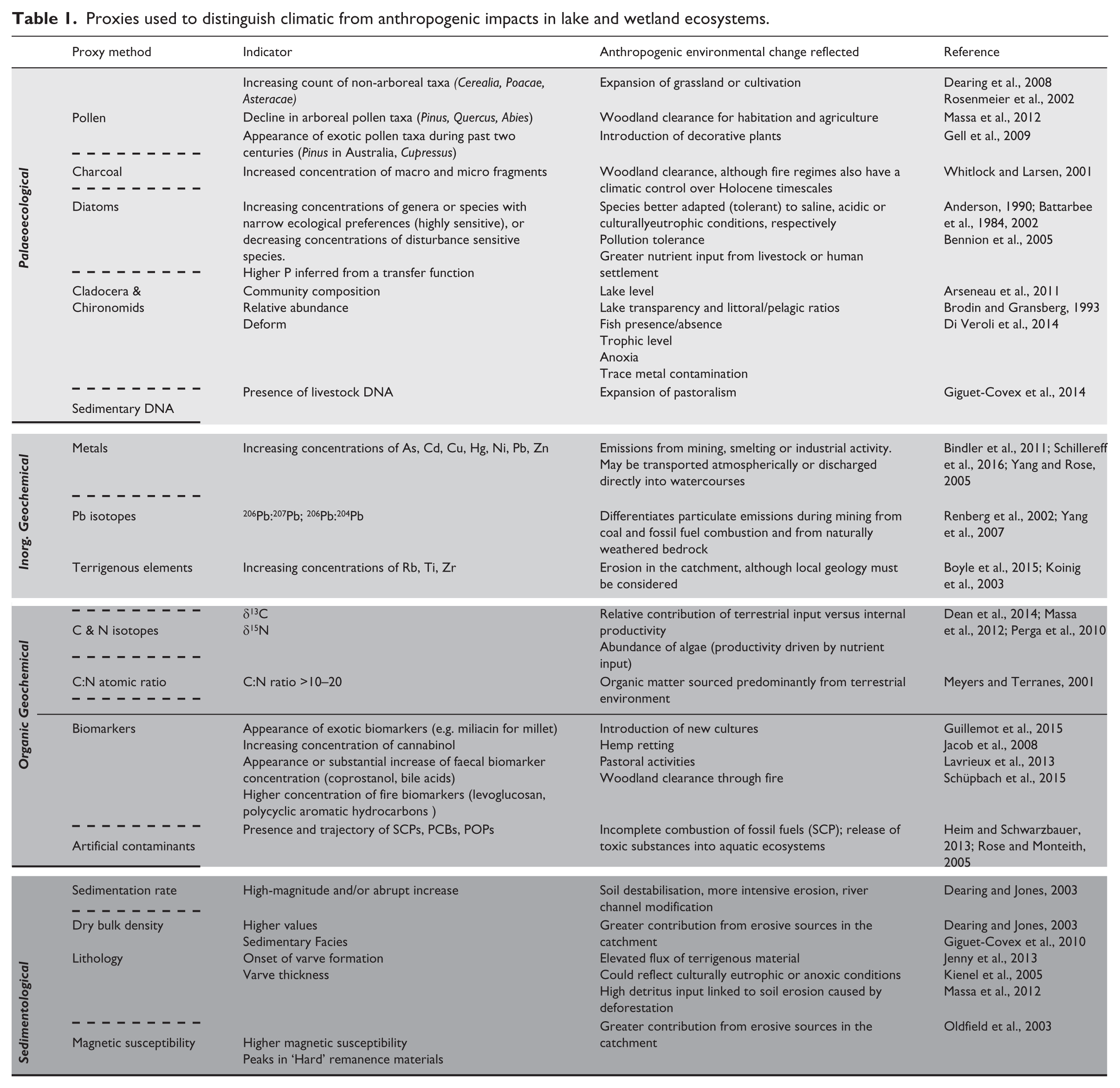
A major challenge in identifying human impact on the functioning of aquatic ecosystems is the ability to confidently separate aquatic responses driven by people from those that can be attributed to natural climate change (Battarbee and Bennion, 2011; Mills et al., 2014, 2017). Closely coupled with this is the need to understand potential leads and lags within the system, hysteresis, loss of resilience, and the sensitivity of lake sites to a given perturbation (Magnuson et al., 2004), and whether lakes have crossed a tipping point (Langdon et al., 2016). For example, lakes in the English Lake District (UK) have been shown to become increasingly sensitive to climate following recent eutrophication (Dong et al., 2012a, 2012b; Moorhouse et al., 2014). A number of methodological approaches can be employed to tackle the question of what impacts or disturbances can be attributed to climate and what can be attributed to people (Mills et al., 2017). These range from site selection using a paired lake approach, such as non-impacted versus impacted sites (Moorhouse et al., 2014), to spatial scale (e.g. landscape-scale analyses; Keatley et al., 2008) and the careful selection of proxy indicators (Table 1). When undertaking studies to disentangle multiple drivers on ecosystems, the use of a multi-proxy approach is invaluable, especially when drivers operate at different frequencies. Using multiple lines of evidence can help overcome ambiguity and ratify the identification of the vectors of change in a system (Table 1; Mills et al., 2017; Perga et al., 2015). Furthermore, a number of advanced statistical methods (e.g. multivariate regressions, Bayesian networks) allow apportionment of climate versus anthropogenic impacts on lake systems (Birks et al., 2012; Simpson and Anderson, 2009; Simpson and Hall, 2012).
Proxies used to distinguish climatic from anthropogenic impacts in lake and wetland ecosystems.
Anthropogenic drivers of catchment and lake changes
Vegetation clearance is often the first evidence of human activity recorded in sediments (typically observed through changes in the pollen and charcoal record), but this change in the catchment does not necessarily trigger an abrupt response in the aquatic ecosystem itself, at least not initially (Armesto et al., 2010; Barr et al., 2013), and may vary with the degree of resistance of the lake ecosystem. Similarly, pollution, which often results from human activities outside the lake, can be relatively easily detected in the sedimentary record, but its ecological impacts are often variable. Conversely, some early human activities directly affected lakes and their ecosystems, such as modification of the aquatic habitat (for instance changes in lake level) or species introductions. Lakes are sensitive to various degrees to elements that modify their physical and chemical structure. For example, increased input of allochthonous matter tends to result in decreased water transparency, which can in turn affect the depth and the frequency of mixing in the water column which regulate nutrient distribution impacting the lake’s biota.
Here we consider any non-natural change in the flows of energy and material between a lake and its surroundings (both atmospheric and terrestrial) as a potential driver of ecological impact. Key factors are split into two subgroups: first, the anthropogenic drivers which result from human activities in the catchment or beyond (atmospheric transport), including (1) soil erosion and (2) pollution; second, the anthropogenic drivers of changes in the lake itself; (3) habitat modification and (4) species introductions (see Figure 2 for a schematic illustration of these drivers and the associated lake responses).
