
Abstract
Introduction:
Breast cancer is the second most common cancer and a leading cause of cancer death in U.S. women. The tumor microenvironment, especially nearby adipocytes, plays a crucial role in its progression. Therefore, this study aimed to investigate the effects of human adipose mesenchymal stem cells-derived conditioned medium (SUP) and extract (CE) from on breast cancer cells.
Methods:
Human adipose-derived mesenchymal stem cells were isolated and characterized by flow cytometry using Cluster of Differentiation (CD) markers (CD34, CD45, CD90, and CD105). The differentiation potential was confirmed via adipogenic and osteogenic induction. MCF-7 and MDA-MB-231 cells were treated with SUP and CE, and cell viability was assessed using the 3-(4,5-Dimethylthiazol-2-Yl)-2,5-Diphenyltetrazolium Bromide (MTT) assay at 24, 48, and 72 h. Doubling time, colony formation, wound healing, and gene expression for key cancer-related genes (TIMP1, TIMP2, MMP2, PDL1, IDO, Bax, caspase 3, and caspase 9) were also evaluated.
Results:
Both SUP and CE significantly inhibited the viability of MCF-7 and MDA-MB-231 cells, reduced their doubling time, and suppressed colony formation. In wound healing assays, cell migration was notably impaired in MDA-MB-231 cells but less so in MCF-7 cells. Real-time polymerase chain reaction revealed downregulation of TIMP1, MMP2, PDL1, and IDO in MDA-MB-231 cells after treatment, while CE increased certain gene expressions in MCF-7 cells. Bax, caspase 3, and caspase 9 expressions were significantly upregulated in MDA-MB-231 cells but not in MCF-7 cells after treatment.
Conclusion:
Human adipose-derived mesenchymal stem cells-derived SUP and CE exhibit antitumor effects on breast cancer cells, suggesting a potential therapeutic strategy to suppress tumor progression. Mesenchymal stem cells-SUP and CE could be a safe and novel regenerative approach for breast reconstruction postmastectomy without tumor recurrence risk.
Graphical abstract

Introduction
Breast cancer is the most common malignancy and the second leading cause of death in women worldwide. 1 Unfortunately, the incidence and mortality rates are increasing in developing and developed countries. 2 The global burden of breast cancer in women is rising due to a lack of early detection, limited awareness of symptoms, and a lack of cost-effective treatments. 3 The current treatment modalities for breast cancer are surgery, chemotherapy, radiotherapy, immunotherapy, and endocrine therapy, which are usually not completely effective. 3 It seems that a novel efficient, anticancer therapeutic approach is needed.
Cancer cells can survive and progress through several mechanisms, including invasion and metastasis, evading immune detection, and creating an immunosuppressive environment. 4 Several genes are responsible for these processes, with tissue inhibitor of metalloproteinase 1 (TIMP1), matrix metalloproteinase 2 (MMP2), programmed death-ligand 1 (PDL1), and indoleamine 2,3-dioxygenase (IDO) being particularly critical.5,6 TIMP1 inhibits matrix MMPs, which are involved in tumor invasion and metastasis, whereas MMP2 facilitates extracellular matrix breakdown, which is crucial for cancer cell invasion. 5 By suppressing immune responses, PDL1 allows cancer cells to evade immune detection, and IDO creates an immunosuppressive environment by metabolizing tryptophan, aiding in tumor growth. 6 Additionally, B-cell lymphoma 2 (Bcl-2), Bcl-2 associated X protein (Bax) and caspase 3 and 9 are essential regulators of apoptosis. 7 Understanding these molecular mechanisms is crucial for developing effective treatments, and regenerative medicine offers promising strategies in this regard.
Regenerative medicine is an emerging field that uses three strategies—cell/stem cell therapy, tissue engineering, and gene therapy—to repair, regenerate, or replace cells, tissues, and organs, and is particularly relevant to cancer treatment due to its potential to both restore damaged tissues and target cancer cells. 8 Several promising regenerative approaches, such as cell-based therapy, are now under investigation to fight against cancers. 9
Mesenchymal stem cells (MSCs), which are multipotent cells with high self-renewal and differentiation potential, are found in different types of tissues, including bone marrow (BM), adipose tissue, dental pulp, placenta, and umbilical cord. 10 MSCs have been used for efficient therapeutic applications in various diseases due to their plasticity, intrinsic homing to injured or inflamed sites, and secretion of numerous soluble bioactive molecules such as growth factors and cytokines. 11 MSCs carry out their functional mechanism through direct cell-to-cell contact and secretion of paracrine factors (including extracellular vesicles and soluble factors). 12 The regenerative potential of MSCs has been primarily attributed to paracrine factors, 13 some recent studies reported that MSCs could be used for cancer therapy as both a therapeutic agent 14 and a drug delivery vehicle. 15
Adipose-derived stem cells (ASCs) are a group of stem cells that are easier to harvest and yield higher volumes compared to MSCs from BM and cord blood. ASCs also exhibit a longer lifespan, higher proliferative capacity, shorter DT, and later in vitro senescence than BMMSCs. 16 Alongside these benefits, ASCs secrete a potent array of growth factors, cytokines, and chemokines that have demonstrated therapeutic effects in various cancer types.16,17 Similarly, stromal vascular fraction, 18 a diverse group of cells including MSCs isolated from adipose tissue, also holds therapeutic potential and is being explored in breast cancer treatment.19,20 The isolation of ASCs and SVF from adipose tissue can be achieved through various methods, including enzymatic digestion, explant outgrowth (mechanical microfragmentation), and mixed enzymatic-explant digestion. 21 Reviewed studies have generally reported positive outcomes and a favorable safety profile for SVF therapy, with complications rarely mentioned, suggesting relatively low risk. 19 However, in postoncologic reconstruction, SVF therapy is approached with caution due to concerns about potential tumor progression.18,22
Additionally, the clinical application of MSCs in cancer therapy remains distant, primarily because of the risk of immune rejection of stem cell allografts. 23 Several potential adverse effects of MSCs administration, including tumor formation, growth, and metastasis, need to be carefully considered. Despite the concerns, clinical trials of allogeneic MSC-based therapy have not observed malignancy or tumor recurrence. 13 Interestingly, the secretome or CE has shown similar therapeutic effects with no MSCs potential adverse effects, and they have now emerged as potential candidates for cell-free anticancer treatment. A promising role of MSCs secretome or CE may be the potential ability to regenerate and reconstruct postsurgical tissues (e.g., breast tissue reconstruction after mastectomy) with no morbidity. Furthermore, breast cancer cells can be categorized based on estrogen receptor alpha (ERα) expression into ERα-positive or ERα-negative cells, a distinction that plays a critical role in determining treatment approaches. 24
Adipose tissue constitutes the primary microenvironment for breast cancer, and ASCs play a crucial role in this environment. 25 ASCs have shown a significant impact across various cancers. These cells can induce apoptosis and modulate the tumor microenvironment through immunosuppression, but their effects on tumor progression differ between cancer types. 26 For instance, research indicates that ASCs can inhibit prostate cancer cells while promoting the growth of colorectal cancer cells, highlighting their complex and varied roles in different malignancies. 27 Additionally, there are some controversies surrounding the use of ASCs in breast cancer patients despite their notable characteristics. 28 Thus, our study aimed to evaluate the effects of SUP and CE of hAMSCs on the proliferation, apoptosis, DT, colony formation ability, migration, and genes involved in tumor invasion, metastasis, and immune evasion (TIMP1, MMP2, PDL1, IDO), regulators of apoptosis (Bcl-2, Bax, caspase 3 and 9) in both ERα-positive (MCF-7) and ERα-negative (MDA-MB-231) breast cancer cells.
Materials and methods
Extraction of MSCs from human adipose tissue
The study protocol was approved by the Ethics Committee of Rafsanjan University of Medical Sciences, Rafsanjan, Iran (Code: IR.RUMS.REC.1399.098). For this study, hAMSCs were isolated and cultured according to the method described in a previous study. 29 Briefly, lipoaspirates of healthy adult donors were obtained after written consent and approval. Adipose tissue samples were washed twice with phosphate buffered saline (PBS; Gibco, UK) and treated with 0.1% type 1 collagenase (45 min). The inactivation of collagenase activity by adding an equal volume of Dulbecco’s modified Eagle’s medium (DMEM; Gibco) containing 10% fetal bovine serum (FBS; Gibco), the mixture was centrifuged at 1000g for 5 min. The supernatant was removed, and the cell pellet was resuspended in complete medium (DMEM/F12 (Gibco) supplemented with 100 U/mL penicillin–streptomycin (Gibco), 1% glutaMAX (Gibco), and 10% FBS and seeded into the culture flasks (SPL, Korea) at a density of 100,000 cells/cm2. After 1 day, the culture medium was replaced to eliminate nonadherent cells. The culture medium was changed every 3 days, and the cells in passages 3 to 6 were used for the following experiments.
MSCs characterization
Flow cytometry analysis was used to identify the MSC surface marker profile. 30 Approximately, 1 × 106 cells at passage three were treated with specific phycoerythrin (PE)- or fluorescein isothiocyanate (FITC)-conjugated mouse antihuman CD34, CD45, CD90, and CD105 antibodies (Sigma, USA). Cells were analyzed using a Beckman–Coulter flow cytometer and FACScan program (eBioScience, USA).