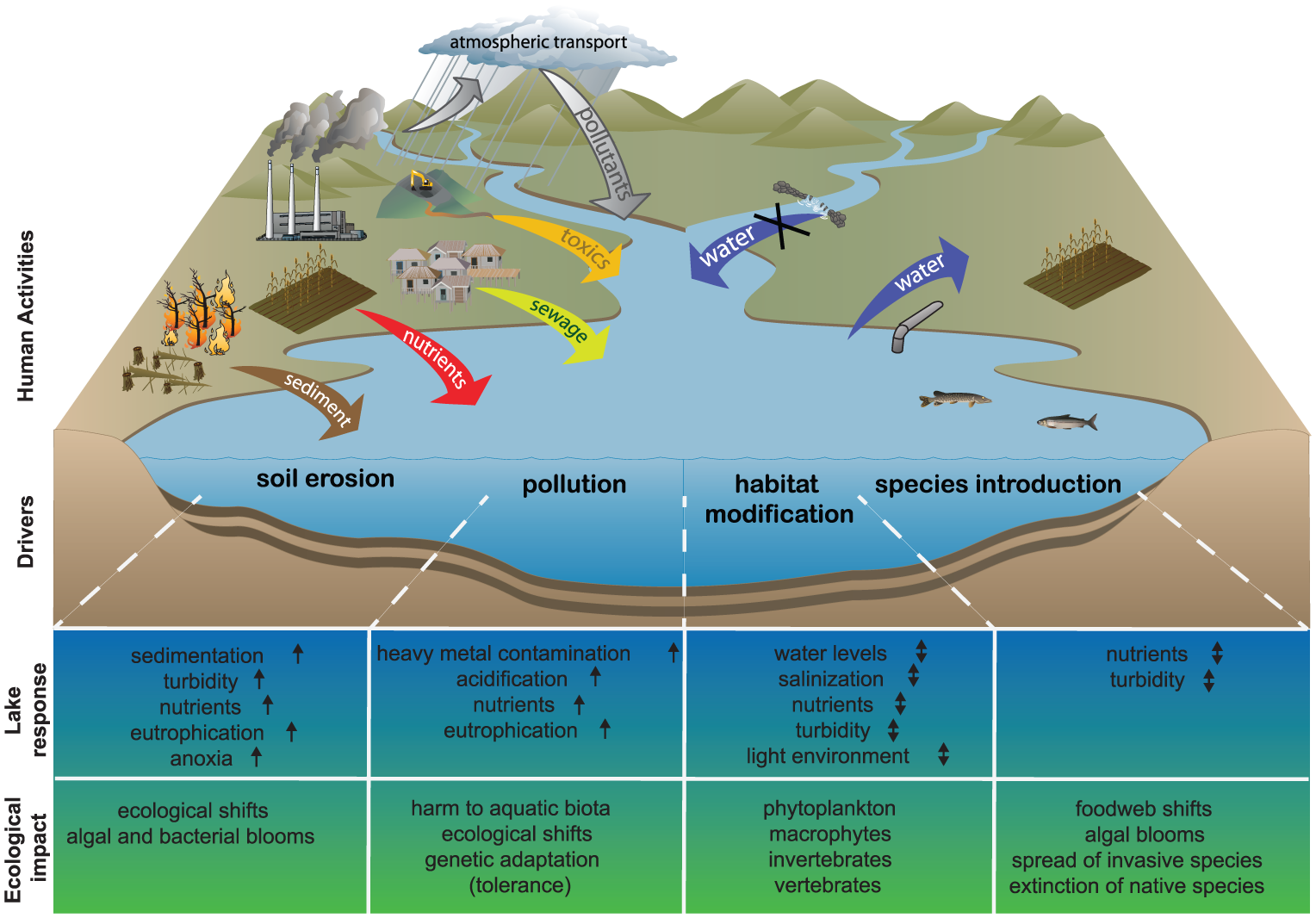
Diagram illustrating the four main anthropogenic drivers (upper drawing) and their associated lake-ecological responses (lower table). The drivers on the left (soil erosion, pollution) reflect human activities occurring in the catchment, whereas the drivers on the right are human activities affecting directly the aquatic system.
Soil erosion
Erosion is a natural process controlled globally by climate and tectonic cycles (Peizhen et al., 2001). Anthropogenic modification of catchments, including vegetation clearance, burning, agricultural and urban expansion leads to rapid fluctuations in soil erosion rates. Increased soil erosion impacts freshwater ecosystems through the enhanced external loading of sediment, nutrients and contaminants and can result in eutrophication, turbidity and/or flooding (Boardman, 2013). These fluctuations are often recorded in lake archives as variations in sediment accumulation rates (SAR; Dearing and Jones, 2003) or geochemical profiles (Boyle, 2001; Kylander et al., 2013) but concurrent ecological shifts have not been fully explored in many of these studies. Over the course of the Holocene, SAR in temperate and high latitude regions generally show a gradual decline as soils began to stabilise over the postglacial period as a result of increasing vegetation cover (Dearing, 1991). Abrupt and large perturbations to this pattern are often attributed to anthropogenic triggers whereas gradual changes in accumulation rates are attributed to climate forcing (Mills et al., 2017).
Basin morphometry and geology play key roles in determining the timing of initial recording of a perturbation linked to soil erosion. Factors such as lake:catchment ratio (Dearing and Jones, 2003), lake depth (Chiverrell, 2006; Dearing, 1991; Milan et al., 2015) and the geology and erosive potential of the catchment (Koinig et al., 2003) are involved. Few lakes studied to date contain an erosion signal linked to the rise of agriculture (12,000–4000 b2k, depending on the region). Those that do are primarily small, upland basins in temperate Europe (Edwards and Whittington, 2001). Later signals are more widely preserved, relating to more intensive land use during the Bronze (5000–2500 b2k) or Iron Ages (3200–1200 b2k), population expansion and agricultural intensification (Roman occupation, Mediaeval period), as a result of colonial activity (America and Oceania) or widespread impacts associated with industrialisation (Enters et al., 2010; Humane et al., 2016). Large lakes and drainage basins (>103 km2) tend to mask even a 20th-century acceleration in accumulation rate (Dearing and Jones, 2003), however this is not true for large oligotrophic lakes.
Pollution
Palaeolimnological evidence for pre-industrial contamination over millennial timescales is widespread, but careful analysis is required to distinguish early low-level anthropogenic effects from natural variation (Lindeberg et al., 2006). Earliest available records of human pressure from pollutants are generally from direct inputs at the site, while in remote regions, such as the Arctic or mountain regions, the earliest evidence is more recent (post-1850) and derives from atmospheric deposition (Wolfe et al., 2013).
Lake sediments provide evidence that numerous heavy metals have been in use for millennia around the globe. High levels of copper (Cu), lead (Pb) or mercury (Hg) resulting from smelting were observed in 3000–4000 year-old lake sediments from across Europe, China and Peru (see corresponding regional sections below). While pre-industrial contamination was widespread, contaminant concentrations in lake sediments were generally low compared with those related to industrial emissions and concentrations considered likely to cause harm to aquatic biota (i.e. Probable Effect Concentrations – PECs) (Macdonald et al., 2000). For instance, metallic pollution is known to affect diatom assemblages and morphologies (e.g. Hamilton et al., 2015). One notable exception is the Pb concentration in Laguna Roya (NW Iberia, close to Las Médulas, the largest Roman gold mine), where the concentration during Iberian-Roman times (c. 2300 b2k to 120
Nutrients can be considered as non-toxic pollutants, leading in the worst cases to eutrophication of aquatic ecosystems. Soil erosion (see section ‘Soil erosion’) is only one of the processes through which humans can increase nutrient delivery. Fibre processing operations, such as hemp retting, allow nutrients to be leached out directly into the water. Further, the use of phosphorus (P) in detergents and agricultural fertilisers led to a dramatic worldwide increase in P inputs into lakes during the 20th century (Jenny et al., 2016a).