The differentiation potential of hAMSC was assessed in the same manner as previously reported. 29 For adipogenesis and osteogenesis, cells at passage three were cultured in specialized culture media to induce adipogenic and osteogenic differentiation as describe in manufacture protocol (Kia zist, Iran). After 3 weeks, adipogenic and osteogenic differentiation was confirmed using Oil red O (Merck, Germany) and Alizarin red S (Merck) staining, respectively. 29
Preparation of SUP and CE
In order to prepare SUP, the complete culture medium of hAMSC was harvested from the culture flask after 48 h of culture, when the cells reached a confluence rate of 70%–75% or higher, and then frozen under sterile conditions (−80°C). 31
In order to prepare the CE, 1 × 106 hAMSC at passages 3–6 were isolated after trypsinization, washed twice with PBS, resuspended five times the packed cell volume, and then three successive freeze/thaw cycles were done. After centrifugation (12,000 g, 10 min, 4°C), the supernatant was immediately stored at −80°C. 31
Cell viability analysis
MTT assay was performed to assess cell viability. The MCF-7 (ATCC HTB-22) and MDA-MB-231 (ATCC HTB-26) cells were seeded in 96-well plates (SPL) and cultured in DMEM medium supplemented with 10% FBS. After 24 h incubation, 100 µL of fresh medium (control), SUP, and CE were added into each well and incubated for 24, 48, and 72 h. At the end of incubation, MTT solution (20 µL, 5 mg/mL in PBS) was added to each well and incubated at 37°C for 2 h, after which time the supernatant was removed, and 100 μL DMSO (Merck) was added. The plates were incubated and shaken at 600 rpm for 15 min, and the absorbance was measured at 570 nm using a spectrophotometer ELISA plate reader (BioTek ELX800). All experiments were performed in triplicate and repeated four independent times.
Doubling time
After 72 h treated with SUP and CE, MCF-7 and MDA-MB-231 cells were obtained by trypsinization and counted using a hemocytometer (BLAUBRAND®), then DT or time of cell duplication was calculated according to the following formula 32 :

Investigation of colony formation potential
Five cells per cm2 were cultured into 25 cm2 cell culture flasks (SPL) and incubated for 14 days. At the end of incubation, colonies were washed with PBS and fixed with 4% formaldehyde (Merck). Cell culture flasks were then stained with 0.5% crystal violet (Sigma), and the numbers of colonies with more than 50 cells were counted. 32 Colony counting was performed manually using an inverted microscope.
Wound healing assay
The cancer cells were seeded in a 12-well plate (SPL) at a density to achieve a well-adhered and monolayer confluent (approximately 90%) after 24 h. The wounds were generated using a sterile 100 μL pipette tip, and detached cells were removed by washing with PBS. Fresh medium, SUP, and CE were added into each well, and cell migration was observed at 0, 24, and 48 h. 15
Study of gene expression by real-time PCR
In this study, gene-specific forward and reverse primers were designed using Primer-BLAST tool (https://www.ncbi.nlm.nih.gov/tools/primer-blast/). The oligoanalyzer tool (IDT) was used to validate the intrinsic properties of the primers, including melting temperature, GC content, hairpin formation, and dimerization. The designed primers are shown in Table 1. The expression levels of TIMP1, TIMP2, MMP2, PDL1, IDO, Bcl-2, Bax, and caspase 3 and 9 genes were analyzed by real-time PCR (Q2000C, LongGene). Total RNAs from the study groups were extracted using the RNA extraction kit (Pars Tous, Iran) according to the manufacturer’s protocol. RNA purity and concentration were assessed by spectrophotometry. Total RNA was reverse transcribed into cDNA using the cDNA Synthesis Kit (Pars Tous).
Primers list for real-time PCR.

Real-time PCR was performed to determine the level of mRNA expression using SYBR green Master Mix (Ampliqon). The sequences of the PCR primers are presented in Table 1. Relative gene expression was calculated using the 2−ΔΔCt method, with GAPDH (glyceraldehyde-3-phosphate dehydrogenase) as the reference gene. 33
Statistical analysis
All data are presented as mean ± standard deviation (SD). One-way ANOVA was performed using SPSS 19 software (IBM SPSS Inc., USA). Statistical significance was considered when p < 0.05. Experiments were performed in triplicate and repeated three times.
Results
MSCs characterization
Isolated hAMSC showed fibroblastic morphology (Figure 1(a)) with plastic adherent properties to the culture plate and were positive for the MSC-specific markers CD90, CD105 and negative for hematopoietic markers CD34 and CD45 (Figure 1(b)). Positive staining with Oil red O and Alizarin red confirmed adipogenic (Figure 1(c1)) and osteogenic (Figure 1(c2)) differentiation capacity of isolated hAMSC under in vitro condition.

MSCs characterization. (a) Fibroblast-like morphology of hAMSCs (×40, Scale bar = 100 µm). (b) Flow cytometry histogram of CD markers on hAMSCs at passage 3, positive for CD90, and CD105 markers and negative for CD34 and CD45. (c) Multilineage differentiation potential of hAMSCs (c1: Adipogenic; c2: Osteogenic) (×40, Scale bar = 100 µm).
The effect of SUP and CE cell viability and cell proliferation
An MTT assay was used to assess the impacts of SUP and CE on cell viability and cellular metabolic activity. The data showed that CE had no significant changes in MCF-7 viability after 24, 48, and 72 h treatment. In contrast, SUP treatment significantly reduced MCF-7 cell viability at 48 h but promoted cell proliferation at 72 h (Figure 2(a)).

Effect of SUP and CE on cell viability in breast cancer cells. (a) MCF-7 cells were treated with SUP and CE for 24, 48, and 72 h (***p < 0.001 vs MCF-7 group). (b) MDA-MB-231 cells were treated with SUP and CE for 24, 48, and 72 h (***p < 0.001 vs MDA group). The cell viability was quantitated by MTT assay. The data were presented as means ± SDs, representative of three independent experiments.
The findings of the study revealed a noteworthy observation that the treatment of MDA-MB-231 cells with CE resulted in cytotoxicity after 48 and 72 h of exposure. Interestingly, the results showed that CE was cytotoxic for MDA-MB-231 after 48 and 72 h of treatment. However, the cell proliferation rate of MDA-MB-231 was not significantly affected by SUP treatment (Figure 2(b)).
Doubling time
As presented in Table 2, SUP remarkably increased cell number and decreased DT in MCF-7 but no significant change in MDA-MB-231 cancer cells. In contrast, treatment with CE reduced cell number and increased DT in MDA-MB-231 cancer cells.
Doubling time (DT) and cell number of MCF-7 and MDA-MB-231 breast cancer cells in different groups after 24 h. The data represent the mean ± SD (***p < 0.001 vs MCF-7 group; ###p < 0.001 vs MDA-MB-231 group).
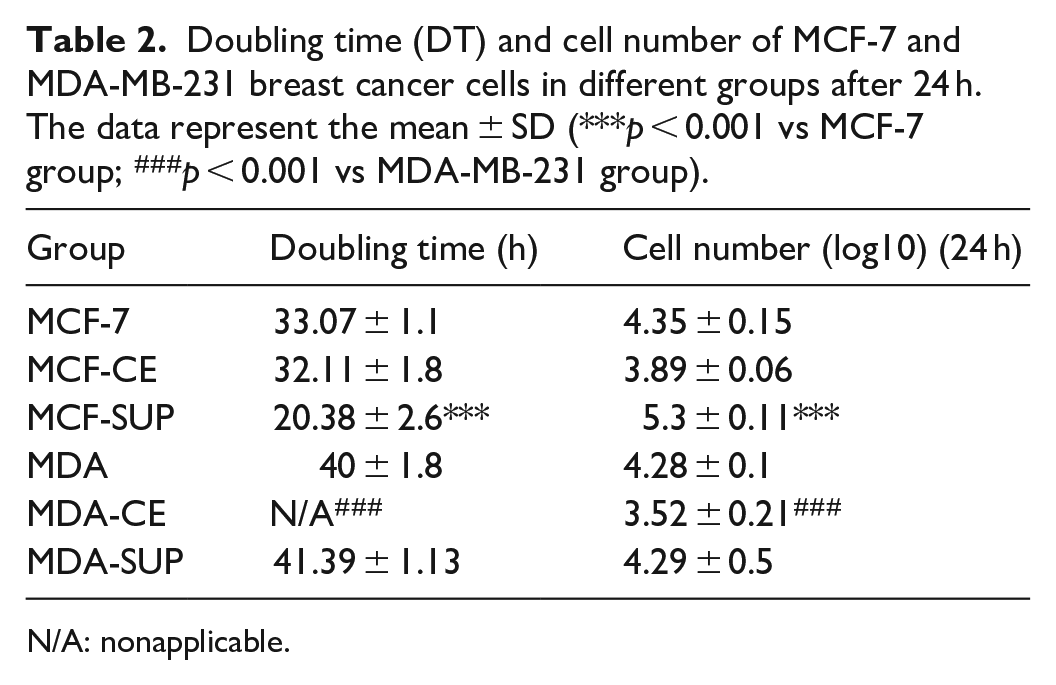
N/A: nonapplicable.
Colony formation potential
A colony formation assay was used to determine the effects of SUP and CE on the clonogenic potential of breast cancer cell lines (Figure 3). Based on this assay, both SUP and CE significantly decreased the capacity of MCF-7 and MDA-MB-231 cells to form colonies. Figure 3 shows that CE has more inhibitory effect on the colony formation than MCF-7 in comparison to SUP.