Habitat modification
Under the term ‘habitat modification’ we include any human activity that has directly affected the physical flow of water into the system. This includes changes in river flows such as river engineering, groundwater manipulation, and water level management. The creation of new aquatic systems through damming could also be considered as an habitat modification, however we exclude artificial water bodies from this review (but see Saulnier-Talbot and Lavoie, in review).
The dynamics of a lake and how this is reflected in its sediment record tend to be influenced by the nature of the local system (e.g. basin morphometry and presence of floating or submerged vegetation), including the situation of a lake within a catchment (throughflow lake, floodplain lake, exo- or endorheic), and climate. For example, all of these factors affect variability in water levels and surface area, which in turn influence the availability of aquatic habitats, such as deep water or littoral environments, and simple or complex substrates, to all levels of the biota. The construction of impoundments raises water levels and rapidly increases the availability of deeper, pelagic habitats (e.g. Reeves et al., 2016) while resulting soil inundation can lead to increased mercury methylation and hence bioavailability. Intensive water extraction or diversion of inflows can lower water levels, increasing the proportion of shallow, littoral zones, as well as affecting the concentration of elements such as metals or salts in the water (e.g. Verschuren, 2001). Aquatic plant habitats may be impacted by the underwater light environment (e.g. inorganic or biogenic turbidity; Reid et al., 2007) and plant loss may also be caused directly by the release of toxicants (e.g. Sayer et al., 2006). Such changes may lead to a state shift in shallow lakes (sensu Scheffer et al., 1993) whereby elevated nutrients and turbidity provide an advantage to phytoplankton over macrophytes, which are shaded by suspended fine sediment and high phytoplankton biomass. This new state is then reinforced by further sediment entrainment and nutrient release from the sediments (Kattel et al., 2017). Furthermore, hydrological change can influence flushing and the onset of stratification, which may affect the lake nutrient budget (Tolotti et al., 2010) or exacerbate lake water anoxia causing plant loss (Dick et al., 2012). The loss of plant habitat likely results in a simplification of aquatic habitat and leads to a reduction in the diversity of dependent invertebrates (e.g. Davidson et al., 2013; Kattel et al., 2014) and vertebrate fauna.
Species introductions
Freshwater ecosystems are considered to be particularly susceptible to biological invasions because of relatively high levels of isolation and endemism (Vander Zanden and Olden, 2008). Thus, the introduction of non-native species into lakes and wetlands can induce important changes to their ecosystem structure and functioning (e.g. Gurevitch and Padilla, 2004; Kamenova et al., 2017; Pimentel et al., 2005; Schindler et al., 2001). Given the long history of biological invasions and their substantial effects, there is clear potential that biological invasions were the result of significant early human impacts on freshwater ecosystems, especially in the Americas, Australia and on oceanic islands.
Not surprisingly, most examples of biological invasions in palaeoecological records come from those groups of indicators that readily preserve in taxonomically definable forms, such as diatoms (Brunel, 1956; Reavie et al., 1998; Stoermer et al., 1985) and cladocera (Duffy et al., 2000; Hairston et al., 1999; Hall and Yan, 1997). Given the ability of diatoms to disperse, it is still controversial whether or not they are truly non-native taxa, but the application of molecular work on these and other organisms might help to resolve these questions (e.g. Novis et al., 2017).
In some cases, invasions by less well-preserved taxa can be inferred by tracking the patterns of change in other taxa. For example, the introduction of fish in formerly fishless lakes can be tracked based on the abundance, size and morphology of the invertebrate predator Chaoborus and its prey Bosmina (Labaj et al., 2013). Determining the degree to which biological invasions drive ecosystem change, or are a response to changes driven by other factors such as catchment disturbance, eutrophication or pollution, is an important question for biological invasion science, and one that palaeoecology, with its capacity to extend temporal perspectives, is in a strong position to answer (Kamenova et al., 2017; Willis and Birks, 2006).
One of the key issues that previously had limited the capacity of palaeoecology to track biological invasions and identify examples where invasions have driven significant early human impacts on lake and wetland ecosystems, is the small number of biological groups that can reliably be identified to species levels from preserved remains. In addition, the ability of small organisms (such diatoms) to disperse, makes the concept of non-native taxa rather controversial. However, recent developments in the analysis of ancient DNA preserved in lake sediments (see Anderson-Carpenter et al., 2011; Domaizon et al., 2017; Giguet-Covex et al., 2014) may make it possible to trace the timing and extent of species introductions with better accuracy, or to assess if certain species are in fact invasive or native (Preston et al., 2004; Stager et al., 2015). This powerful new tool in the palaeolimnological toolbox provides the possibility of tracing a greater range of species introductions even further into the past and to link them with early human presence.
Regional variation in first detectable human impact in and around aquatic ecosystems
Human civilizations colonised different regions at different periods of their technological development. This had a considerable impact on the timing, magnitude and nature of environmental change that they induced, on the landscape in general, and in lakes and wetlands in particular. Figure 3 shows an overview of the timing and nature of the first human impacts detected to date on lakes and tropical wetlands around the world. The first detectable human impacts are most often linked to catchment deforestation and soil erosion effects. Hydrological disturbance and metal contamination followed, leading to an array of new impacts.
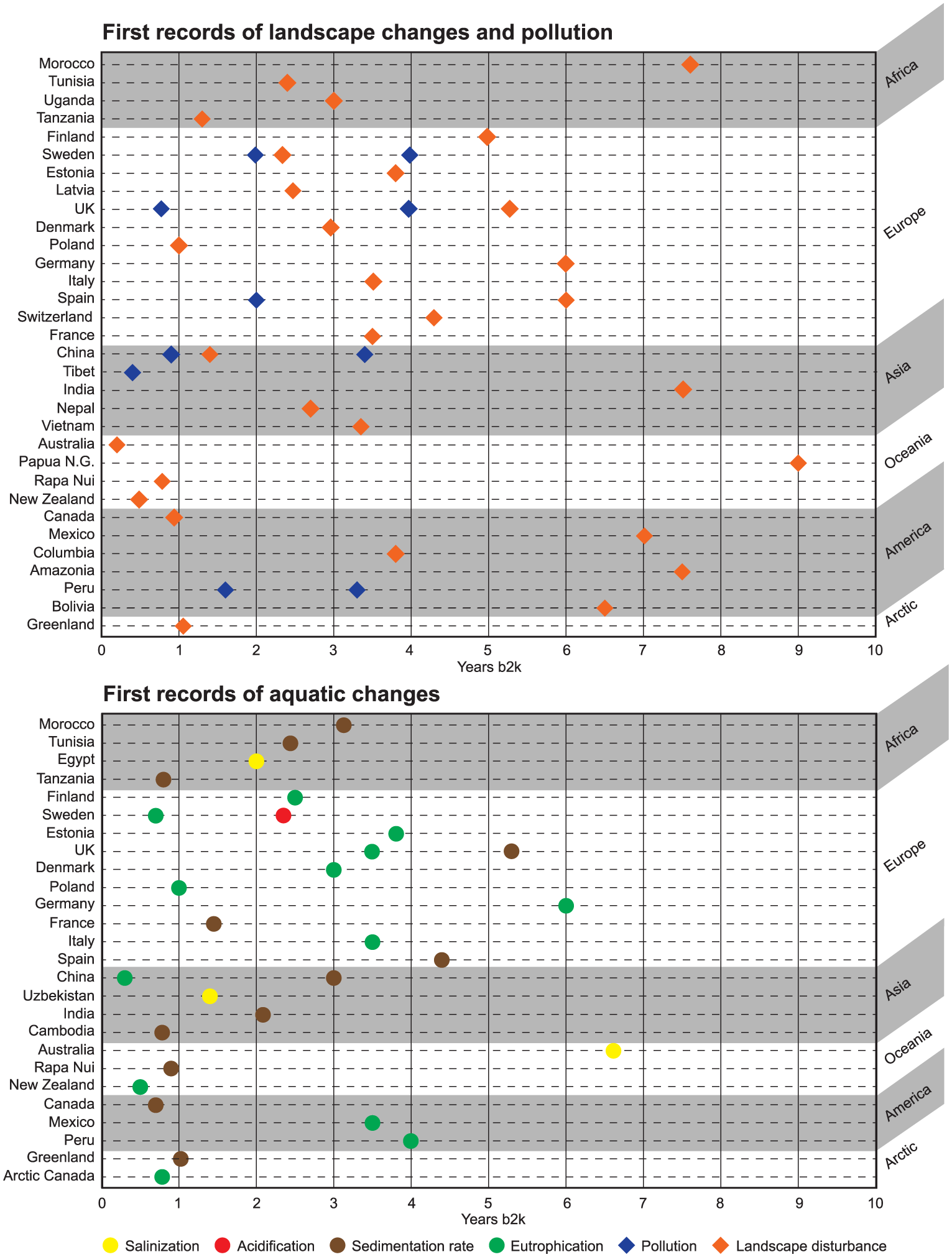
Top: Timing of the first anthropogenic landscape disturbance (orange) and pollution (blue) recorded in palaeolimnological archives around the world (b2k = time based on years before year 2000). Bottom: First records of aquatic changes worldwide. Salinization/hydrological change (yellow), acidification (red), sedimentation rate (brown), eutrophication (green). Only the first signal for each country (region) and each impact is shown.