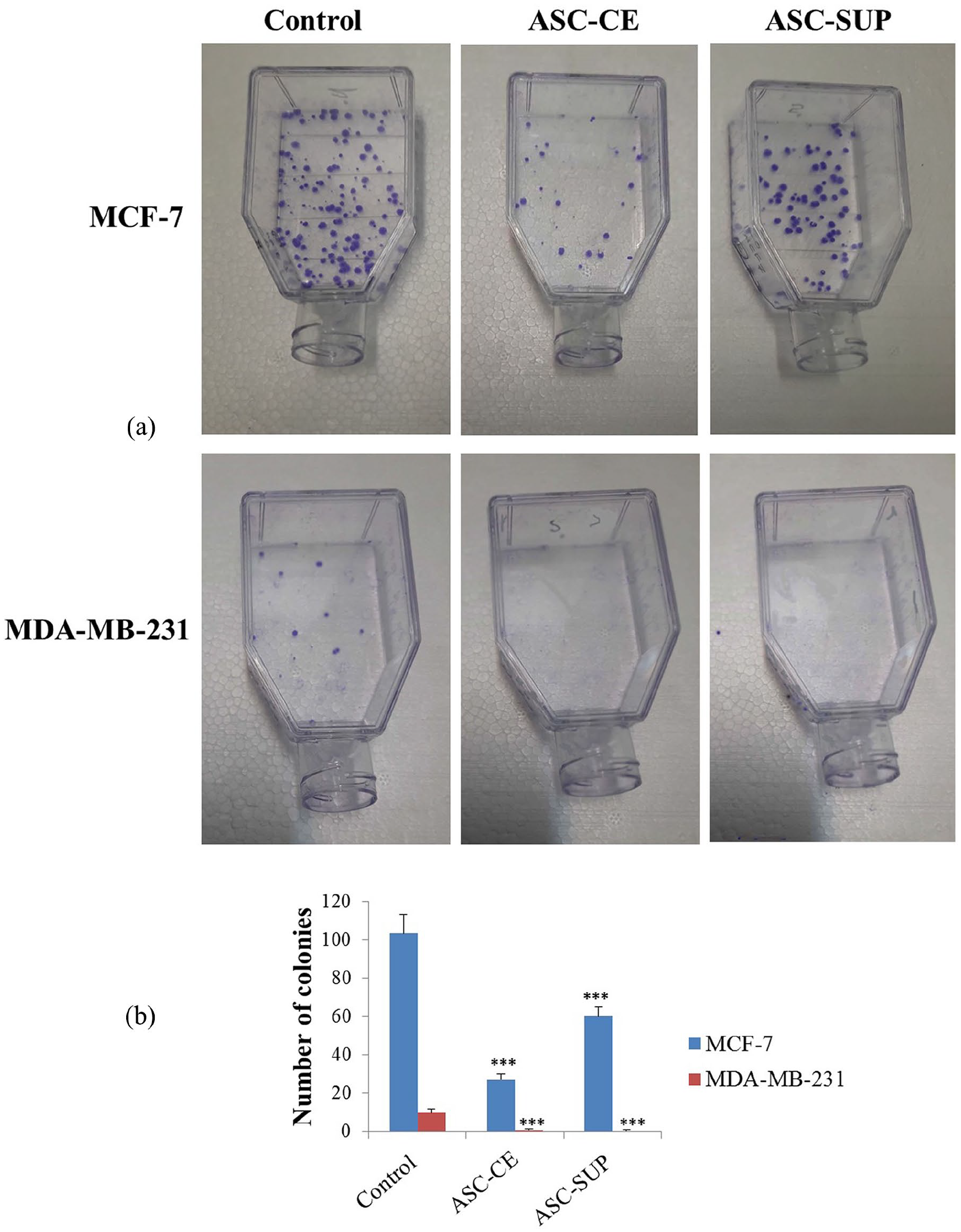
Effect of SUP and CE on colony formation capability of breast cancer cells. (a) Representative colony formation assay. (b) The colony formation histograms for MCF-7 and MDA-MB-231 cells after treatment with SUP and CE. The data were presented as means ± SDs, representative of three independent experiments. ***p < 0.001 versus untreated control groups.
Wound healing assay
A scratch test was performed to investigate the impact of SUP and CE on breast cancer cell migration. Changes in cell migration were confirmed by observing the extent of wound closure using an inverted microscope (Figure 4(a)). After 24 h of treatment, the speed of the scratch healing in MCF-7 cells was accelerated, indicating that both SUP and CE induced robust migrating capacity of MCF-7. Higher wound closure was found following treatment with SUP compared to CE.

Effect of SUP and CE on migration of breast cancer cells. (a) Representative wound healing assay of MCF-7 and MDA-MB-231 after 0, 24, and 48 h treatment. (b) Graph of the wound width percentage histograms for MCF-7 and MDA-MB-231 cells after treatment with SUP and CE. The data were presented as means ± SDs, representative of three independent experiments. ***p < 0.001 versus untreated control groups.
The results of 24 h treatment of MDA-MB-231 cells were the opposite, and scratch healing was significantly inhibited. As shown in Figure 4(b), the migration of SUP-treated MDA-MB-231 was enhanced compared with CE.
Similar results were observed after 48 h treatment with SUP and CE. The migration rate of MCF-7 and MDA-MB-231 markedly increased and decreased, respectively. There was no significant difference between the SUP and CE treatment groups in both MCF-7 and MDA-MB-231 cells (Figure 4(b)).
Gene expression
Real-time PCR was carried out to investigate some genes expression involved in cancer development: (1) invasion and metastasis (TIMP1, TIMP2, MMP2); (2) immune response (PDL1, IDO); and (3) apoptosis (Bcl-2, Bax, caspase 3, caspase 9).
The treatment of MDA-MB-231 cells with SUP and CE was found to induce a significant down-regulation of TIMP1, TIMP2, and MMP2, which are known as matrix effectors. Treatment with SUP and CE resulted in significant down-regulation of TIMP1, TIMP2, and MMP2 as matrix effectors in MDA-MB-231 cells. Interestingly, CE-treated MCF-7 cells showed a significant increase in TIMP1 and MMP2 gene expression, whereas TIMP2 was upregulated after treatment by SUP (Figure 5(a)–(c)). As shown in Figure 5(d) and (e), the expression levels of PDL1 and IDO, immune-related genes, were decreased in SUP- and CE-treated MDA-MB-231 cells. Although CE treatment led to an increase in the expression levels of both PDL1 and IDO genes in MCF-7 cells, indicating a potential pro-tumorigenic effect, SUP treatment resulted in a significant reduction in PDL1 expression. We found that after treatment with SUP and CE, the pro-apoptotic genes (Bax, caspase 3, and caspase 9) were remarkably upregulated in MDA-MB-231 cells, whereas no significant changes were observed in MCF-7 cells. Antiapoptotic gene Bcl-2 showed no significant difference in MDA-MB-231 and MCF-7 cells after SUP and CE treatment (Figure 5(f)–(i)).

Effect of SUP and CE on gene expression of breast cancer cells. Relative mRNA expression data of genes related to invasion and metastasis (TIMP1 (a), TIMP2 (b), MMP2 (c)); immune response (PDL1 (d), IDO (e)); and apoptosis (Bcl-2 (f), Bax (g), caspase 3 (h), caspase 9 (i)) after SUP and CE treatment in MCF-7 and MDA-MB-231 cells. The data were presented as means ± SDs, representative of three independent experiments. *p < 0.05, **p < 0.01 and ***p < 0.001 versus untreated control groups.
Discussion
Breast cancer, the most typical type of tumor in women, has the high potential to grow, invasion, and metastasis with very complex pathogenesis. Despite extensive research and development of novel systemic therapeutic approaches in recent decades, cancer remains one of the most challenging diseases. Many studies have investigated the effects of MSCs on tumors in recent decades. Controversial results have shown the dual impact of MSCs on tumor progression and suppression. 34 It has been found that MSCs are recruited to the tumor environment and can promote cancer progression through differentiation into cancer-associated myofibroblasts, cancer stem cell niche maintenance, and secretion of angiogenic factors. 35 Nevertheless, risk and safety should be essential considerations in the use of MSCs to treat cancer.
Compared with MSCs, extracellular secretions and CE of MSCs have several advantages, including higher safety and stability, lower tumorigenic risk, less immunogenicity, and lower lung entrapment. 36 Many studies have reported that MSCs secretions can promote tumor progression, but others have suggested their inhibitory effects on tumor progression. 37 To our knowledge, our study, for the first time, investigates the possible suppressive effects of hAMSC-derived SUP and CE on breast cancer cells.
The results of cell viability and DT assays indicated that hAMSC CE appears to have cytotoxic and inhibitory effects on TNBC cells, whereas SUP is ineffective on growth inhibition and instead increases the risk of proliferation of breast cancer cells. However, both SUP and CE significantly decreased the ability of a breast cancer cell to develop into a colony or to unlimited division.