Africa
Sub-Saharan Africa
The African continent has the longest history of human occupation, yet few studies explicitly address the impacts of people on African lake sites (Gelorini and Verschuren, 2013). Historically, much of the research on African lakes has focused on the large rift lakes and the long-term climatic records contained within (e.g. Finney and Johnson, 1991; Lærdal et al., 2002; O’Reilly et al., 2003; Olaka et al., 2010; Verburg et al., 2003). However, the sedimentary archives of the East African soda lakes (such as Naivasha, in Kenya; Verschuren et al., 2000, and Manyara in Tanzania; Casanova and Hillaire-Marcel, 1992) and the western Ugandan crater lakes (e.g. Mills et al., 2014; Ryves et al., 2011; Saulnier-Talbot et al., 2014; Ssemmanda et al., 2005; Verschuren and Russell, 2009) have also received considerable attention in recent years, mainly directed at inferring past climate or vegetation changes over various timescales. The same is true for the limited number of palaeoenvironmental studies of Western African lakes, except for Lake Bosumtui (Ghana; e.g. Russell et al., 2003; Shanahan et al., 2008; Talbot and Johannessen, 1992), which has been extensively used as a source of past climatic data for the region. The restricted number of lakes in Southern Africa is likely a factor for the low number of palaeolimnological studies conducted in this region.
Ancient human impact (pre-dating the Common Era) on aquatic systems is not obvious in Africa, despite the known presence of people. Given that many African regions experience very strong climatic variability and that industrialization has developed slowly and gradually on this continent, the evidence of human impact in lake sediments may be blurred. Evidence from the pollen record suggests early human disturbance c. 3000 b2k (related to Bronze Age iron ore smelting; Hamilton et al., 1986) – yet there is a lack of compelling evidence of synchronous shifts in aquatic indicators reflecting impacts on lake water quality and/or ecosystem function (or at least not as a signal that is distinguishable from the dominant climate system). In tropical Africa, it appears that the ‘first impacts’ that are detectable in the aquatic record are concurrent with colonial and/or post-colonial societal changes, even though we know modern humans have inhabited many parts of this region for thousands of years. However, this may be revised once palaeolimnological records become more abundant.
Some of the earliest evidence of aquatic change in tropical Africa comes from the swamp sediments in the North Pare Mountains in northeastern Tanzania. Analyses conducted by Heckmann et al. (2014) provided evidence for strong anthropogenic landscape transformations beginning in the 7th century
North Africa
There are several notable issues that have made the study of North African lake systems difficult for detecting early human impacts. In particular, sediment records in North African lakes are often incomplete, as a seasonally hot dry climate, shallow brackish water and high pH combined with rapid sediment accumulation are inimical to the preservation of some sediment microfossils (Flower et al., 2006). Nonetheless, there are numerous ephemeral lakes (sebkhas), as well as several coastal wetland lakes and lagoons, with sediments suitable for palaeolimnological analyses (e.g. Marquer et al., 2008; Soulié-Märsche, 2008).
Humans have been present in North Africa since at least the middle Pleistocene (780,000–125,000 b2k) and climate variation is closely linked to cultural development (Hassan, 1988). The first detectable impacts on lake sediment records occurred after the early-Neolithic when farming practices diffused from the Iberian Peninsula and the Near East around 7000 b2k (Linseele et al., 2016; Tassie, 2014).
Considerable palaeolimnological evidence from the Maghreb region indicates that human impacts on lake systems were slight until at least the middle Holocene. Human activities were first detected by vegetation indicators (including wetland plants) at several lowland sites in Morocco around 6000 to 7600 b2k (Reille, 1979; Zapata et al., 2013). Major vegetation disturbances began several thousand years later when farming, forest clearance and soil disturbance began under the Phoenician-Roman occupations of North Africa c. 2100 b2k. Pollen records from Atlas Mountain Lakes, Tunisian Lakes and sebkhas indicate anthropogenic forest disturbance and evidence for cultivation (Cerealia-type pollen) at that time (Cheddadi et al., 2015; Marquer et al., 2008; Mikesell, 1960; Reille, 1976; Stevenson et al., 1993). Early cultural interventions likely had only marginal or faltering effects on the lake ecosystems but multi-proxy studies are scarce.
Land-use changes promoted soil erosion, as evidenced in a number of North African lakes. An early soil erosion signal was reported in two Middle Atlas lakes beginning about 3000 b2k, possibly as pastoralism increased (Lamb et al., 1991). Alluviation, around what is now the lagoon of Ghar El Melh (Tunisia), occurred rapidly between 2400 and 1500 b2k, partly coinciding with the Punic-Roman period (Delile et al., 2015). Baioumy et al. (2010) attributed lithological variations in a Lake Qarun sediment core to lake level lowering by people regulating the lake inflow around the beginning of the Common Era and Keatings et al. (2010) showed that the ostracod species changed around this time.
Biological remains of aquatic macrophytes, diatoms and micro-invertebrates in North African aquatic ecosystems have provided insights into regional salinity variation. However, clear evidence for major anthropogenic impacts are only found for the 20th century in Tunisia, Morocco and Egypt, and are linked to large-scale hydrological modifications for irrigation and agricultural development (Barker et al., 1994; Birks et al., 2001; Stevenson et al., 1993). Foraminiferan records from the inland Egyptian Lake Qarun began in the 16th century
Europe
Holocene sediment records from European lakes provide evidence of eutrophication resulting from human activity dating as far back as the Neolithic period, >5000 b2k. Lakes in catchments with fertile soils have the longest history of human impact, dating from the start of primitive agriculture (following forest clearance), which is typically reflected by a decrease in tree pollen relative to herb and crop taxa (Birks et al., 1988; Räsänen et al., 2006; Ruddiman, 2003).
In a number of agriculturally rich lowland regions of Europe, the apparent pressure of nutrient enrichment on lakes extends back to the Bronze Age (c. 4000–2500 b2k; e.g. Bradshaw et al., 2006; Dressler et al., 2011; Fritz, 1989). A multi-proxy study of Dallund Sø, Denmark (Bradshaw et al., 2005b) provides one of the best examples of early enrichment during the forest clearances of the Late-Bronze Age (3000–2500 b2k) and the expansion of arable agriculture during the Iron Age (500
Across numerous European sites, there is a close correspondence in the timing of vegetation shifts associated with the onset of agriculture and aquatic indicators. For example, in northern Germany, lake trophic indicators and pollen data clearly reflect the onset of agriculture (6000 b2k) and the middle Neolithic (5400 b2k) expansion of agricultural activity (Dreibrodt and Wiethold, 2015). Kalis et al. (2003) describe the onset of late Neolithic human impact as ‘amazingly’ contemporaneous in the Juessee, Steisslinger See, Schleinsee and Degersee in northern and southern Germany, dating to around 6300 b2k.
Several Finnish lakes exhibit first signs of early land-use influences associated with Neolithic (5000–3500 b2k) and clear signs of enrichment in the Iron Age (around 2500 b2k) (Räsänen et al., 2006; Tolonen et al., 1976). Likewise, Bronze Age (3500–2500 b2k) forest clearance and cultivation activities inferred from a study of Diss Mere, England, were linked closely in time with changes in the diatom record (Fritz, 1989). In Italy, a close link was observed between forest clearance and the cladocera assemblages of lakes Albano and Nemi, which shifted from Daphnia to Bosmina around 3500 b2k (Guilizzoni et al., 2002).
In southwestern Sweden, the lake Lilla Öresjön recorded the expansion of agriculture (as seen by the appearance of cereal pollen) as a period with higher pH (as indicated by the diatom assemblage) starting in 2350 b2k (Renberg, 1990). In Kassjön, a small northern Swedish lake, Anderson et al. (1995) argue that the diatom assemblage is more sensitive to the start of agriculture than the pollen record. The varved record allows the changes to be determined very precisely, with dramatic changes in the diatom assemblages observed in 1268
Further east, Heinsalu and Veski (2007) reported palaeolimnological evidence of permanent rural land-use around Rõuge Tõugjärv, Estonia, from the onset of the Bronze Age (c. 3800 b2k). Diatom species shifts indicate a switch from a mesotrophic to eutrophic state, with no lag between the change in land use and the lake response. In Latvia, the palaeovegetation records from Lake Trikāta show increasing signs of human impact from 2500 b2k, with the beginning of continuous cereal cultivation (Stivrins et al., 2016). The most significant changes are, however, recorded much later, in the early 13th century
Even in regions that have been assumed to be naturally eutrophic, palaeolimnological records have shown that landscape changes elicited responses in aquatic ecosystems. In particular, Boyle et al. (2015) coupled changes in sediment flux, titanium (Ti) concentrations (as an indicator for soil erosion) and the P yield to change in population density, which in turn provided a quantitative record of human impacts spanning the last 6000 years in northwest England. The concurrent decline in arboreal pollen and doubling of P yield at c. 5300 b2k revealed an early influence of Neolithic farming on the aquatic ecosystem in the Cheshire and Shropshire meres that were assumed for decades to be naturally eutrophic.
From the Iberian Peninsula, pollen records dating back to Neolithic times (c. 6000 b2k) reflected first human impacts (Carrión et al., 2010 and references therein), but the first clear changes in erosion rates were detected during the Iron and Bronze Ages (starting at c. 4400 b2k; Carrión et al., 2007, 2010; Martínez-Cortizas et al., 2005). However, it is during the Iberian Roman period (500
In the Northern French peri-alpine region (Lake La Thuile, above 800 m a.s.l.), anthropogenically driven erosive phases began approximately 550
Soil erosion induced by agricultural activities is thus clearly the first human impact recorded in palaeolimnological records throughout Europe, but additional activities were also detected early on, such as mining or the processing of fibres. Local pollution derived from copper (Cu) smelting was, for instance, detected around 4000 b2k in Wales (Mighall et al., 2002). Almost simultaneously, an ‘ancient period of atmospheric lead (Pb) pollution’ was detected in multiple Swedish lakes (Renberg et al., 2000). Early mining impacts associated with Roman activities (2100–1800 b2k) were found throughout Europe as both an increase in heavy metal atmospheric deposition and run-off (Guyard et al., 2007; Renberg et al., 2001), although the impact on the aquatic systems remains unclear (Camarero et al., 1998, 2017; Hillman et al., 2017). The processing of fibres, especially hemp retting, on the other hand, had clear impacts on some aquatic systems, even though only nutrients (i.e. non-toxic pollutants) were released. Grönlund et al. (1986) show that the beginning of intensive hemp retting in c. 1590
Asia, Southeast Asia and Oceania
Asia
In China, first signs of human impact are again linked to catchment processes. Pollution and agricultural activities are both recorded around 3000 b2k. For instance, in the Yunnan region (China), local pollution derived from copper (Cu) smelting started c. 3400 b2k (Dearing et al., 2008; Hillman et al., 2015). Increases in lead (Pb), silver (Ag), zinc (Zn) and cadmium (Cd) were observed only from 1100
Human-induced soil erosion of large river basins in China has been detected by the volume of deltaic-marine sediment deposition, starting 3000 years ago and accelerating 1000 years ago (e.g. the Yellow River; Syvitski and Kettner, 2011; Wang et al., 2007). In the Erhai lake-catchment system, southwest China, Dearing et al. (2008) identified a period of agricultural expansion around the 6th century
Lakes across the lower Yangtze River Basin in China have shown long-term eutrophication with substantial loss of biological diversity and deterioration of the lake ecosystems. Diatom proxies such as Cyclotella meneghiniana, C. atomus and Cyclostephanos dubius, that are responsive to total P enrichment, suggest that increased nutrient concentrations in these lakes began around the late-1700s to the early-1800s
Further to the west, the Aral Sea (Uzbekistan) is well known as an example of anthropogenic desiccation. Intensive irrigation in the 20th century
South and Southeast Asia
The Sundarban (Bangladesh and India) foothills of the Nepalese and Indian Himalayas, the Mekong River delta (Vietnam) and the lower Yangtze River basin are examples of major wetland systems where the effects of humans and climate change are now evolving simultaneously (Erwin 2009; Kattel et al., 2016; Khadka and Ramanathan, 2013; Mushtaq et al., 2015; Kattel et al., 2016). Stratigraphic deposits from lakes, ponds and palaeochannels across the central Gangetic plains in India indicate strong climate-induced fluvial activity in the region until 7000–5000 b2k, followed by aeolian aggradation. Humans substantially accentuated these conditions by mobilizing the water resources since they began practising agriculture in the region, at the beginning of the Common Era (Gupta, 2004).