Some investigations have reported that CM from UCMSC (UCMSC-CM) and BMMSC (BMMSC-CM) dramatically inhibit cell proliferation in MDA-MB-23138,39 and MCF-7, 40 but another study found that UCMSC-CM increases MDA-MB-231 and MCF-7 cell proliferation. 41 Mirabdollahi et al., 42 demonstrated that UCMSC secretome has a cytotoxic effect on MCF-7 through the induction of apoptosis. Similarly, other studies also reported that MSCs-CM from different sources (such as BM, adipose, Wharton’s Jelly/umbilical cord, and amniotic fluid) could decrease cell viability and proliferation and increase cell apoptosis in various cell lines, including glioma cells, 31 squamous cell carcinoma cell lines, 43 hepatic cancer cells, 44 prostate cancer cells, 45 and melanoma. 46 Cell lysates from BMMSCs and Wharton’s jelly MSCs (WJMSCs) can potentially inhibit the proliferation of U87MG glioma cells by G1 cell cycle arrest. 31
The MCF-7 proliferation was also inhibited after treatment with BMMSC-CM. 47 He et al. indicated that the clonogenic potential of MDA-MB-231 and MCF-7 was inhibited by UCMSC-CM. 47 However, Li et al. found a controversial result. They reported that UCMSC-CM augmented the colony-forming ability in MCF-7 and MDA-MB-231. 41
In this study, by scratch test, we confirmed that SUP and CE markedly inhibited migration of TNBC cells with no effects on ER+/PR+/HER2− breast cancer cells, consistent with the previous studies.38,48 In contrast to the behavior of MDA-MB-231 cells, SUP and CE promoted the MCF-7 migration after 24 h and no changes in the migration after 48 h treatment compared with the control group. The observed difference in the migration may be attributed to specific biological context and mechanisms involved. The secreted factors from hAMSCs may target specific signaling pathways or molecular mechanisms more relevant to MCF-7 cells, or influence microenvironmental conditions that affect MCF-7 cell migration.
Our findings on MDA-MB-231 cells are in concurrence with a recent study, which revealed the inhibition of growth, proliferation, migration, and colony formation capacity in ovarian cancer cell lines, specifically A2780 and SKOV-3, following treatment with CM derived from human amniotic MSCs. 49 High production and secretion of cytokines and factors, such as interferon α (IFN-α), interferon β (IFN-β), IL-8, IL-12, tumor necrosis factor-related apoptosis-inducing ligand (TRAIL), TIMP1 and TIMP2 by MSCs are known to be responsible for their antitumor effects. 42 Inhibition of Wnt and Akt signaling pathways is a possible mechanism for the growth suppressor effects of MSCs on cancer cell lines. 42
MMP plays regulatory roles in invasion, immune system avoidance, and metastasis, whereas TIMPs effectively inhibit and regulate MMP proteolytic activity. 50 MMP/TIMP physiological equilibrium changes are reported in invasive and metastatic cancers. 51 Our results show SUP and CE of hAMSC decrease the expression of TIMP-1, 2, and MMP-2 in MDA-MB-231 cells. Conversely, in MCF-7 cells, the TIMP-1 and MMP-2 expression levels were significantly elevated after hAMSC CE treatment, while TIMP-2 expression increased after hAMSC SUP treatment. Consistent with these results, Khalil et al. previously reported that MSC-CM significantly increases the mRNA expression levels of TIMP-1, -2, and -3, and also reduces the mRNA expression levels of MMP-2 and -9 in different ovarian cancer cell lines. 52
PDL1, as a pro-tumorigenic factor, is crucial and responsible for cancer immune escape and tolerance and is expressed various metastatic and high tumor-grade cancer cells. Strauch et al. also reported that inflammation sites, such as cancer, promote PDL1 expression in MSCs by inflammatory cytokines. 53 According to this, it is suggested that the use of MSCs secretion or CE may have lower immune tolerance potential than MSCs administration in cancer treatment. In this study, we showed the anticancer activity of hAMSC-SUP via PDL1 downregulation in both ER-positive (MCF-7) and triple-negative (MDA-MB-231) breast cancer cells. Whereas in contrast to MDA-MB-231, PDL1 expression was upregulated in MCF-7 after treatment with hAMSC CE. Inhibition of PI3K and STAT3 pathways is considered the cause of PDL1 expression reduction in cancer cells. 54 A recent study found that PDL1 suppression promotes apoptosis and inhibits proliferation, tube formation, and migration in tumor cells. 55
Heme-containing enzyme IDO is another immunosuppressive molecule contributing to eluding anticancer immunosurveillance. 56 IDO inhibition has been proposed as a promising approach to (re)activation of anticancer immune responses. 57 Targeting GCN2 and mTOR pathways leads to IDO downregulation in tumor cells following IDO inhibitor therapy. 58 In this study, after treatment of MCF-7 and MDA-MB-231 cells with SUP and CE, we found that IDO had an expression pattern similar to PDL1.