In central India, the invasion of ruderal plants such as Artemisia or Cannabis sativa in the open mixed deciduous forest from 7500 to 6250 b2k reflects the first detectable human impacts on the wetlands (Chauhan et al., 2013). Further west, several palaeolimnological studies related waxing and waning of the Harappan Civilization (aka the Indus Valley Civilisation) with drier and wetter climatic periods, albeit with conflicting interpretations (Enzel et al., 1999; Menzel et al., 2014; Singh et al., 1990). Singh et al. (1990) provided evidence for forest clearance and commencement of agricultural practices during Harappan times (c. 7000 b2k) on the basis of the presence of Cerealia-type palynomorphs, along with fine fragments of charcoal in several lakes (Didwana, Lunkarsar, Sambhar and Pushkar) of Rajasthan (western India). Fragments of charred grass epidermis with clear evidence for agro-pastoralism appeared in the high-altitude lake Tso Moriri (NW India) c. 2700 b2k (Leipe et al., 2014). At the same time, the first significant human impact on the landscape was detected in Lake Rukche (3500 m a.s.l. in central Nepal) consisting of a charcoal layer and a marked emergence of fire-induced pollen communities (Schültz and Zech, 2004).
No research specifically into human-induced aquatic ecological transitions has yet been undertaken in the Indian sub-continent. Given the pressing need to manage water resources in this part of the world, it should be a priority region for palaeolimnological and palaeoecological investigations.
Integrated palaeolimnological records and archaeological sequences covering the mid- to late-Holocene were recently synthesized in Northeast Thailand (Chawchai et al., 2013, 2015, 2016; Wohlfarth et al., 2016; Yamoah et al., 2016). Archaeological information found around the sites provides a chronological framework, from the initial Neolithic settlements by rice farmers (c. 3700 b2k) to the end of the prehistoric Iron Age around 700
Evidence for historical water resource management practices in regions such as Cambodia and Vietnam was dated as far back as the 13th century
Oceania
The antiquity of humans in Australia extends over 60,000 years (Figure 1a; Clarkson et al., 2017). However, other than the still contentious nature of the use of fire or the role in the extinction of megafauna (Field et al., 2013), there is little evidence of human impact on lakes and wetlands at this timescale. It is recognized that native populations increased over recent millennia and that their technologies evolved. In the Lake Condah region of Victoria, they are known to have built eel traps and smokehouses from 6600 b2k (McNiven et al., 2012). Palynological evidence suggests the traps modified the local hydrology sufficiently to increase the abundance of aquatic plants, but there is little evidence for the impact of this on the lake ecosystems through increased fish predation. It is only following the arrival of European colonizers in the 18th century that significant changes in the aquatic ecosystems are apparent in the palaeolimnological record (Gell et al., 2005a). For instance, land clearance, drainage practices and river regulation post-European settlement led to a suite of widespread impacts evident through the Murray River system (Australia; Gell and Reid, 2016), and associated impacts such as the dramatic salinization of Lake Curlip (Gell et al., 2005b; MacGregor et al., 2005) following the transfer of Snowy River water into the Murray.
Papua New Guinea is known as one of the independent birthplaces of agriculture, specifically Pacific agriculture (Bellwood 1990; Kirch, 2000). This technological leap – the transformation of plant exploitation to agriculture – shaped the numbers of humans, their food supply, and their cultures throughout Oceania. Early forest clearance and agriculture induced a sharp increase in erosion rates between 9000 and 6000 b2k in a highland catchment, but with no obvious damage or land degradation in the catchment (Hughes and Sullivan, 1991).
Lakes are rare on small oceanic islands, where rivers and groundwater provide most freshwater resources. During colonization of the Pacific Islands, humans often settled on islands and atolls devoid of lakes. On some islands, however, a few lakes provide palaeolimnologists with the possibility of investigating the timing of human arrival and the extent to which they transformed their constrained environment. For example, three crater lakes on Rapa Nui (Easter Island; the furthest outpost of Polynesian settlement) show that humans arrived there around 1200
New Zealand is the last landmass of considerable size to have been settled by humans (Figure 1a). The Māori arrived there around 1300
Americas
North America
Across North America, many palaeoecological studies have shown the evolution of post-glacial vegetation and aquatic change, but many of these were executed at a fairly coarse temporal resolution and thus do not always allow for the detection of pre-European human impact by First Nations inhabitants to lake ecology. Nevertheless, early human activities (e.g. agriculture, deforestation) have been noted in several studies starting around 1100
An example of a First Nations’ occupation around a temperate lake was provided by Ekdahl et al. (2004, 2007) who demonstrated that a 200-year-long Iroquoian settlement next to Crawford Lake (Ontario, Canada) permanently altered the ecosystem. In particular, these studies showed increases in the accumulation rates of Al, Na, K, Fe, and Ti, which reflected an influx of allochthonous siliciclastic material in sediments and were associated with increased erosion rates related to farming and deforestation during the Iroquoian occupation of the watershed (1268–1486
Central America and Mexico
Several studies from southwest Mexico down to Panama report that lake edge settings were being exploited by 7000 b2k through burning and farming, as shown by the presence of phytoliths, chipped stone artefacts, charcoal, maize and squash pollen (Piperno, 2006; Piperno et al., 2007; Ranere et al., 2009). However, no palaeolimnological evidence for the impact on the lake ecosystem has been described for this interval. The first ecosystem impact was detected around 3500 b2k in lakes in the volcanic highlands of Central Mexico: multi-proxy investigations of their sediment records clearly associated human activity such as forest clearance and agricultural expansion with aquatic degradation, including eutrophication (Metcalfe et al., 1989).
In Central America, large-scale human impacts on lakes started with the Mayan culture (Beach et al., 2015; Curtis et al., 1998). Pollen data from Lake Cobá, on the Yucatán Peninsula, indicates land clearing at 3600 b2k, with first maize pollen detected at 2800 b2k (Leyden et al., 1998). Economic, political, and demographic changes were recorded in the archaeological record of the Mayan Lowlands region during the Terminal Classic (800–1000
South America
Micro-charcoal deposits in lakes, reservoirs and bog sediments have been used to infer early human presence in South America (Supplementary Material S1.4, available online). A common pattern in South American fire histories is that they start with a large charcoal signature indicating initial burning, followed by a stepwise decay in charcoal abundance (Bush et al., 2007). The pre-Hispanic period from 5000 b2k to 500
Several palaeolimnological studies from the northern region of South America revealed anthropogenic landscape disturbances. In Amazonia, human signatures of disturbance were shown by pollen, phytolith and charcoal data in continuous lake sediment records pointing to discrete periods of high and low levels of human activity from 7500 to 5500 b2k (Bush et al., 2016). In the Colombian savannas, Behling and Hooghiemstra (2000) attributed a marked increase of palm pollen at c. 3800 b2k (under the wettest Holocene climate regime) to human activities. In Bolivia, the first occurrence of maize pollen (a key crop for the region in pre-Columbian times) was taken as an indication of the start of agriculture: at c. 6500 b2k in the sediments of the Llanos de Moxos in the lowlands of northern Bolivia (Brugger et al., 2016), and at c. 2500 b2k in the Bolivian Amazon (Carson et al., 2015).
The only regional study that investigated aquatic ecosystem responses comes from Lake Sauce (Peru), where intense human activity between c. 4000 b2k and 1300
Peruvian lakes have also been investigated for early pollution. Hg pollution in Peruvian lakes was first detected starting around 3300 b2k (Cooke et al., 2009). It was attributed to cinnabar (HgS) extraction by the Chavín culture for the production of vermillion. Pb contamination, on the other hand, is evident from c. 400
Arctic
The small lakes and ponds that occur extensively in arctic and subarctic landscapes are highly sensitive ecosystems to environmental change (Korhola and Weckström, 2004). However, due to low population densities, traces of past human presence and impact in Arctic lake sediments are generally scarce. There are some indications that humans, either sedentary or semi-nomadic, were among the drivers of slight changes in lacustrine ecosystems in Finland around the beginning of the
At other sites in the High Canadian Arctic, the first detectable impacts of humans in the form of cultural eutrophication of lakes are much more recent (mid-20th century), such as those associated with military presence (e.g. Meretta Lake, Cornwallis Island, where hypolimnetic hypoxia developed rapidly in response to sewage enrichment; see Antoniades et al., 2011 and citations therein).
In Greenland, there is currently little evidence that the first inhabitants of various Palaeo-Eskimo cultures had any impact on aquatic ecosystems. Palaeolimnological investigations showed that the agro-pastoral lifestyle of the early Norse settlers increased erosion rates in a few lake basins (thereby increasing sediment accumulation rates in lakes) through the clearing of vegetation and grazing pressure between c. 985 and 1450
Recent climatic change and increased atmospheric deposition of pollutants such as metals and N have already significantly impacted physicochemical and biotic components of isolated high-altitude and high-latitude lakes (e.g. Catalan et al., 2013; Chen et al., 2013; Hobbs et al., 2016). Some even argue that no entirely pristine lakes remain, even in remote regions such as the Arctic (Wolfe et al., 2013). For the great majority of arctic lakes and ponds, these recent changes in their structure and functioning constitute the first detectable trace of human impact and reflect anthropogenic modification of global biogeochemical cycles.
Synthesis of lake responses to early human impacts
Sediment accumulation rates and turbidity
Sediment load entering a lake and lake-water turbidity are influenced by complex natural processes: the susceptibility of regional soils to erosion, hydrological regimes and residence time, exposure and sensitivity to wind-disturbance and re-suspension as well as external nutrient loading and internal productivity. Human pressures, including land-use change, have increased river-borne sediment and sediment accumulation rates at many sites over millennia (Dearing and Jones, 2003; Gell et al., 2009; Rose et al., 2011). Such high sediment loading is a pervasive type of pollution that can impede the ecological functioning and primary productivity of a lake (Donohue and Molinos, 2009; Wood and Armitage, 1997), which may trigger or be exacerbated by turbidity and large algal blooms associated with cultural eutrophication.
Many palaeolimnological studies from regions of the world that experienced a shift from hunter-gatherer to sedentary farming show early evidence of accelerated sediment accumulation rate (SAR) (c. 5200 b2k; Chiverrell, 2006; Dearing and Jones, 2003; Edwards and Whittington, 2001), although the biological effects on lake ecosystems at that time are rarely explicitly investigated. Mayan colonisation around Lake Salpetén during the Middle Preclassic (2500 b2k; Rosenmeier et al., 2002), the Norse arrival on Greenland (around 1000
Eutrophication and anoxia
While lakes naturally lie along a spectrum of nutrient loading (primarily N and P) from oligotrophic (clear water, low nutrient) to eutrophic (elevated nutrient supply, high primary productivity), cultural eutrophication of aquatic ecosystems is an acute anthropogenic impact (Smith, 2003). Excessive nutrient supply from domestic (e.g. sewage disposal), industrial (e.g. chemical discharge) and agricultural (e.g. fertiliser application) sources has triggered algal blooms, altered species composition, depleted deep water oxygen levels and reduced water quality and local aesthetic valuations in numerous lakes and wetlands worldwide (Pretty et al., 2003; Smith and Schindler, 2009). Unfortunately, the impacts of eutrophication on the ecology of lakes have many similarities with those of climate change, making it difficult to disentangle the impact of one from the other, in particular where the eutrophication impacts are greatest (Davidson and Jeppesen, 2013).
Cultural eutrophication of lakes and wetlands is strongly associated with industrialisation and urbanisation of catchments, with most European studies showing the first clear evidence of nutrient enrichment linked to human activity around the mid-19th century
The biological effects of eutrophication can be inferred from sedimentary organism assemblages; certain diatom taxa, in particular, are sensitive to specific nutrient concentrations (e.g. Hall and Smol, 2010; Tremblay et al., 2014). An abrupt appearance of abundant diatoms amenable to higher P and/or N loading is thus reliable evidence of lake eutrophication, especially after a period of landscape stability. For instance, the transition from a Cyclotella-dominated to a Stephanodiscus-dominated assemblage, or the appearance of species with high nutrient optima, such as Fragilaria crotonensis, is a widely employed indicator (Ekdahl et al., 2004; Fritz, 1989). Other biotic proxies can also be used to indicate eutrophication and associated aquatic transitions, such as chironomids (e.g. Brooks et al., 2001; Langdon et al., 2006; Zhang et al., 2012), and changes in plant macrofossil communities (e.g. Sayer et al., 2010). Sedimentary pigments are also a reliable proxy for pinpointing the onset of eutrophication through the detection of past changes in algal and bacterial community composition (e.g. Leavitt and Findlay, 1994). Anthropogenically driven changes in algal productivity are further indicated by increased CaCO3 accumulation, higher TOC and lower C/N values. A signal of historical nutrient status can be detected in δ15N (McLauchlan et al., 2013) and δ13C (Hollander and Smith, 2001) sedimentary ratios as well as other macrofossil records, such as the disappearance of plant-associated cladocerans from Australian wetlands (Kattel et al., 2014; Ogden, 2000).
A potentially severe consequence of intensifying eutrophic conditions is the reduction (hypoxia) or disappearance of dissolved oxygen (anoxia) in the hypolimnion (Diaz 2001; Friedrich et al., 2014). Whereas certain basin configurations are susceptible to aquatic anoxia through natural processes (e.g. where deep, confined zones preclude mixing; Davison, 1993; Zolitschka et al., 2015) excessive human-sourced nutrient loading and man-made alterations to hydrological flows have led to more frequent anoxic events of greater spatial extent (Jenny et al., 2016b ; Vitousek et al., 1997). For example, bottom-water anoxia has been ascribed to a human trigger where sediment records show a shift from naturally well-oxygenated conditions that coincide with proxy evidence of cultural eutrophication (Giguet-Covex et al., 2010).
Stratigraphic inspection has highlighted the presence of anoxia by an absence of bioturbation, owing to the disappearance of benthic fauna under deoxygenated conditions (Meyers and Ishiwatari, 1993; van Geen et al., 2003; Zolitschka and Enters, 2009). The appearance of seasonal laminations (i.e. varves) can pinpoint the onset of anthropogenically induced anoxic conditions to a particular year, as in Lake Bourget, France (Giguet-Covex et al., 2010; Jenny et al., 2013) or in Baldeggersee, Switzerland (Lotter and Birks, 1997). A recent synthesis of laminated records from Europe demonstrated that lake water hypoxia started spreading across sites affected by urbanization (Jenny et al., 2016b).
Anoxia also leads to the diagenetic loss of sediment P (Boyle, 2001; Rippey and Anderson, 1996), as well as the dissolution of several trace metals, an imprint detectable in core sequences (Belzile and Morris, 1995; Naeher et al., 2013; Tribovillard et al., 2006). Anoxic conditions have also been characterised by higher TOC content of sediment (Giguet-Covex et al., 2010), while certain biomarkers, including the 5β configuration of stenol or perylene, have been shown to form diagenetically under anoxic conditions (Meyers and Ishiwatari, 1993).