The recent review article indicates that the ASCs 39 secretome, containing a variety of bioactive factors, has the potential to treat breast cancer by inducing apoptosis in cancer cells, though clinical trial results evaluating the safety and effectiveness of ASC-derived secretome therapy are still pending. 59
Additionally, the suppressive and antitumor activities of MSCs-derived CM have been attributed to various types of molecules, such as Dickkopf-1 (DKK1), fms-like tyrosine kinase-3 (FRTK3), IFN-α, IFN-β, IFN‑γ, IL-1RA, IL-2, IL-5, IL-7, IL-12, IL-15, IFN‑γ inducible protein-10 (IP-10), latency associated protein (LAP), tumor necrosis factor superfamily member 14 (TNFSF14) and TRAIL which effect on cancer cell properties, such as proliferation, cell cycle, migration, invasion, colony forming ability, and apoptosis. 47 Future studies are needed to identify the involved molecules that exert the antitumor effects in hAMSC-derived SUP and CE.
Mastectomy is a therapeutic procedure to treat breast cancer. In this regard, regenerative medicine is a promising field that aims to improve the quality of life and provide better cosmetic outcomes for women after mastectomy by restoring breast physiology and morphology without the morbidity or capsule contraction associated with conventional flap surgical techniques. The two crucial elements in regenerative medicine are stem cell therapy and tissue engineering. 60 Fat grafting, a well-established technique, is a natural and autologous approach for breast reconstruction and soft tissue repair, restoring breast shape and volume. Unlike more invasive procedures, fat grafting employs the patient’s own adipose tissue, leading to more natural-appearing and durable outcomes.61 –63 Additionally, the procedure involves smaller incisions and a shorter recovery process compared to implants or injectable fillers. Despite their advantages, fat grafts can be prone to complications, including infection, high resorption rates, oil cyst formation, fat necrosis, and calcification. 64 Interestingly, combining autologous fat grafting with SVF, which includes MSCs and progenitor cells, can significantly improve graft survival and make it a more effective option for breast augmentation.64 –66
Researchers are seeking to find out new strategies to regenerate or construct tissue-engineered breasts. The findings of this study suggest that the use of ASC-derived CE appears to be a safe and novel regenerative approach for the breast reconstruction following mastectomy without any risk of tumor recurrence.
This study has several limitations that should be considered. The primary limitation is its reliance on in vitro experiments, which may not accurately reflect the complex interactions that occur in vivo. The study’s findings may not fully translate to physiological conditions within the human body, and it only examined the effects of SUP and hAMSCs CE on a limited set of genes involved in cancer development. A more comprehensive analysis of transcriptional and signaling changes is necessary to understand the mechanisms of action. The study also does not provide a clear explanation for the observed differences in effects on MCF-7 and MDA-MB-231 cell lines, which could be attributed to various cellular mechanisms. Furthermore, the study did not provide a comprehensive analysis of the potential mechanisms by which MSCs CM and CE affect breast cancer cells, which are crucial for elucidating therapeutic potential.
Conclusion
We conclude that hAMSC-derived SUP and CE have promising antitumor activity in breast cancer cells with different ERα statuses, specifically ER-positive (⩾10% ER expression) and ER-negative (<1% ER expression). The cell-free approach may be an exciting and promising therapeutic option instead of MSC’s direct application for treating pathological conditions such as malignancy. While this study has shed some light on the potential of MSCs-CM and CE to combat breast cancer, there is still much to be learned. To truly understand their regenerative effects, we need to study them in animal models, where we can assess their antitumor potential and safety. Additionally, we should use more breast cancer cell lines and evaluate the effects of the SUP and CE over a longer period of time, which are critical areas for future research.
Footnotes
Acknowledgements
The authors would like to acknowledge RUMS’s support and contributions for providing financial assistance for this research. Additionally, they express their gratitude to the Molecular Medicine Research Center for providing the necessary facilities and resources for conducting the experiments.
Authors’ contributions
Faezeh Kazemi designed the experiments, contributed to sample preparation, and wrote the manuscript; Fatemeh Sadeghian designed the experiments and contributed to sample preparation; Ali Pirsadeghi contributed to sample preparation, wrote and revised the manuscript; Fatemeh Asadi designed the experiments, performed the experiments, supervised the project; Hossein Javdani designed the experiments and data analysis; Aliakbar Yousefi-Ahmadipour designed and performed the experiments, data analysis, wrote the manuscript, supervised the project.
Data availability statement
The data supporting this study’s findings are available upon request from the corresponding author.
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This project was funded by Rafsanjan University of Medical Sciences (Funding No. 98160).
Ethics approval and consent to participate
This study was approved by the Rafsanjan University of Medical Sciences ethical review board (Ethics Code: IR.RUMS.REC.1399.098).
Consent for publication
All authors have given their consent for publication.
Informed consent
Written informed consent was obtained from all subjects before the study.
Trial registration
Not applicable.