Multiple biological sediment proxies have been used to pinpoint the appearance of anoxic conditions. Overall, benthic invertebrate populations are likely to decline in deoxygenated water; Clerk et al. (2000) showed that during a phase of expanding human activity in Peninsula Lake (Ontario, Canada) beginning in c. 1870, there was an overall shift to chironomid taxa that characterize oxygen-depleted conditions (Chironomus, Procladius) and concurrent change from a benthic-dominated diatom assemblage to one dominated by mesotrophic planktonic species.
Acidification
Lake acidification is commonly associated with marked ecological impacts, such as changes in biological composition, water transparency and stratification. Timing for initial ecological change resulting from acidification follows historical industrial emissions, with earliest effects observed in the mid-19th century
Palaeoecological proxies have proven to be a key tool for reconstructing anthropogenically induced acidification, because of species-specific pH preferences and tolerances (Charles et al., 1990). Diatoms have been widely used, owing to the high sensitivity to pH of certain taxa (e.g. Battarbee and Charles 1986; Battarbee et al., 2010; Charles et al., 1990). The diversity of diatoms implies that some region- and site-specific indicators of acidification exist, though some, such as Tabellaria species, are widespread indicators (e.g. Flower and Battarbee, 1985). Diatom-pH transfer functions, which utilise the modern diatom subfossil assemblages to generate long-term quantitative reconstructions from core sequences of diatom assemblages, have been widely employed (e.g. Birks et al., 1990; Ginn et al., 2007; Simpson and Hall, 2012). The composition of cladoceran assemblages can similarly be ascribed to particular water pH (e.g. Paterson, 1994), while members of the genus Daphnia are known to be particularly sensitive to lake acidification (Keller et al., 1990). Similarly, certain species of chironomids are rare at low pH levels, offering potential as indicators of accelerated acidification, but uncertainties remain (Brodin and Gransberg, 1993). The multi-proxy approach of Arseneau et al. (2011) at Big Moose Lake (New York), which integrated diatoms, chrysophytes and cladoceran data, is an important means of verifying trends of lake acidification in light of varied response rates to pH fluctuations. In addition, a contemporaneous decline in dissolved organic carbon (DOC) content is often observed in acidifying lakes (Donahue et al., 1998) and vice versa during their recovery (Monteith et al., 2007).
Lakes may also acidify through the oxygenation of sulphides when coastal salt field sediments are exposed to air. The impact of acid sulphate soils in coastal zones have come to prominence particularly in regions where canals are constructed through coastal sediments for navigation, or for the development of residential resort complexes. In some circumstances, salinization and river regulation have combined to form sulphidic sediments that owe their origin to processes over decades rather than millennia. Exposure of these sediments through direct drawdown or drought similarly leads to wetland acidification (Gell et al., 2002; Hall et al., 2006).
The Anthropocene in palaeolimnological records
Lake sediment archives constitute an important element putting into perspective the magnitude of anthropogenic changes that occurred before/during the yet-to-be-defined start of the Anthropocene. Based on lake sediment archives, the most intense phase of human impact worldwide undoubtedly started in the mid-20th century (Waters et al., 2016; Wolfe et al., 2013), and many lakes located in remote regions have irrefutably recorded the signal of planet-wide human influence on the atmosphere, hydrosphere and biosphere since the beginning of the Great Acceleration (Catalan et al., 2013). However, the evidence presented in this review shows that the initial impact of humans on continental aquatic ecosystems (including their catchments) pre-dates the mid-20th century in most regions of the world. Although major restructuring of aquatic ecosystem function, or transitions away from their natural state, have been especially widespread since the mid-19th century, we have shown through this review that the scale of human impact on these systems has been spatially and temporally heterogeneous across the world, even going back several millennia in certain cases, depending on the timing of the arrival of humans and their degree of technical advancement. As these first human impacts were not globally synchronous, they cannot be considered to define a new geological Epoch. However, our review supports the concept of the ‘pre- (or proto-) Anthropocene’, as a period of flexible – region-dependent – duration, during which human impacts can already be detected but are not globally synchronous.
Conclusion
First human impacts to aquatic systems and their watershed have been highly variable in time and space. The first detectable impacts of human activity on lakes and wetlands often appear only after thousands of years of human presence in any given region (see Figures 1 and 3), but the time lapse between initial human presence and first impact shortens as the timing of initial settling approaches the present (e.g. New Zealand, see section ‘Asia, Southeast Asia and Oceania’). First anthropogenic landscape changes often pre-date by a few millennia (1000–4000 years, e.g. Morocco, Spain, Mexico; see Figure 3) the first detectable response in aquatic systems. However, exceptions exist, especially in Europe, where initial landscape modifications are synchronous with the first aquatic response (e.g. UK, Germany, Italy; Figure 3). Each region has its own environmental setting (e.g. geology, vegetation, climate) and history of human disturbances, which influences the susceptibility of its lakes to various threats (e.g. eutrophication in Europe, salinization and aridity in Asia). In addition, the source of impacts can be local, regional or global. Only in the industrial period have human impacts on lakes become widespread and severe through generalized air, water and soil pollution, and increasingly through species introductions and extirpations.
Identifying first impacts and associated aquatic responses are not without challenges. Detection of anthropogenic impacts leading to a change in ecosystem structure or function depends on lake characteristics, human activities conducted in the catchment and environmental proxies considered. Poor dating control and discrimination of appropriate sedimentary proxies remain major constraints for detecting the subtle effects of early human activities on lake ecosystems. Furthermore, many case studies reporting anthropogenic landscape disturbance did not look for corresponding ecological shifts. Until recently, lake sediment research focused strongly on generating climate records and on reconstructing regional changes in terrestrial vegetation rather than on identifying human-mediated changes caused by exploitation of land and water resources. However, while detecting human impacts at local scales can be challenging for the stated reasons, recent techniques are promising, including specific biomolecular markers (see Dubois and Jacob, 2016, for a review), ancient DNA (e.g. Anderson-Carpenter et al., 2011; Domaizon et al., 2017; Giguet-Covex et al., 2014), high-resolution stratigraphy and sediment scanning techniques (e.g. Arnaud et al., 2012; Flower et al., 2012; Marquer et al., 2008) as well as wider use of macrofossils combined with stable isotope studies (e.g. Hassan et al., 2012; Li et al., 2008).
Palaeolimnology has an important role to play in understanding ways in which people have brought about changes in freshwater ecosystems. Although landscape and environmental impacts of emerging civilizations may still be best realized by formulating and testing time-based hypotheses (cf. Deevey, 1969) predicated on the wealth of information that resides in cultural history and geoarchaeology, palaeolimnology provides a wealth of reliable data linked with environmental response to human activities. Given the paucity of published data in many regions (e.g. Asia, America, Africa, Australia), further studies are clearly necessary to improve our confidence in linkages between human settlements, their activities, and the downstream effects on lakes. It is further necessary to underscore that, while the effects of humans at the landscape level are relatively easily demonstrated, effects of human disturbance on structure and function of aquatic ecosystems need to be investigated further. To mitigate this deficiency, further palaeolimnological studies should be conducted with the specific aim of detecting the effects of human-induced changes on the structure and functioning of aquatic ecosystems.
Footnotes
Acknowledgements
Funding
The authors acknowledge the support of PAGES, and its umbrella IGBP and Future Earth, for funding workshops of the Aquatic Transitions Working group in Keyworth (UK), Lanzhou (China), Orono (ME, USA) and Kuala Lumpur (Malaysia).
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.